
四川省湿地种子植物多样性及区系研究
2017-06-05 15:18刘贤安
浙江林业科技 2017年1期
王 恋,王 娟,刘贤安,赵 丹
(1. 成都理工大学 地球科学学院,四川 成都 610059;2. 成都理工大学 生态资源与景观研究所,四川 成都 610059)
四川省湿地种子植物多样性及区系研究
王 恋1,2,王 娟2,刘贤安1,2,赵 丹1,2
(1. 成都理工大学 地球科学学院,四川 成都 610059;2. 成都理工大学 生态资源与景观研究所,四川 成都 610059)
根据四川省2011-2012年第二次湿地资源调查结果,对四川省湿地种子植物区系进行分析。结果表明,四川省现有湿地种子植物948种,隶属80科334属(裸子植物1科2属2种,被子植物79科332属946种),其中国家重点保护野生植物6种,珍稀濒危植物5种。湿地种子植物中大科、大属的优势现象明显,区系较为古老,特有属较少。从分布区类型看,科级水平上有8个类型、6个亚型,其中世界分布种数最多,泛热带成分次之,表现出较强的热带性质;属级水平上有14个类型、8个亚型,以温带区系成分为主,北温带分布及其亚型地位突出,表现出典型的温带性质;温带成分略多于热带成分,中国-喜马拉雅成分占较大比重,说明四川省湿地种子植物的分布受青藏高原隆升所形成的亚热带山地气候影响较大,从热带向温带过渡的区系特点;同时中国-日本成分比重也较大,表明与日本植物区系关系密切。科的分化强度为4.2,属的分化强度为2.8,种的分化强度为0.07,在科、属水平上均表现出较强的区系分化特征。相似性分析表明该区湿地种子植物区系与云南、西藏、湖北具有共同的区系起源和物种分化形成背景。
种子植物;物种多样性;区系组成和分化;湿地;四川省
湿地植物是指生活在地表经常过湿、常年积水或浅水的环境中,植物的基部浸没在水中,茎叶大部分挺于水面之上,暴露在空气中的植物,包括沼生植物、湿生植物和水生植物[1]。湿地植物是湿地生态系统的重要组成部分,在维持湿地生态系统功能上有重要作用。20世纪 70年代以来,我国学者对湿地植物组成及其区系特征、群落类型与结构、湿地植被类型划分等进行了研究。李恒等根据优势种和植物的生活型将泸沽湖的植被划分为 10种群落类型,并研究了横断山区的湖泊湿地植被[2-3]。阳小成、李英南、谭志卫、李小艳等学者在泸沽湖水生植物的植物种类、植被组成和分布类型等方面进行了深入研究[4-7]。赵佐成阐明了川西高原四县的水生植物群落结构和区系特征[8-10]。何飞等研究了川西水生维管束植物区系地理和生态类型[11-12]。但至今未见省域尺度上的湿地种子植物多样性、区系性质等方面的研究报道。基于四川省第二次湿地资源调查资料,从植物区系学角度,分析了全省湿地种子植物区系组成,并与邻近的云南、西藏及同纬度的湖北进行对比,旨在阐明四川省湿地种子植物区系特征以及与云南、西藏、湖北三省(区)的植物区系的分布和演化关系,为四川省湿地资源的保护提供理论依据。
1 研究区概况
四川省地处青藏高原和长江中下游平原的过渡带,26°03′ ~ 34°19′ N,92°21′ ~ 108°12′ E,总面积为48.6× 104km2,居全国第五位。四川盆地区属亚热带湿润气候,气温较高,无霜期长,雨量多,日照少,年均温16℃以上,年降水量1 000~1 400 mm;川西南山地区,冬暖夏凉,干湿明显,气候垂直变化大,年均温12 ~ 20℃,年降水量900~1 200 mm;川西高山高原区有暖温带、温带、寒温带和高原气候,冬寒夏凉,水热不足,年均温4~12℃,年降水量500 ~ 900 mm。四川省复杂的地质和自然地理条件,孕育了丰富多样的湿地植物资源。
2 研究方法
以2011-2012年四川省第二次湿地资源调查资料为基础,参考《中国植物志》、《四川植物志》、《中国湿地植被》、《中国常见湿地植物》[13-16]及相关文献进行补充,建立四川省湿地种子植物名录数据库。
参照吴征镒的世界种子植物科的分布区类型、中国种子植物属的分布区类型、论中国植物区系的分区问题和种子植物分区的类型及其起源和分化中关于中国种子植物的分布区类型[17-20],以及郝日明、李仁伟、谢大军等学者对四川省植物区系的研究[21-27],对四川省湿地种子植物区系科、属的分布区类型进行统计和分析,同时与湖北、云南、西藏湿地种子植物区系进行比较研究。
区系分化强度分析:
科的区系分化强度=属数/科数,属的区系分化强度=种数/属数,种的区系分化强度=种下单位数/原种数。
区系过渡性分析:
区系过渡性=温带区系成分比重/热带区系成分比重的比值。
相似性系数分析:
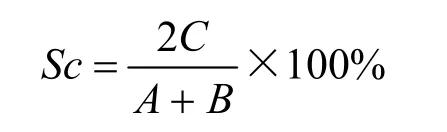
式中Sc代表两个区域的科(或属、种)相似性系数,C代表两地共有的非世界分布科(或属、种)数,A和B代表两地各自的非世界分布总科(或属、种)数。
3 结果与分析
3.1 湿地种子植物区系组成
3.1.1 科、属、种的数量组成 统计表明,四川省现有湿地种子植物948种,隶属80科334属。其中,裸子植物2种,隶属1科2属;被子植物946种,隶属79科332属(表1)。四川省湿地种子植物的科、属、种占四川省种子植物科、属、种的百分比分别为。42.33%,21.65%,9.37%,占全国湿地种子植物科、属、种的百分比分别为59.70%,52.68%,48.89%(表2)。

表1 四川省湿地种子植物Table 1 Spermatophyte in wetlands in Sichuan province

表2 四川省湿地种子植物与全国湿地和四川省种子植物科、属、种的比较Table 2 Comparison on families, genera and species of spermatophyte in wetlands in Sichuan province with that in China and total spermatophyte in Sichuan
3.1.2 重点保护及珍稀濒危物种 根据《国家重点保护野生植物名录(第一批)》和《中国珍稀濒危保护植物名录》统计,四川省湿地种子植物中有国家重点保护野生植物6种,其中国家Ⅰ级重点保护植物有2种,分别为水杉 Metasequoia glypt ostroboides和莼菜 Brasenia sc hreberi,国家Ⅱ级重点保护植物有 4种,包括金荞麦Fagopyrum dibotrys,莲Nelumbo nucifera,野菱Trapa incisa var. quadricaudata,芒苞草Acanthochlamys bracteata;中国珍稀濒危植物有5种,其中稀有植物有星叶草Circaeaster agrestis,桃儿七Sinopodophyllum hexandrum,渐危植物有八角莲 Dysosma versip ellis,海菜花 Ottelia a cuminate,极度濒危灭绝植物有疏花水柏枝 Myricaria laxiflora。
3.2 湿地种子植物区系组成分析
参照植物区系的科、属组成常用划分方法[27-28],根据各科或属在区系中所含种的多少,将四川湿地种子植物科划分为四个等级:①单种科,1种的科;②寡种科,2 ~ 9种的科;③中等科,10 ~ 19种的科;④大科,20种以上的科。四川湿地种子植物属也可划分为四个等级:①单种属,仅1种的属;②寡种属,2 ~ 5种的属;③中等属,6 ~ 9种的属;④大属,10种以上的属。
3.2.1 科的区系组成分析 统计结果表明(表3),寡种科所占比例最大,共39科,占总科数的48.75%,有车前科 Plantaginaceae、千屈菜科 Lythraceae等。单种科 18科,包含芒苞草科 Acanthochlamydaceae、金鱼藻科Ceratophyllaceae等。寡种科和单种科共占总科数的71.25%,但所含种数占总种数的18.46%,说明这些科在四川省湿地种子植物中占从属地位。中等科10科,主要包括伞形科Umbelliferae,天南星科Araceae等。大科包括禾本科Gramineae,莎草科Cyperaceae等13科637种,其科数仅占总科数的16.25%,但种数占总种数的67.19%,说明大科优势明显,在四川省湿地种子植物中占主导地位。

表3 四川省湿地种子植物科的组成Table 3 Family components of seed plants in wetlands in Sichuan province
3.2.2 属的区系组成分析 从植物属的组成来看(表4),单种属所占比例最大,共179属,占总属数的53.59%,包含白酒草属Conyza,芒苞草属Acanthochlamys等。其次是寡种属,共117属,占总属数的35.03%。中等属有23属,主要包括杜鹃属Rhododendron,酸模属Rumex等,占总属数的6.89%。大属15属,占总属数的4.49%,主要包括薹草属Carex,蓼属Polygonum等,共含有265种,占该区系总种数的27.95%,可见该区大属优势较为明显。

表4 四川省湿地种子植物属的组成Table 4 Genera components of seed plants in wetlands in Sichuan province
3.2.3 国家重点保护植物及珍稀濒危物种组成分析 四川省湿地种子植物中,国家重点保护和珍稀濒危共 11种,隶属于8科11属,分别占四川省湿地种子植物科、属、种总数的10%,3.3%和1.1%。其中单种属5属,单种科2种,单种科、属占较大比重。
3.3 湿地种子植物区系地理成分
3.3.1 科的分布类型 四川省湿地种子植物区系科的分布型共有9种(表5)。其中,世界分布42科,占总科数的52.50%,包含禾本科、莎草科、睡莲科Nymphaeaceae等,广泛分布于本区湿地植被中。热带分布21科,占非世界分布科的 55.26%,其中以泛热带及其变型为主,占热带分布的80.93%,如凤仙花科Balsaminaceae,鸢尾科 Iridaceae等,是四川湿地重要的植物资源,常构成湿地植被中的优势种或常见种。温带分布16科,占非世界分布科的43.24%,以北温带及其变型最多,占温带分布的 81.24%,含忍冬科Caprifoliaceae,黑三棱科Sparganiaceae等。中国特有分布1科——芒苞草科,分布于四川西部和西藏东南部。
3.3.2 属的分布类型 属作为分类学上最自然的类群,相互间能更好地划清界限,因此植物属的分布区类型比科更能具体反映植物的演化扩展过程、区域分异及地理特征。四川省湿地种子植物334属共有14个分布型8个亚型(表6),体现了该区系地理成分的复杂性。

表5 四川省湿地种子植物科的分布型Table 5 The areal-types of families of seed plants in wetlands in Sichuan province
3.3.2.1 世界分布 四川省湿地种子植物中世界分布59属,占总属数的17.66%,体现了该区系与世界其他地区区系的广泛联系。多为草本植物类群,其中龙胆属Gentiana和珍珠菜属Lysimachia为典型的世界分布属,薹草属、蓼属、蔊菜属Rorippa等为该区湿地植被的主要组成类型。
3.3.2.1 热带分布 共计104属,占非世界分布属的37.81%。其中泛热带及其亚型最多,共67属,其中冷水花属Pilea,凤仙花属Impatiens,飘拂草属Fimbristylis等的多种常见草本植物构成了四川湿地植被的优势种或建群种。热带亚洲和热带美洲间断分布包括凤眼蓝属Eichhornia,过江藤属Phyla等3属。旧世界热带分布及其亚型包括楼梯草属Elatostema,小丽草属Coelachne等10属。热带亚洲至热带大洋洲分布包括黑藻属Hydrilla等5属,多分布于河流、沟边或林缘。除泛热带分布外,热带亚洲至热带非洲在四川湿地种子植物的热带分布
类型中优势相对突出,共10属,其中水麻属Debregeasia,野茼蒿属Crassocephalum在四川湿地中较常见,是湿地植被中主要的伴生种。热带亚洲分布共9属,常见属有蛇莓属Duchesnea和薏苡属Coix。
3.3.2.3 温带分布 共计165属,占非世界分布属的60.00%。显然,温带分布型在四川省湿地种子植物区系中占有绝对优势,是最主要的地理成分。其中北温带分布及其亚型地位突出,共95属,占温带分布型的57.58%,报春花属Primula,马先蒿属Pedicularis,嵩草属Kobresia等构成了川西高寒湿地植被的主体成分。东亚和北美洲间断分布包含菖蒲属Acorus等8个草本属和落羽杉属Taxodium 1个木本属。旧世界温带分布及其亚型共22属,如橐吾属Ligularia等。温带亚洲分布共8属,如大黄属Rheum,虎杖属Reynoutria等,该分布属大多在喜马拉雅地区得到进一步的发展,向西南各省四周发散。中亚分布仅中亚至喜马拉雅和我国西南部分布亚型的高河菜属Megacarpaea和三角草属Trikeraia。东亚及其亚型共29属,占温带分布的17.57%,其中中国-喜马拉雅分布亚型有垂头菊属Cremanthodium,鬼臼属Dysosma等9属,中国-日本分布亚型有半夏属Pinellia等5属。其中垂头菊属是喜马拉雅山及毗邻地区的特有属,在高山草甸常成片分布。
3.3.2.4 中国特有分布 四川湿地种子植物属区系中共有6属,占区系非世界分布属的2.19%,包括裸蒴属 Gymnotheca,马蹄黄属 Spenceria,水杉属Metasequoia,虾子草属 Mimulicalyx,血水草属Eomecon和芒苞草属,其中血水草属是第四纪冰川后的孑遗成员,为第三纪古热带起源。芒苞草属是一个残遗单种属,分布于四川西部和西南部以及西藏东南部的横断山区腹地,以中国西南部的中国-喜马拉雅森林植物亚区为分布中心,是古地中海植物区系残遗的体现。
3.4 与邻近地区湿地种子植物区系组成比较
3.4.1 相似性分析 地区植物区系之间的亲缘关系通常由它们共有的科、属、种相似性系数来反映[29]。从表 7可以看出, 四川省湿地种子植物区系的科、属、种的组成与邻近的湖北、云南和西藏的区系联系都十分密切,科的相似性系数分别为63.01%,58.33%和55.74%,属的相似性系数分别为51.64%,42.30%和35.18%,种的相似性系数分别为39.20%,27.32%和23.09%。由此可见,四川省湿地种子植物区系与湖北、云南和西藏都属于近缘区系,可能具有共同的区系起源和物种分化形成背景;与湖北的相似性在科、属、种上均高于与云南和西藏相似性,说明与湖北亲缘关系更近。这是因为四川湿地小部分属于第一级青藏高原区域,而大部分处于第二级长江中下游平原,与处于同纬度的湖北省的地理格局更为接近,气候条件更为相似,所以相似性更高。
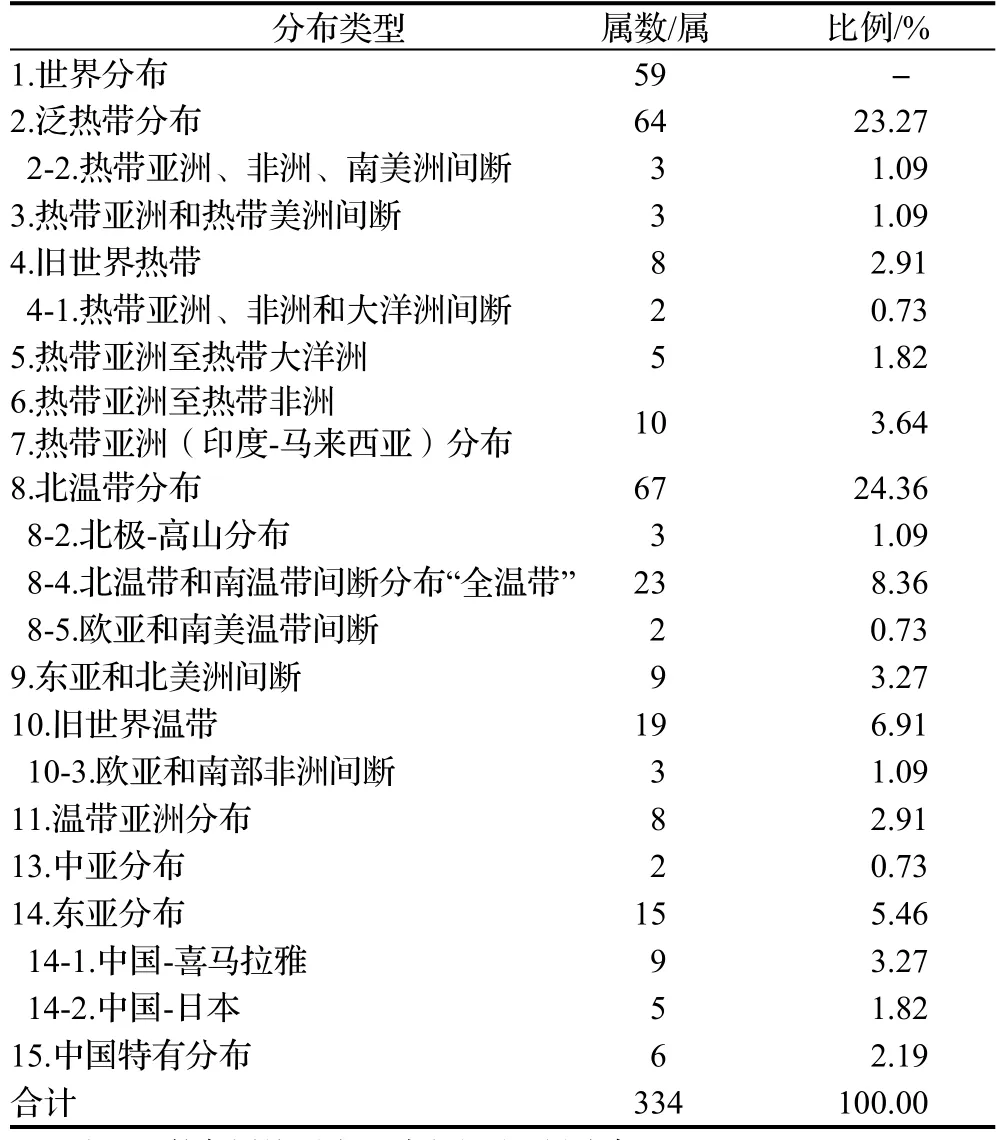
表6 四川省湿地种子植物区系属的分布区类型Table 6 The areal-types of genera of seed plants in wetlands in Sichuan province

表7 四川省与邻近地区湿地种子植物区系的共有科属种数及相似性系数Table 7 Similarity coefficients and number of same families, genera and species among wetland seed plants of Sichuan and neighboring provinces
3.4.2 组成比较 从表8可以看出,四川省湿地种子植物区系组成中温带和热带属性成分介于云南和西藏之间,与湖北相差不大,突出了地带性特点。4省区中国-喜马拉雅成分都较多,说明其植物区系的演化受到第三纪末第四纪初的喜马拉雅抬升和青藏高原隆起的影响较大,其中云南受到的影响最大,其次是四川。中国-日本成分,四川最多,云南、湖北次之,西藏最少,这是因为四川位于第一级青藏高原和第二级长江中下游平原的过渡带,受低海拔的丘陵和盆地气候影响较大,与日本区系联系较为紧密。东亚成分较高也说明了四川湿地种子植物区系在东亚地区具有代表性。

表8 四川省与邻近省区湿地种子植物区系组成成分比较Table 8 Floristic elements among wetland seed plants of Sichuan and neighboring provinces
3.5 区系过渡性和分化分析
3.5.1 区系的过渡性分析 从属的分布区类型分析可得,属的温带成分与热带成分所占比例分别为60.00%和37.81% ,说明四川省湿地种子植物区系存在强烈的温带性质,同时存在一定的区系过渡性,即四川湿地种子植物的温带区系与热带区系之间存在相互交汇关系。从环境角度看来,四川省湿地种子植物的区系过渡性与该地区的海拔梯度一致。四川地势地貌复杂多样,山川河流纵横交错,拥有丰富的高山高原、山地丘陵以及平原气候,海拔梯度差异较大,能量尤其是热量的梯度差异造就了丰富的生境,形成了多种多样的植物群落类型。从区系地理的角度来看,四川的过渡性与青藏高原的隆升及造山运动有关。四川曾在地质时期里长期处于热带——亚热带气候,随着青藏高原运动地层的抬升,高山高原和山地气候显著,整体气候趋于温凉,温带性质的植物迁入,使得该地区湿地种子植物区系的热带性质减弱,温带区系成分增多。由此可见,四川湿地种子植物表现出强烈的温带性质,同时向热带过渡。
3.5.2 区系分化分析 在科的水平上看,区系分化强度为 4.2;属的区系分化强度为 2.8;种的区系分化强度为0.07。其中区系分化能力最强的科主要为禾本科(63属)、菊科Compositae(35属)、莎草科(14属);分化能力最强的属主要为薹草属(43种)、蓼属(39种)、灯心草属(21种)、柳叶菜属(17种)。因此,四川湿地种子植物在科级及属级水平上表现出较强的区系分化特征,种级水平分化较弱,总体来说分化较为强烈。强烈的区系分化可能与四川的地质历史有关。随着造山运动的发生气候趋于温凉,地形趋于复杂,曾经适应亚热带气候的植物为了适应这种温凉的气候而出现显著的区系分化,同时,复杂的地形又对植物个体之间遗传信息的交流产生阻碍,经过长期的演化和进化,地理隔离和生殖隔离的发生形成了新的物种,加强了区系分化。
4 结论
四川省湿地种子植物种类丰富,共有80科334属948种,是中国湿地种子植物区系的重要组成部分。其中占总科数16.45%的大科包含总种数的67.05%;占总属数4.5%的大属包含总种数的27.98%,可见四川省湿地种子植物中大科及大属优势明显。
分布类型多样,地理成分复杂,具有明显的过渡性质。四川湿地种子植物区系类型十分复杂,成分丰富多样。科级水平上有8个类型和6个亚型,属级水平上有14个类型和8个亚型。区系过渡性分析表明四川省湿地种子植物区系具有较强的温带性质,同时表现出从热带向温带过渡的性质。不论科或属的分布型中世界分布都占有较大比例,表现出了四川省湿地种子植物区系同全国乃至世界其它植物区系有着广泛的联系。
植物区系较为古老,特有成分较少。自白垩纪以来,四川省气候相对稳定,没有受到第四纪大陆冰川毁灭性侵袭,保留了水杉属、血水草属等许多古老残遗植物,同时为植物区系演化创造了条件。四川湿地种子植物虽然种类较多,但中国特有属仅有6属,表明四川省湿地种子植物区系的个性特征不明显和湿生植被的隐域性。
受喜马拉雅抬升影响较大,与日本植物区系联系密切。四川省湿地种子植物区系中东亚成分的两个亚型——中国-喜马拉雅和中国-日本较多,说明湿地种子植物的分布受喜马拉雅和青藏高原隆升所形成的亚热带山地气候影响较大,同时与日本植物区系联系密切。
区系分化特征明显。四川省湿地种子植物在科级及属级水平上表现出较强的区系分化特征,强烈的区系分化可能与四川地区的地质历史有关。
与湖北、云南、西藏可能具有共同的物种起源和物种分化形成背景。从植物区系上看,四川湿地种子植物区系与湖北、云南、西藏的相似性都较高,可能具有共同的物种起源和物种分化形成背景。且区系组成及相似性分析都表明四川湿地种子植物区系与湖北亲缘关系更近。
[1] 严承高,张明祥. 中国湿地植被及保护策略[J]. 湿地科学,2005,3(3):210-215.
[2] 李恒,徐廷志. 泸沽湖植被考察[J]. 云南植物研究,1979,1(1):125-137.
[3] 李恒. 横断山区的湖泊植被[J]. 云南植物研究,1987,9(3):257-270.
[4] 阳小成. 沪沽湖的水生植被[J]. 重庆师范学院学报(自然科学版) ,1993,10(2):84-88.
[5] 李英南,赵晟,王忠泽. 泸沽湖特有水生生物的保护初探[J]. 云南环境科学,2000,19(2):93-95.
[6] 谭志卫,董云仙. 泸沽湖水生植被现状[J]. 环境科学导刊,2011,30(6):27-32.
[7] 李小艳. 高原湿地泸沽湖草海水生维管植物多样性研究[J]. 湖北农业科学,2015,54(15):3646-3655.
[8] 赵佐成. 四川省红原县水生植物群落调查[J]. 武汉植物学研究,1996,14(3):213-222.
[9] 赵佐成. 四川省泸定、康定县水生植物群落调查[J]. 武汉植物学研究,1996,14(2):147-152.
[10] 赵佐成. 青藏高原甘孜县水生植物群落调查[J]. 武汉植物学研究,1996,14(1):33-40.
[11] 何飞. 川西植物区系地理研究与优先保护区域分析[D]. 北京:北京林业大学,2009.
[12] 何飞,刘世荣,刘兴良,等. 川西水生维管植物区系地理及生态类型[J]. 湿地科学,2011,9(1): 33-41.
[13] 中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,2004.
[14] 四川植物志编辑委员会. 四川植物志[M]. 成都:四川人民出版社,1981.
[15] 郎惠卿,赵魁义,陈克林. 中国湿地植被[M]. 北京:科学出版社,1999.
[16] 张树仁. 中国常见湿地植物[M]. 北京:科学出版社,2009.
[17] 吴征镒,周浙昆,李德铢. 世界种子植物科的分布区类型系统[J]. 云南植物研究,2003,25(3):245-257.
[18] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究(增刊),1991:1-139.
[19] 吴征镒. 论中国植物区系的分区问题[J]. 云南植物研究,1979,1(1):1-20.
[20] 吴征镒,周浙昆,孙航,等. 种子植物分布区类型及其起源和分化[M]. 昆明:云南科技出版社,2006.
[21] 郝日明. 试论中国种子植物特有属的分布区类型[J]. 植物分类学报,1997,35(6):500-510.
[22] 李仁伟. 四川种子植物区系研究[D]. 广州:中山大学,2011.
[23] 王强,何兴金,周颂东,等. 马边大风顶国家级自然保护区种子植物区系研究[J]. 西北植物学报,2006,26(11):2345-2350.
[24] 李仁伟,张宏达,杨清培. 四川被子植物区系特征的初步研究[J]. 武汉植物学研究,2001,23(4):403-414.
[25] 李仁伟,张宏达. 四川裸子植物区系研究[J]. 广西植物,2001,21(3):215-222
[26] 李仁伟,张宏达. 四川种子植物区系组成的初步分析[J]. 武汉植物学研究,2002,20(5):381-386.
[27] 谢大军,向成华,张海军,等. 若尔盖高寒湿地自然保护区种子植物组成及区系分析[J]. 西南大学学报(自然科学版),2007,29(12):89-94.
[28] 王荷生. 植物区系地理[M]. 北京:科学出版社,1992.
[29] 朱华. 中国植物区系研究文献中存在的几个问题[J]. 云南植物研究,2007,29(5):489-491.
[30] 蔡述明. 湖北湿地[M]. 武汉:湖北科学技术出版社,2006,156-174.
[31] 杨岚,李恒,等. 云南湿地[M]. 北京:中国林业出版社,2010,370-433.
[32] 刘务林,朱雪林,等. 中国西藏高原湿地[M]. 北京:中国林业出版社,2013,306-326.
Diversity and Flora of Seed Plants of Wetland in Sichuan Province
WANG Lian1,2,WANG Juan2, LIU Xian-an1,2,ZHAO Dan1,2
(1. College of Earth Sciences, Chengdu University of Technology, Chengdu 610059, China; 2. Ecological Resources and Landscape Institute, Chengdu University of Technology, Chengdu 610059, China)
Based on the second wetland resources investigation in Sichuan from 2011 to 2012, analysis was made on flora of seed plants in wetlands of Sichuan province. The results showed that there were 948 species, belonging to 334 genera and 80 families, including gymnosperms (2 species of 2 genera and 1 family) and angiosperms (946 species, 332 genera and 79 families). Among them, 6 species are listed in national key protection and 5 ones are rare and endangered. The dominance of plurimotypic families and genera of wetland seed plants was obvious, and the flora was ancient with rare endemic genus. At family level, there were 8 types and 6 sub-types, dominated by Cosmopolitan and Pantropic, indicating strong tropical property, and at genus level, there were 14 types and 8 sub-types dominated by Temperate, especially North temperate elements, showing tropical temperate characteristics. It had slightly more Temperate elements than tropic ones, and Sino-Himalaya components were dominant, illustrating that distribution of wetland seed plants in Sichuan province had been strongly influenced by subtropical mountain climate led by Tibet plateau uplift, and reflecting a transition from tropical to temperate. Meanwhile, Sino-Japan components occupied large ratio, which indicated that it had chose relation with flora of Japan. Differentiation intensity of family was 4.2, that of genus was 2.8 and of species was 0.07. Similarity analysis demonstrated that seed plant flora of wetland in Sichuan,Yunnan, Tibet and Hubei shared the same origin and the background of species differentiation.
seed plant; species diversity; floristic composition and differentiation; wetland; Sichuan
S718.3
A
1001-3776(2017)01-0006-07
10.3969/j.issn.1001-3776.2017.01.002
2016-09-18;
2016-12-21
四川省第二次湿地资源调查项目(80303-KHS043),四川省第二次全国重点保护野生植物资源调查项目(80303-SRQ012)
王恋,硕士研究生,主要从事植物系统进化和植物生态研究;E-mail:lianwang2016@163.com。通信作者:王娟,讲师,博士研究生,主要从事植物学及生物多样性方面的研究;E-mail:p6309@126.com。
猜你喜欢
疯狂英语·新悦读(2022年7期)2022-11-23
热带作物学报(2022年9期)2022-10-17
北京大学学报(自然科学版)(2022年4期)2022-08-18
云南农业科技(2021年6期)2021-12-30
河北地质(2021年2期)2021-08-21
世界热带农业信息(2019年11期)2019-01-05
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
青春岁月(2017年1期)2017-03-14
安徽农业科学(2015年30期)2015-12-25
湖北农业科学(2014年4期)2014-05-04
