
光周期对‘泰山-橙黄’万寿菊花芽分化和开花的影响1)
2017-06-01 12:27朱玲俐周兰英
东北林业大学学报 2017年4期
朱玲俐 周兰英
(四川农业大学,雅安,625014)
光周期对‘泰山-橙黄’万寿菊花芽分化和开花的影响1)
朱玲俐 周兰英
(四川农业大学,雅安,625014)
以泰山系列橙黄色万寿菊为试材,以雅安地区自然光周期(长日照)为对照,通过遮光处理,研究不同光周期(昼夜8/16 h、10/14 h、12/12 h)对‘泰山-橙黄’万寿菊花芽分化和开花的影响。结果显示:8/16 h处理的花芽分化起始和完成分别在处理后第5天和第2天,较10/14 h处理分别提前了2和5 d,较12/12 h处理分别提前了2和7 d,较对照分别提前了5和16 d;短日照处理使万寿菊开花提前及花期明显延长,其中8/16 h处理效果最明显;与对照相比,短日照处理下万寿菊的鲜花产量显著增加,而盛花期的舌状小花数和花径均显著减小;随着日照时间的减少,株高、基茎、株幅和干质量均有不同程度的减小。以上结果表明,光周期处理可以有效地调节‘泰山-橙黄’万寿菊花芽分化和开花进程,日照时间越少,开花越早,花期越长,而植株长势越弱。
‘泰山-橙黄’万寿菊;光周期;花芽分化;开花
万寿菊(Tageteseresta)性喜温暖湿润和阳光充足的环境,喜湿,耐干旱,因其花大色艳、花期长和花型紧凑等优点,常作草坪点缀、花坛布景和优良的鲜切花材料。近年来,人们对于万寿菊的市场需求越来越大,传统的栽培方式并不能满足其市场需求。已有的研究表明,通过花期调控技术能整年生产菊花,从而满足人们常年对其的消费需求[1]。花期调控的关键在于花芽分化[2],花芽分化和开花是一个十分复杂的形态建成过程,是植物体内各种因素相互作用和协调,以及外界环境共同作用的结果,其中光周期是最重要的影响因素之一[3]。研究光周期诱导花芽分化进程能更好地认识光周期调控花期的机制。迄今,关于万寿菊光周期调控花期的研究报道较少,并且研究不够深入。前人的研究表明,持续的短日照处理可以促使甜万寿菊和色素万寿菊提前开花,延长花期[4-5],并且不同万寿菊品种对光周期的敏感程度有较大的差异。为此,本试验选择具有高观赏价值的‘泰山-橙黄’万寿菊为研究对象,以雅安地区自然光照为对照,通过研究不同光周期处理对其花芽分化和开花的影响,旨在探讨其对光周期敏感程度,以期为万寿菊的光周期调控技术提供理论依据。
1 材料与方法
1.1 材料培养
试验材料选用美国泛美种子公司培育的优良泰山系列橙黄色万寿菊的种子。本试验在四川省雅安市四川农业大学林学院教学基地大棚内进行。2014年4月,种子在25 ℃温水中浸泡8 h,自然晾干后,播种于穴盘中,放入塑料大棚内保温育苗。达到20 d后,植株上盆。缓苗20 d后,选择长势一致的植株集中进行光周期处理。试验采用园土,盆栽种植,统一水肥管理。
1.2 试验方法
自2014年5月17日起,以黑色遮光塑料膜作为遮光材料,对万寿菊进行短日照处理,至植株开花时停止遮光。每处理15株,重复3次。处理如下:每天17:00到翌日09:00进行遮光(16 h),植株受光时间为8 h;每天19:00到翌日09:00进行遮光(14 h),植株受光时间为10 h;每天19:00到翌日07:00进行遮光(12 h),植株受光时间为12 h;对照为当地自然光照长度(5—8月份雅安地区的光照长度可达到14~16 h)。光周期处理后,每隔2~3 d取样一次,剥去展开的叶,采用FAA(V(70%乙醇)∶V(冰醋酸)∶V(37%甲醛)=18∶1∶1)固定,以冰冻切片法制作切片[6],显微镜观察。花芽分化开始所需时间是从处理到总苞分化期(生长点扩大,呈半球形),花芽分化完成所需时间是从处理到花瓣分化期(小花先端出现放射性开裂)。
1.3 生长和开花性状的调查
于处理日(2014年5月17日)起每隔7 d测定各处理株高、基茎、株幅,每次每处理选择12株,取12株平均值。植株开花后,每天记录开花数量,以确定单株开花量(朵/株)、初花期(10%花朵开放)、盛花期(50%花朵开放)、末花期(85%花朵开放)和开花持续时间(末花期—初花期);并测定盛花期各处理的花径(十字交叉测定)和舌状小花数(个/朵)。待植株花朵完全枯萎后,测定植株的地上、地下部分干质量。
1.4 数据分析
数据整理和分析用Excel 2013和SPSS 21.0,显著水平为0.05,数据作图用Sigmaplot 12,并以LSD法进行多重比较分析。
2 结果与分析
2.1 不同光周期对‘泰山-橙黄’万寿菊花芽分化和花期的影响
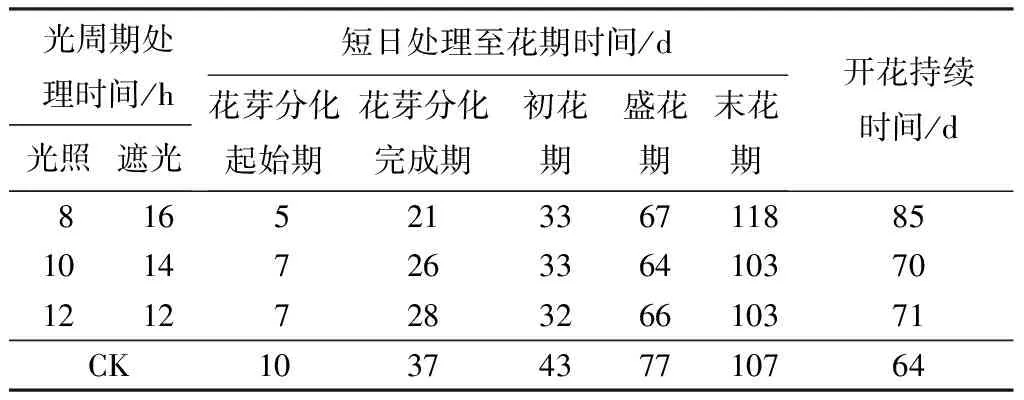
从表1可以看出,‘泰山-橙黄’万寿菊在不同光周期处理下都能正常花芽分化,花芽分化与开花进程规律一致。8/16 h处理的花芽分化开始和完成分别在处理后的第5天和第21天,花芽分化历时16 d;10/14 h处理的花芽分化开始和完成分别在处理后的第7天和第26天,花芽分化历时19 d;12/12 h处理的花芽分化开始和完成分别在处理后的第7天和第28天,花芽分化历时21 d;CK处理的花芽分化开始和完成分别在处理后的第10天和第37天,花芽分化历时27 d。8/16 h、10/14 h、12/12 h处理下,初花期分别较CK提前10、10、11 d;盛花期分别较CK提前10、13、11 d;10/14 h和12/12 h的末花期较CK均提前4 d,但是8/16 h的末花期比CK推迟11 d;开花持续时间分别较对照延长21、6、7 d。
表1 ‘泰山-橙黄’万寿菊在不同光周期处理下花芽分化及花期的规律
光周期处理时间/h光照遮光短日处理至花期时间/d花芽分化起始期花芽分化完成期初花期盛花期末花期开花持续时间/d81652133671188510147263364103701212728326610371CK1037437710764
注:CK(对照)为当地自然光照长度(5—8月份雅安地区的光照长度可达到14~16 h)。
2.2 不同光周期对‘泰山-橙黄’万寿菊开花性状的影响
从表2可以看出,与对照相比,8/16 h、10/14 h和12/12 h的‘泰山-橙黄’万寿菊的鲜花数量均显著增加,但各处理之间的开花量差异不显著;与对照相比,短日照处理下盛花期的舌状小花数量都显著减少,其中,仅8/16 h和12/12 h存在显著差异;盛花期的花朵直径随着日照时间的减少而减小,其中,8/16 h和10/14 h之间差异不显著,都与12/12 h差异显著。
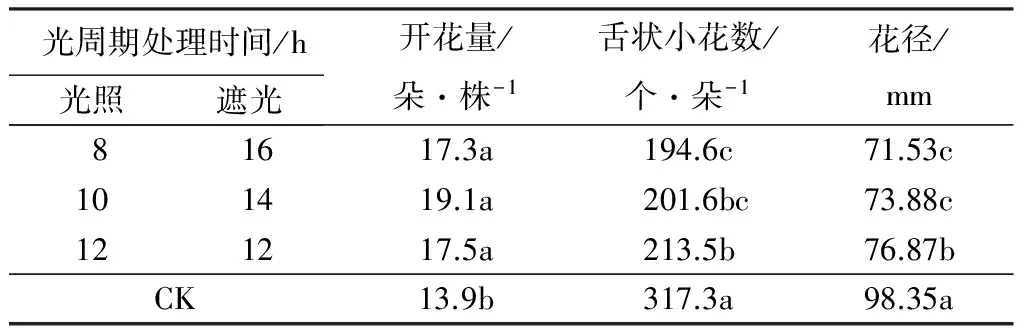
表2 不同光周期处理对‘泰山-橙黄’开花的影响
注:同列不同字母表示在0.05水平上差异显著;CK(对照)为当地自然光照长度(5—8月份雅安地区的光照长度可达到14~16 h)。
2.3 不同光周期对‘泰山-橙黄’万寿菊营养生长和物质积累的影响
由表3可知,‘泰山-橙黄’万寿菊的干质量与日照时间存在正相关关系,即随着日照时间的减少,‘泰山-橙黄’万寿菊地上部分和地下部分的干质量都显著减小。
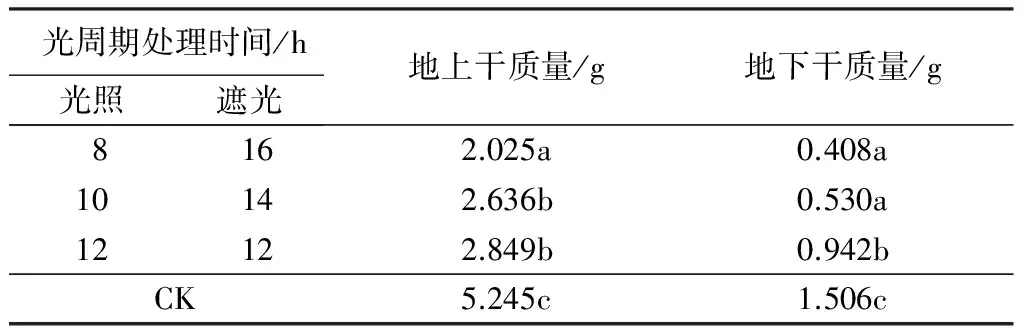
表3 不同光周期处理对‘泰山-橙黄’万寿菊干质量的影响
注:同列不同字母表示不同光周期处理间干质量在0.05水平上差异显著;CK(对照)为当地自然光照长度(5—8月份雅安地区的光照长度可达到14~16 h)。
从表4可以看出,处理前期,‘泰山-橙黄’万寿菊的生长性状差异不显著,随着处理时间增加,短日照处理的影响逐渐显现。处理后期,随着光照时间的减少,‘泰山-橙黄’万寿菊的株高呈现显著下降趋势,与对照相比,短日照处理均显著抑制了植株的高生长;不同处理间基茎差异达到显著水平,与对照相比,12/12 h处理对基径的影响较小,8/16 h和10/14 h则显著抑制了植株的径生长;‘泰山-橙黄’万寿菊的株幅逐渐下降,与对照相比,短日照处理显著抑制了植株的株幅生长。
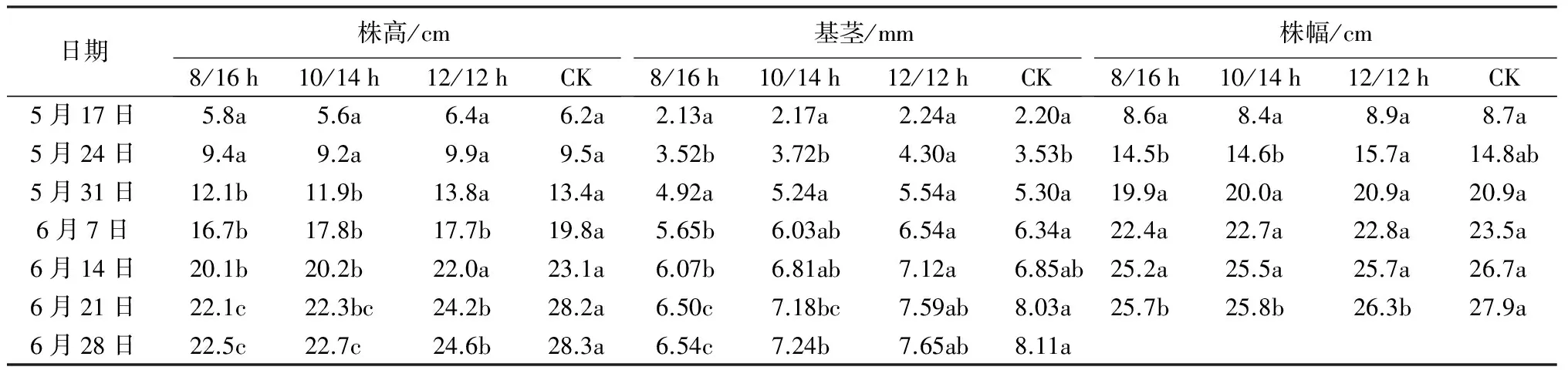
表4 不同光周期处理对‘泰山-橙黄’万寿菊生长性状的影响
注:同行不同字母表示同一时间不同光周期处理间形态指标在0.05水平上差异显著。
3 结论与讨论
本研究发现,8/16 h处理下花芽分化开始最早,其次为10/14 h处理,最后为CK,这与毛洪玉等[7]对万寿菊花芽分化的研究结果一致。光周期效应取决于暗期长度而非光期长度或光暗期之比,即暗期长度为光周期的感应时期[8]。说明‘泰山-橙黄’万寿菊对短日照较敏感,即万寿菊开花需要一定黑夜的积累。经过短日照处理后,虽然万寿菊体内激素水平很快就发生变化,但并不立即出现花芽形态分化,而是经过5~7 d后才开始出现,这与林贵玉等[2]和郑成淑等[9]研究得到的短日照下菊花花芽分化规律一致,说明‘泰山-橙黄’万寿菊的花芽型态分化需要短日照的诱导过程,在这个过程中体内发生复杂的生理生化反应,其中包括内源多胺在内的多种生理生化物质和内源激素的代谢过程,当这些物质水平达到一定程度或一定比例关系时,植物体才能由营养生长转变为生殖生长[10]。不同光周期处理间花芽分化历时也有明显的差异,8/16 h处理花芽分化历时16 d,较10/14 h提前3 d,较12/12 h处理提前5 d,较CK处理提前11 d。这与马芳芳[11]和杨娜等[12]的研究结果均不同,上述差异主要是因为不同研究间植物材料、处理方法和栽培方法等不同造成的。研究还发现,缩短日照时间对‘泰山-橙黄’万寿菊的花期有不同程度的提前及延长,这是由于短日照(长黑夜)促进了花芽分化诱导过程,而花芽分化和开花进程规律一致,从而使万寿菊的花期提前及延长[7]。以上结果表明,‘泰山-橙黄’万寿菊是典型短日照植物,日照时数越短,越有利于其花芽分化和开花。
通过对开花性状的观测表明,短日照条件下,舌状小花数显著减少,花径显著减小,与任旭琴[13]的研究结果相吻合;相对于CK,短日照处理显著提高了鲜花产量,这可能与短日照(长黑夜)更容易诱导植物体生长枝由营养生长向生殖生长转换有关。因此,短日照处理有利于提高万寿菊的鲜花产量。研究表明,不同光周期处理间万寿菊外观形态有较大差异,尤其表现在株高和干重上。短日照虽然促进了‘泰山-橙黄’万寿菊的生殖生长,但抑制了其营养生长,导致其生长速度减慢,株高变矮,基径和株幅减小,同时也抑制了其物质积累,导致其干质量显著降低,这与周志凯等[14]对万寿菊的研究结果吻合。这可能是由于短日照处理对其光合作用、营养元素吸收、激素合成和信号物质的表达等生理生化代谢过程产生了不良影响[15]。
综上所述,光周期处理可以有效地调节‘泰山-橙黄’万寿菊花芽分化和开花进程,日照时数越少,开花越早,花期越长,而植株长势越弱。本研究结果为该地区万寿菊的光周期调控精准化生产提供了一定的理论依据。此外,本试验仅明确了光周期对万寿菊开花效应的影响,而其他环境因子(如光强、温度等)的改变是否也能达到调控其开花效应的目的,仍需在今后的工作中作进一步的研究,以便更好地指导万寿菊周年生产。
[1] YAMAMOTO S, MISUMI M, NAWATA E. Effects of photoperiod on vegetative growth, flowering and fruiting ofCapsicumfrutescensL. andC.annuumL. in Japan[J]. Environment Control in Biology,2008,46(1):39-47.
[2] 林贵玉,郑成淑,孙宪芝,等.光周期对菊花花芽分化和内源激素的影响[J].山东农业科学,2008(1):35-39.
[3] WARNER R M. Temperature and photoperiod influence flowering and morphology of fourPetuniaspp[J]. Hortscience A Publication of the American Society for Horticultural Science,2010,45(3):365-368.
[4] 刘宝勇.光周期诱导促进甜万寿菊(Tageteslucida)花期初探[J].中国园艺文摘,2010,26(12):44-77.
[5] 贾兰虹,张雪岩,吕桂菊.色素万寿菊开花光周期调控规避病害增产效果研究[J].北方园艺,2014(17):83-86.
[6] 刘剑锋,程云清,阎秀峰,等.植物冰冻切片技术的改进[J].南京林业大学学报(自然科学版),2006,30(3):128-130.
[7] 毛洪玉,顾钊宇,祝朋芳.不同光周期处理对菊花‘C029’花芽分化及开花的影响[J].西北植物学报,2010,30(10):2074-2080.
[8] 洪艳,陈之琳,戴思兰.切花菊‘丽金’响应光照诱导成花特性研究[J].北京林业大学学报,2015,37(3):234-234.
[9] 郑成淑,孙宪芝,王文莉,等.光周期对菊花花芽分化及其叶片和芽内源多胺含量的影响[J].西北植物学报,2008,28(7):1349-1353.
[10] 常海龙,王勤南,周峰,等.光周期诱导甘肃花芽分化及成花逆转过程中生理代谢的变化[J].植物生理学报,2014,50(7):1045-1052.
[11] 马芳芳.光周期调控‘流星’翠菊开花的初步研究[D].武汉:华中农业大学,2009.
[12] 杨娜,郭维明,陈发棣,等.光周期对秋菊品种‘神马’花芽分化和开花的影响[J].园艺学报,2007,34(4):965-972.
[13] 任旭琴,周志凯,王连臻,等.叶面施肥和遮光对孔雀草生长和开花的影响[J].甘肃农业大学学报,2010,45(5):96-99.
[14] 周志凯,任旭琴.遮光对万寿菊营养生长和开花的影响[J].北方园艺,2010(5):107-109.
[15] 张春燕,马芳芳,王琴,等.2种光周期条件下‘日光’翠菊的生长发育特性[J].华中农业大学学报,2014,33(1):35-38.
Effect of Photoperiod onTageteseresta‘Mount Tai Orange’ Floral Bud Differentiation and Flowering//
Zhu Lingli, Zhou Lanying(Sichuan Agricultural University, Ya’an 625014, P. R. China)//
Journal of Northeast Forestry University,2017,45(4):33-35,59.
We studied the effects of different photoperiod (8/16 h, 10/14 h, 12/12 h, natural light for control) on floral bud differentiation and flowering withTageteseresta‘Mount Tai Orange’. The beginning of floral bud differentiation of 8/16 h was 5 d after treatments, which advanced by 2 d, 2 d and 5 d than that of 10/14 h, 12/12 h and CK. The ending of floral bud differentiation of 8/16 h was found 21 d after treatments, which advanced by 5 d, 7 d and 16 d than that of 10/14 h, 12/12 h and CK, respectively. All short-day treatments could significantly expedite flowering and extend flowering duration, and 8/16 h treatment had the most effect on flowering. Flowering amount increased significantly under the continuous short-day conditions, but tongue florets and flower diameter decreased significantly. Height, basal diameter, plant width and dry weight were declined with the shortening of lights to some extents. The photoperiod treatment could effectively adjust the floral bud differentiation and flowering ofT.eresta‘Mount Tai Orange’.
Tageteseresta‘Mount Tai Orange’; Photoperiod; Floral bud differentiation; Flowering
朱玲俐,女,1992年12月生,四川农业大学林学院,硕士研究生。E-mail:1529134784@qq.com。
周兰英,四川农业大学林学院,教授。E-mail:949460359@qq.com。
2016年6月13日。
S681.9;Q945.43
1)四川省农作物育种攻关项目(2011NZ0098-10)。
责任编辑:潘 华。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
黑龙江粮食(2022年6期)2022-11-23
中国水土保持(2022年2期)2022-04-07
云南农业(2021年9期)2021-09-24
落叶果树(2020年3期)2020-06-17
河北果树(2020年1期)2020-02-09
浙江农业学报(2019年7期)2019-08-14
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
现代农业(2016年5期)2016-02-28
