
海拔和坡向对北亚热带檫木天然次生林生长、空间结构和树种组成的影响1)
2017-06-01 12:27孙洪刚姜景民万志兵
东北林业大学学报 2017年4期
孙洪刚 姜景民 万志兵
(中国林业科学研究院亚热带林业研究所,杭州,311400) (黄山学院)
海拔和坡向对北亚热带檫木天然次生林生长、空间结构和树种组成的影响1)
孙洪刚 姜景民 万志兵
(中国林业科学研究院亚热带林业研究所,杭州,311400) (黄山学院)
基于对北亚热带檫木天然次生林的野外调查和数据分析,研究了海拔和坡向对檫木生长、空间结构和树种组成等方面的影响。结果表明:在海拔1 000 m以上山地,檫木的胸径和树高生长量及胸径/树高均差于海拔1 000 m以下的山地,人工林造林地选择宜在海拔1 000 m以下;随阳坡→半阳坡→半阴坡→阴坡坡位变化,檫木生长量和胸径/树高呈逐渐减小趋势。阳坡和半阳坡差异不显著,是理想的檫木造林坡向;檫木纯林或者混交林可以采用均匀造林模式。在海拔500 m以下阳坡,可以实现混交增产造林树种的混交伴生树种为杉木、香樟和枫香;在海拔500 m以下半阳坡,混交伴生树种为杉木、亮叶桦、枫香和枳椇;在海拔500~1 000 m的阳坡山地,混交伴生树种为甜槠、枫香和木荷;在海拔500~1 000 m的半阳坡山地,混交伴生树种为木荷、麻栎、杜英、枫香和紫茎。初步解决了北亚热带山地檫木分布区划分、空间结构变化及互利混交树种等问题,可以为檫木人工造林的林地选择、造林模式和混交伴生树种选取等方面提供理论依据。
北亚热带;檫木;海拔;坡向;空间结构
檫木(Sassafrastsumu(Hemsl.) Hemsl.)是樟科(Lauraceae)落叶大乔木,散生于亚热带丘陵和低山地区,具有树干通直圆满、木材节疤少、材质细腻、纹理美观和具芳香气味等优点。20世纪70—80年代,江苏、浙江、安徽、江西、湖南、福建、四川、云南、贵州等亚热带省份曾进行过大规模的人工造林[1-5]。但大部分檫木人工林由于造林地瘠薄导致林分灌丛化,成活不成林;造林密度过大,林分过早进入种内竞争阶段,出现生长早期衰退、人工纯林结构单一、日光直射树干、引起树皮开裂等问题,给造林生产单位带来重大经济损失。近年来,随着人们生活水平的提高,家私家具、室内装修、工艺品等行业对木材的视觉和嗅觉等特性提出更高要求,由于檫木在阔叶用材中具有独特的木材色泽、纹理和芳香气味,再次受到人们的重视。但由于目前檫木人工林集约栽培技术不成熟,原木产量低下,而檫木天然次生林被划分为国家公益林保护范畴,严格禁止采伐,从而导致檫木的需求矛盾进一步突出。随着木材需求多样化和需求量的不断提高,世界上主要原木生产出口国家和地区正积极开展阔叶树的培育机理和技术研究。如通过研究海拔和坡向等非生物因素引起的环境梯度变化与树种分布的关系,确定树种的地理最适分布区[6];研究不同生长阶段混交林内种间关系能否促进林分增产,以确定最佳混交树种种类和比例[7];分析限制用材林生长的因素,通过混交、疏伐等培育措施促进混交林内种间合作关系,提高对限制性资源的捕获和利用效率,解除其对生长的限制[8]。分析以上文献不难发现:在明确限制阔叶树种生长的生境因子基础上,通过宜林地选择、最适混交造林树种选取,以及通过对林分空间进行调控改善种间关系等,以实现提高木材产量的目标,是阔叶用材林培育机理研究的核心问题之一。因此,笔者以生长在不同海拔和坡向的檫木天然次生林为对象,通过研究檫木林分生长、混交树种种类和空间结构等内容,分析檫木天然次生林生长与环境梯度变化的关系,旨在为檫木人工林定向培育过程中在造林地选取、混交树种选择,以及林分空间结构调控等方面提供理论依据。
1 研究地概况
野外调查样地设置在安徽省黄山市黄山风景区,中心位置地理坐标118°11′E、30°10′N,海拔1 864.8 m,属于亚热带季风气候,年日照时间1 647~1 743 h,年辐射总量45×108~48×108J/m2;年均气温7.8 ℃,极端最高气温40.6 ℃(2000年),极端最低气温-15.5 ℃(1971年),≥10 ℃积温5 240~6 678 ℃;年均降水量2 394.5 mm,无霜期210~240 d。土壤属亚热带黄红土壤带。植被分布具有典型的山地垂直地带性。在海拔500 m以下,植被群落的建群种主要为杉木(Cunninghamialanceolata(Lamb.) Hook.)、马尾松(PinusmassonianaLamb.)、枫香(LiquidambarformosanaHance)、苦槠(Castanopsissclerophylla(Lindl.) Schottky.)等乔木树种及人工的毛竹林(Phyllostachyspubescens)群落;伴生树种多为茅栗(Castaneaseguinii)、化香树(Platycaryastrobilacea)、檵木(Loropetalumchinense)等树种;林下草本主要为乌蕨(Stenolomachusanum(Linn.) Ching)、凤丫蕨(Coniogrammejaponica(Thunb.) Diels)、井栏边草(Pterismultifida)及五节芒(Miscanthusfloridulus(Labill.) Warb. ex Schum. et Laut.)等。在南坡从海拔500~1 000 m,北坡从海拔450~950 m的森林群落,青冈栎(Cyclobalanopsisglauca(Thunb.) Oerst.)、甜槠(Castanopsiseyrei(Champ. ex Benth.) Tutch.)、小叶青冈(Cyclobalanopsismyrsinifolia(Blume) Oersted)等为天然群落的建群种,紫楠(Phoebesheareri(Hemsl.) Gamble)、细叶香桂(Cinnamomumsubavenium)、交让木(DaphniphyllummacropodumMiq.)等为伴生树种;林下灌木主要为羊踯躅(Rhododendronmolle(Blume) G. Don),米饭花(VacciniuimmandarinorumDiels)、山苍子(Litseacubeba(Lour.) Pers.)、盐肤木(RhuschinensisMill.)等;草木植物有麦冬(Ophiopogonjaponicus(Linn. f.) Ker-Gawl.)、淡竹叶(Lophatherumgracile)等。南坡从海拔1 000 m以上,北坡从海拔850 m以上,细叶青冈(Cyclobalanopsisgracilis(Rehd. et Wils.) Cheng et T. Hong)、交让木、枫香、糙叶树(Aphanantheaspera(Thunb.) Planch.)等为群落优势种;林下灌木有连蕊茶(Camelliacuspidata(Kochs) Bean)、胡颓子(ElaeagnuspungensThunb.)、野鸦椿(Euscaphisjaponica(Thunb.) Dippel)、撒八仙(Hydangeaumbellata)等;草木植物有油点草(TricyrtismacropodaMiq.)、日本鹿蹄草(Pyrolajaponica)、庐山石韦(Pyrrosiasheareri)等。
2 研究方法
2.1 野外调查与测定
从黄山山脚(海拔100 m)至山顶(海拔1 200 m)檫木天然分布区,分别在阳坡、半阳坡、半阴坡和阴坡4个坡向,海拔高度每上升50 m,选择以檫木为中心和对檫木生长具有影响的最近4株相邻木作为调查单元[9],并设置30 m×30 m临时样地。分别测定檫木和相邻木的胸径、树高、枝下高、冠幅,以及檫木与相邻木、相邻木与相邻木之间的距离,记录相邻木拉丁种名。利用三角公式分别计算以檫木为中心与另外2株相邻木间的夹角,用手持GPS和地质罗盘仪测定檫木所在地理位置的海拔和坡向信息。
2.2 林分生长指标
利用野外调查檫木单木的胸径、树高、枝下高、冠幅等数量生长指标,分别计算不同海拔和坡向条件下各生长指标的平均值,作为描述檫木天然林分生长特征的指标。
2.3 林分结构和种间关系指标
采用角尺度参数描述檫木次生林的分布特征[10]。当角尺度<0.5时,相邻竞争木为均匀分布;当0.5≤角尺度<0.6时,相邻竞争木为随机分布;当角尺度≥0.6时,相邻竞争木为团状分布。
采用混交度(1-隔离度)描述檫木天然次生林中树种混交程度[11]。
林分种间关系采用竞争指数和生长量双指标进行判定[8-9]。如果相邻檫木间的竞争指数大于檫木与非檫木种间的竞争指数,并且相邻檫木生长量小于檫木与其它相邻树种生长量,则该非檫木树种与檫木之间为互利合作关系,可以作为檫木混交林的造林树种;反之,则为竞争关系,不适合于檫木混交造林。
2.4 海拔高度和坡向划分
依据北亚热带植被分布的地形规律,将海拔高度划分为3个梯度:≤500 m(丘陵)、500~1 000 m(低山)和≥1 000 m(中山),比较海拔高度对檫木生长和林分空间结构的影响[12]。
以正北方为0°,顺时针递增,正东方为90°,正南方为180°,正西方为270°。其中,(315°,45°]之间坡向为阴坡;(45°,135°]之间坡向为半阳坡;(135°,225°]之间坡向为阳坡;(225°,315°]之间为半阴坡。并以此为据,进行不同坡向檫木天然次生林生长和空间结构的统计分析。
2.5 数据处理
采用SPSS18.0软件中广义线性模型进行方差分析,比较不同海拔和坡向对檫木生长和空间结构影响的差异性;对于同一因素不同水平间的数量指标,采用邓肯多重比较判断是否具有差异显著性(P<0.05)。部分数据和图表采用Excel 2010进行处理。
3 结果与分析
3.1 海拔高度对林分生长的影响
表1结果表明:海拔500 m以下和500~1 000 m两个海拔梯度的天然次生林中生长的檫木,两者间的胸径和树高差异并不显著(P>0.05);但均与生长在海拔1 000 m以上山地的檫木存在显著差异(P<0.05)。枝下高和冠幅随海拔高度增加而降低,胸径/树高和冠高/树高生长指标均随海拔高度增加而增加,且3个海拔梯度间差异显著(P<0.05)。这意味着:在海拔1 000 m以下的天然次生林中,檫木生长并没有受到海拔高度的影响。值得注意的是,在海拔高度小于500 m天然次生林中生长的檫木,其胸径和树高均小于海拔500~1 000 m天然次生林中生长的檫木。这主要是由于小于500 m海拔的山地人类活动频繁,生长良好的散生檫木会被生产单位和农户林下抚育时将檫木作为杂木清除,以及天然次生林改种油茶、山核桃、毛竹等经济林种,导致存留檫木多数为生长势衰退的过熟木或多代萌蘖幼龄林。而在500~1 000 m山地多数划归成公益林严禁采伐,同时檫木的早期速生特性,使其在资源竞争中处于优势地位,生长势旺盛,导致生长量大于海拔<500 m的天然次生林中的檫木。
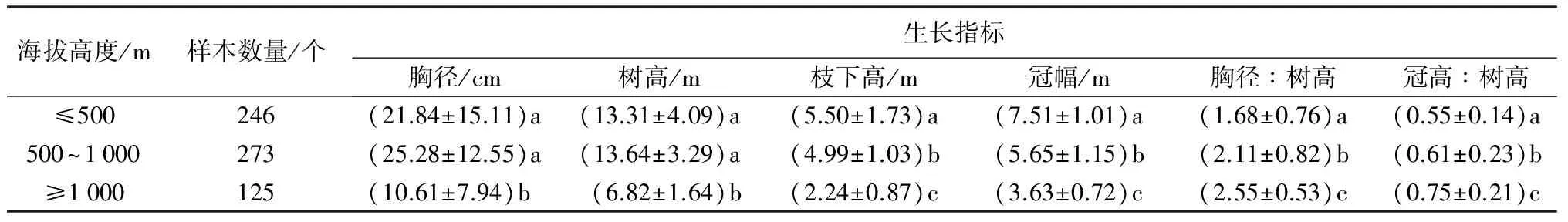
表1 不同海拔高度天然次生林檫木的生长指标
注:同列不同小写字母表示在P<0.05水平差异显著。
3.2 海拔高度对林分空间结构的影响
表2统计结果显示:在海拔高度小于1 000 m的天然次生林中,檫木天然次生林树木为均匀分布,不同树木与檫木之间保持一定的均匀间距;而这种分布方式在自然情况下极为罕见,多见于人工栽培的有一定株行距的植物群落中。在海拔大于1 000 m的天然次生林中,檫木的相邻竞争木呈随机分布,不同树种在不同取样檫木周围出现概率相同。
混交度度数值越大,表明同种个体聚生的可能性越小。在所调查的檫木天然次生林中,混交度随海拔升高而降低,不同海拔高度之间的混交度数值具有显著差异(P<0.05)(表2)。这表明随海拔升高,檫木同种个体聚生可能性增大,混交树种种类减少。
种间竞争数值越大,相邻竞争木对参照木的上方遮盖和侧方挤压程度越强。可知:随海拔升高,竞争强度下降,不同海拔之间竞争强度差异显著(P<0.05)。但总体来说,相邻竞争木对檫木的遮盖和挤压作用很小。
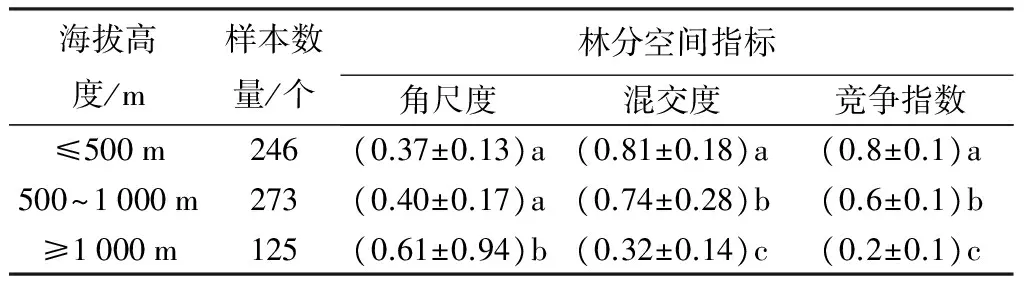
表2 不同海拔高度天然次生林檫木林分空间分布
注:同列不同小写字母表示在P<0.05水平差异显著。
3.3 坡向对林分生长的影响
树种对光照的适应性决定了其分布的坡向[13-14]。檫木为北亚热带天然次生林中的阳性建群树种,不同坡向的光照强度决定了其生长、分布和适应性等状况。对阳坡、半阳坡、半阴坡和阴坡4个坡向的檫木生长的统计分析表明:阳坡和半阳坡为檫木最适生长坡向,胸径和树高生长量最大,二者之间差异不显著(P>0.05);半阴坡次之,阴坡生长量最小(表3)。随阳坡→半阳坡→半阴坡→阴坡光照强度逐渐降低,枝下高和冠幅也呈现相应减小趋势;而胸径/树高和冠高/树高两个相对指标则表现为随阳坡→半阳坡→半阴坡逐渐增大,而在阴坡又呈降低趋势,但仍旧小于阳坡和半阳坡数值。胸径/树高比值越大,檫木尖削度越大,出材量降低;冠高/树高变大,冠高变长会导致树干自然整枝长度变短,原木节疤增多。值得注意的是,阴坡檫木树高和胸径等生长指标小于阳坡,但胸径/树高和冠高/树高两个相对指标却大于或略小于阳坡,这说明阴坡檫木虽然生长量不如阳坡,但干形和材性可能优于阳坡。
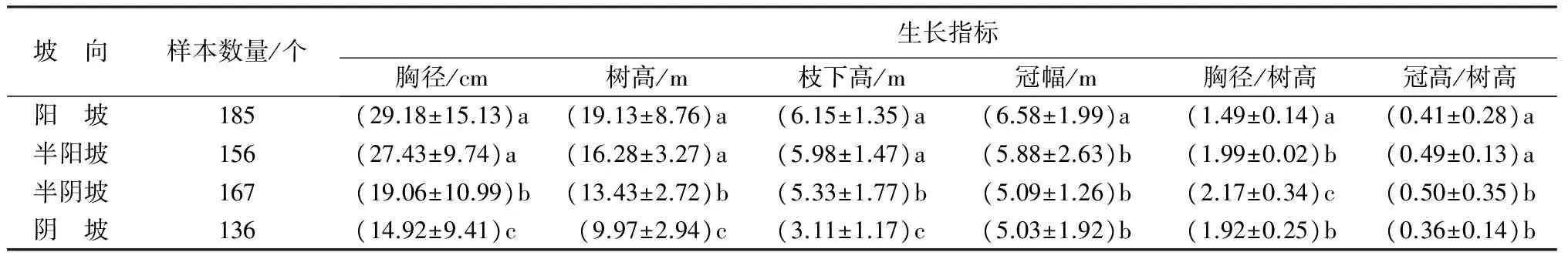
表3 不同坡位天然次生林生长比较
注:同列不同小写字母表示在P<0.05水平差异显著。
3.4 坡向对林分空间结构的影响
不同坡向的檫木天然次生林林分空间结构特征值具有不同差异(表4)。不同坡向檫木天然次生林的角尺度数值随阳坡→半阳坡→半阴坡→阴坡次序呈现逐渐增大趋势,阳坡和半阳坡间角尺度以及半阴坡和阴坡角间尺度均没有显著差异(P>0.05),且都小于0.5,表明不同坡向与檫木混交林内的不同树种都属于均匀分布。不同坡向檫木相邻竞争木的树种隔离度指数和竞争指数随阳坡→半阳坡→半阴坡→阴坡次序均呈现逐渐增大趋势。但阳坡和半阳坡之间的竞争指数差异不显著(P>0.05);竞争指数在阳坡、半阳坡和半阴坡之间差异不显著(P>0.05),但均与阴坡存在显著差异。
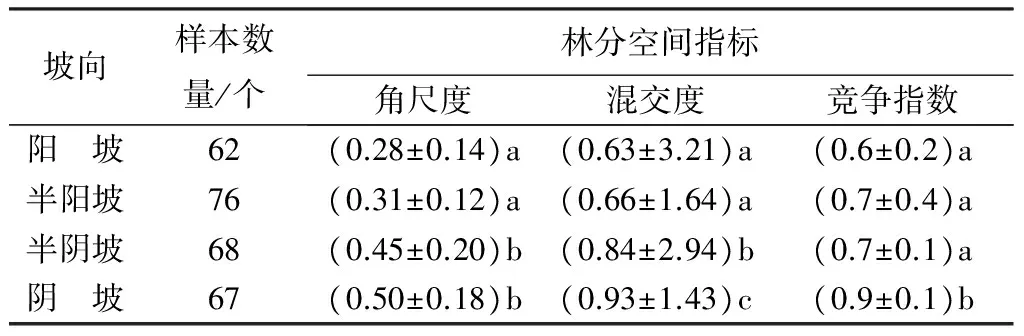
表4 不同坡位天然次生林檫木林分空间分布
注:同列不同小写字母表示在P<0.05水平差异显著。
3.5 海拔和坡向对混交伴生树种分布的影响
不同海拔和坡向会引起生境条件的养分和光照差异,导致不同生物学特性树种的分布区不同。在海拔500 m以下,杉木、马尾松和柳杉(CryptomeriafortuneiHooibrenk ex Otto et Dietr.)等常绿针叶树种,木荷(SchimasuperbaGardn. et Champ.)、香樟(Cinnamomumcamphora(L.) Presl)、苦槠、毛竹和甜槠等常绿阔叶树种,檫木、枫香、响叶杨(PopulusadenopodaMaxim.)、山苍子、麻栎(QuercusacutissimaCarr.)、钩栗(CastanopsistibetanaHance)、锥栗(Castaneahenryi(Skan) Rehd. et Wils.)、泡桐(Paulownia)、枳椇(HoveniaacerbaLindl.)、李(PrunussalicinaLindl.)和亮叶桦(BetulaluminiferaH. Winkl.)等落叶阔叶树种,在阳坡和半阳坡以较高频率与檫木混生。在海拔500~100 m,常绿针叶树杉木、马尾松和柳杉,常绿阔叶树种甜槠、木荷、香樟、毛竹、红楠、杜英(ElaeocarpusdecipiensHemsl.)和柃木(EuryajaponicaThunb),落叶阔叶树种亮叶桦、响叶杨、刺楸(Kalopanaxpictus(Thunb.)、麻栎、泡桐、化香树、紫茎(StewartiasinensisRehd. Et Wils.)、枫香和锥栗,多于檫木混生。在海拔1 000 m以上,黄山松(PinustaiwanensisHayata)在任一坡向都有分布;而混生树种如米心水青冈(FagusenglerianaSeem.)、色木槭(AcermonoMaxim.)、灯台树(CornuscontroversaHemsl)、华东椴(Tiliajaponica(Miq.) Simonk)、橄榄槭(AcerolivaceumFang et P. L. Chiu)、多花泡花树(MeliosmamyrianthaSieb. et Zucc.)、四照花(Dendrobenthamiajaponica(DC.) Fang var. chinensis)等均为落叶阔叶树种(表5)。随着海拔升高,与檫木混生常绿针叶林树种由山脚至山顶均有分布,阔叶混生树种种类则表现为低海拔地区主要以常绿阔叶树种为主,随海拔增加常绿阔叶树种种类降低而落叶阔叶树种增加的趋势。

表5 不同海拔和坡向对檫木竞争木树种分布
檫木天然次生林不同树种混交随海拔和坡向的变化而有所不同(图1)。在海拔500 m以下的阳坡山地,种间竞争指数大于檫木种内竞争指数的树种为毛竹。这表明檫木如与毛竹混交,由于二者间存在明显的竞争关系,将会抑制檫木生长。种间竞争指数小于檫木种内竞争指数的树种有:杉木、马尾松、香樟、响叶杨、山苍子、枫香、柳杉和木荷,以上树种在与檫木混生时竞争强度小于檫木纯林,这很有可能是上述树种与檫木混交生长时,二者之间是互利合作关系。在对这些树种生长量(平均胸径)与檫木生长量(平均胸径)比较时发现:比檫木胸径大的树种为杉木、香樟和枫香,竞争强度小于檫木纯林,但平均生长量(平均胸径)却大于檫木纯林。这种情况就说明上述3个树种与檫木混交时具有互利合作关系,竞争强度减小,生长量增大。因此,在海拔500 m以下的阳坡山地,可以选取杉木、香樟和枫香与檫木混交造林,利用种间互利合作关系,提高生长量。在海拔500以下的半阳坡山地,种间竞争指数大于檫木种内竞争指数的树种为响叶杨、泡桐、柳杉和马尾松,种间竞争指数小于檫木种内竞争指数的树种有香樟、杉木、亮叶桦、枫香、李、枳椇和油茶。在前述7个树种中,平均胸径大于檫木的树种为枫香、杉木、亮叶桦和枳椇。这说明在海拔500 m以下半阳坡山地,可选择枫香、杉木、亮叶桦和枳椇与檫木进行混交造林。在500~1 000 m的阳坡山地,种间竞争指数大于檫木种内竞争指数的树种为马尾松、泡桐、麻栎和刺楸,种间竞争指数小于檫木种内竞争指数的树种为杉木、枫香、木荷和甜槠。在前述4个树种中,平均胸径大于檫木平均胸径的树种为甜槠、木荷和枫香。因此,在500~1 000 m的阳坡山地,甜槠、木荷和枫香是与檫木混交的理想树种。在500~1 000 m的半阳坡山地,种间竞争指数大于檫木种内竞争指数的树种为泡桐,种间竞争指数小于种内竞争指数的树种有毛竹、木荷、化香树、麻栎、杜英、杉木、甜槠、马尾松、亮叶桦、红楠、柃木、枫香、柳杉和紫茎。在前述14个树种中,平均胸径大于檫木平均胸径的树种为枫香、杜英、麻栎和木荷。因此,在500~1 000 m的半阳坡山地,适宜与檫木混交的造林树种为枫香、杜英、麻栎和木荷。
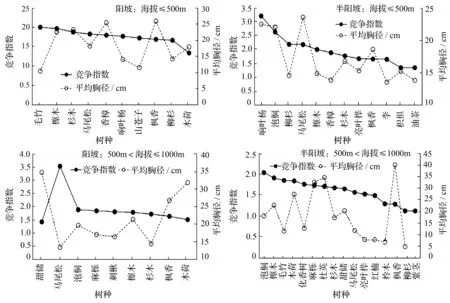
图1 檫木天然次生林混交树种种间关系
4 结论
随海拔高度升高,温度、水分、土壤等自然环境因子变化,形成环境梯度胁迫,从而影响植物生长、适应性和分布等[15-17]。北亚热带天然林中檫木生长,以海拔1 000 m为界限,生长在海拔1 000 m以下的檫木,胸径、树高和枝下高等指标的生长量均大于海拔1 000 m以上的檫木,但胸径/树高和冠高/树高相对指标值小于海拔1 000 m以上的檫木,意味着北亚热带1 000 m以下山地檫木生长量大,胸径/树高和冠高/树高比值较小,可以得到干形良好和材质优良的檫木木材。这主要是由于随海拔升高,温度降低,土壤中水分和养分无法供给生长和繁殖需求,没有成熟老化的组织也会发生冻害,从而限制了树木在高海拔地区的存活和生长[18-19]。
天然林中不同树种的分布方式可分为均匀分布、随机分布和团状分布等3种方式,这主要与树种自身生长特性、种间关系和生境条件有关[20]。天然林中树种的生长分布方式对人工造林培育模式的选择具有重要参考作用。檫木天然次生林在海拔1 000 m以下为均匀分布,这与人工机械造林的树种分布模式相一致;在海拔大于1 000 m天然次生林中呈随机分布,这种模式是天然次生林树木分布的普遍现象。随海拔升高,同种个体聚生可能性越小,多数为混交林分,这一趋势与多数高山树种多样性分布相一致。人工造林可以依据海拔高度变化,选择相应适生混交树种。竞争指数随海拔高度升高而下降,这是由于低海拔与檫木生长生境需求接近的树种较多,从而导致的竞争强度较大。
坡向变化影响树木生长、干形和材质[21]。阳坡和半阳坡檫木胸径和树高生长量最大,树干尖削度小最小,无节干材长度最大,为檫木最适生长坡向。
坡向和海拔共同作用决定了适合于檫木混交的造林树种[14]。在北亚热带海拔500 m以下阳坡山地,适合与檫木混交树种为杉木、香樟、和枫香;在海拔500 m以下半阳坡山地,适合与檫木混交的树种为杉木、亮叶桦、枫香和枳椇;在海拔500~1 000 m的阳坡山地,可以选择甜槠、枫香和木荷与檫木混交;在海拔500~1 000 m的半阳坡山地,木荷、麻栎、杜英、枫香和紫茎都可以与檫木形成混交林。
[1] 庄平忠.黔中地区檫木造林试验初报[J].贵州林业科技,1981(4):14-20.
[2] 刘安兴.缺乏常量元素对檫树苗生长的影响[J].浙江林业科技,1983(2):10-13.
[3] 吴振中,何怡熙.三明莘口檫树林稠查研究初步报告[J].福建林学院学报,1963(3):77-88.
[4] 张焕朝,俞元春.杉木、檫木、柏木等林分下土壤微量元素状况[J].南京林业大学学报,1995,19(2):6-12.
[5] 姜培坤,徐秋芳,钱新标,等.杉木檫树根际土壤磷素研究[J].浙江林学院学报,1995,12(3):242-246.
[6] CHOLER P, MICHALET R, CALLAWAY R M. Facilitation and competition on gradients in alpine plant communities[J]. Ecology,2001,82(12):3295-3308.
[7] TOÏGO M, VALLET P, PEROT T, et al. Overyielding in mixed forests decreases with site productivity[J]. Journal of Ecology,2015,103(2):502-512.
[8] FORRESTER D I, KOHNLE U, ALBRECHT A T, et al. Complementarity in mixed-species stands ofAbiesalbaandPiceaabiesvaries with climate, site quality and stand density[J]. Forest Ecology and Management,2013,304(4):233-242.
[9] 惠刚盈,胡艳波,赵中华,等.基于交角的林木竞争指数[J].林业科学,2013,49(6):68-73.
[10] 惠刚盈,KLAUS von GADOW, MATTHIAS A.角尺度:一个描述林木个体分布格局的结构参数[J].林业科学,1999,35(1):37-42.
[11] 惠刚盈,胡艳波,赵中华.基于相邻木关系的树种分隔程度空间测度方法[J].北京林业大学学报,2008,30(4):131-134.
[12] 中国科学院《中国自然地理》编辑委员会.中国自然地理总论[M].北京:科学出版社,1985.
[13] PRÉVOST M, RAYMOND P. Effect of gap size, aspect and slope on available light and soil temperature after patch-selection cutting in yellow birch-conifer stands, Quebec, Canada[J]. Forest Ecology and Management,2012,274(8):210-221.
[14] GETZIN S, WIEGAND K. Asymmetric tree growth at the stand level: Random crown patterns and the response to slope[J]. Forest Ecology and Management,2007,242(2/3):165-174.
[15] MYERS J A, KITAJIMA K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest[J]. Journal of Ecology,2007,95(2):383-395.
[16] SPANN T M, BEEDE R H, JONG T M. Seasonal carbohydrate storage and mobilization in bearing and non-bearing pistachio (Pistaciavera) trees[J]. Tree Physiology,2008,28(2):207-213.
[17] SIMONE F, SEBASTIAN L, CHRISTIAN K. Moving beyond photosynthesis: from carbon source to sink-driven vegetation modeling[J]. New Phytologist,2014,201(4):1086-1095.
[18] HOCH G, SIEGWOLF R T W, KEEL S G, et al. Fruit production in three masting tree species does not rely on stored carbon reserves[J]. Oecologia,2013,171(3):653-662.
[19] IMAJI A, SEIWA K. Carbon allocation to defense, storage, and growth in seedlings of two temperate broad-leaved tree species[J]. Oecologia,2010,162(2):273-281.
[20] 陈大坷,周晓峰,赵惠勋,等.天然次生林四个类型的结构、功能及演替[J].东北林学院学报,1982(2):1-20.
[21] MOREIA X, ZAS R, SAMPEDRO L. Genetic variation and phenotypic plasticity of nutrient re-allocation and increased fine root production as putative tolerance mechanisms inducible by methyl jasmonate in pine trees[J]. Journal of Ecology,2012,100(3):810-820.
Effects of Altitude and Aspect on Stand Growth, Spatial Structure and Tree species Composition with theSassafrastsumuNatural Secondary Forest in the Northern Subtropical Zone, China//
Sun Honggang, Jiang Jingmin(Institute of Subtropical Forestry, Chinese Academy of Forestry, Hangzhou 311400, P. R. China); Wan Zhibing(Huangshan University)//
Journal of Northeast Forestry University,2017,45(4):8-13.
By the field investigation and data analysis with Mount HuangSassafrastsumu(Hemsl.) Hemsl. natural secondary forest in the northern subtropical zone, we studied the stand growth, spatial structure and tree species composition of the different altitude and aspect. The diameter at breast height (DBH) and total height (TH) of stand growth and DBH/TH below 1 000 m elevation are superior to above 1 000 m elevation. With aspect changing from the sunny slope, the semi-sunny slope, the semi-cloudy slope to the cloudy slope, DBH and TH of the stand growth and DBH/TH withS.tsumuare decreased. TheS.tsumupure and mixed forest plantations take uniform distribution afforestation patterns. The overyielding tree species mixed withS.tsumuchange with the different altitude and aspect. That is,Cunninghamialanceolata(Lamb.) Hook.,Cinnamomumcamphora(L.) Presl andLiquidambarformosanaHance are below 500 m with the cloudy slope;Cunninghamialanceolata(Lamb.) Hook.,BetulaluminiferaH. Winkl.,LiquidambarformosanaHance andHoveniaacerbaLindl. are below the 500 m with the semi-cloudy slope;Castanopsiseyrei(Champ. ex Benth.) Tutch.,LiquidambarformosanaHance andSchimasuperbaGardn. et Champ. are between 500 and 1 000 m with the cloudy slope;SchimasuperbaGardn. et Champ.,QuercusacutissimaCarr.,ElaeocarpusdecipiensHemsl.,LiquidambarformosanaHance andStewartiasinensisRehd. Et Wils. are between 500 and 1 000 m with the semi-cloudy slope.
Northern subtropical zone;Sassafrastsumu(Hemsl.) Hemsl.; Altitude; Aspect; Spatial structure
孙洪刚,男,1976年8月生,中国林业科学研究院亚热带林业研究所,副研究员。E-mail:honggangsun@caf.ac.cn。
姜景民,中国林业科学研究院亚热带林业研究所,研究员。E-mail:jmjiang6001@126.com。
2016年3月6日。
S724;Q948.2
责任编辑:戴芳天。
1) 林业公益性行业科研专项(201404104);中央级公益科研院所基本科研业务费专项(RISF201405);优秀青年人才支持计划重点项目(gxyqZD2016306)。
猜你喜欢
南方农业(2022年23期)2022-02-14
思维与智慧·下半月(2021年7期)2021-08-13
现代农村科技(2018年11期)2018-11-15
现代农业研究(2017年11期)2018-01-12
河北地质(2017年2期)2017-08-16
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
课程教育研究(2017年11期)2017-04-17
现代园艺(2017年20期)2017-02-03
