
烤烟淀粉含量的配合力与遗传分析
2017-05-30 23:45张振臣马柱文李集勤袁清华谢锐鸿李淑玲陈俊标
热带作物学报 2017年2期
张振臣 马柱文 李集勤 袁清华 谢锐鸿 李淑玲 陈俊标


摘 要 为了解烤烟烤后中部烟叶淀粉含量的遗传规律,选用K326、NC89、翠碧一号和红花大金元等4个品种,按完全双列杂交设计,采用Griffing方法Ⅰ进行配合力分析,并应用植物数量性状“主基因+多基因”混合遗传模型对K326×红花大金元组合P1、P2、F1和F2等4个世代群体进行遗传分析。结果表明:烤烟烤后中部叶淀粉含量的遗传主要受细胞核效应的影响,一般配合力方差和特殊配合力方差达极显著水平,一般配合力方差大于特殊配合力方差,翠碧一号的一般配合力表现为较高的正向效应,K326的一般配合力表现为较高的负向效应;K326×红花大金元组合烤后中部叶淀粉含量的遗传符合2对加性-显性-上位性主基因+加性-显性多基因模型(E-1),主基因遗传率(hmg2)为49.53%。
关键词 烤烟;淀粉含量;配合力;遗传分析;品种
中图分类号 S572 文献标识码 A
Abstract In order to understand the genetic regularities of starch content in flue-cured tobacco leaves,four flue-cured tobacco varieties were used as the parents in a complete diallel mating design. Analysis of combining ability was conducted by Griffing -Ⅰ. Inheritance of starch content in flue-cured leaves in P1, P2, F1 and F2 from the cross K326× Honghuadajinyuan was investigated by the mixed major gene plus polygene inheritance model of quantitative traits. The results indicated that starch content in the flue-cured tobacco was controlled by nucleolus inheritance. The significant differences in starch content of GCA and SCA effects existed among parents, and the GCA effect was mainly. The GCA of Cuibiyihao showed positive effect, the GCA of K326 showed negative effect. Starch content in flue-cured leaves of the cross K326×Honghuadajinyuan was controlled by two adding-dominance-epistatic major gene plus adding-dominance polygene model. The heritability of major gene was 49.53%.
Key words flue-cured tobacco leaves; starch content; combining ability; genetic analysis; varieties
doi 10.3969/j.issn.1000-2561.2017.02.009
淀粉是烟叶的重要化学成分之一。淀粉在烤烟叶片细胞中的合成、积累、分解、转化状况,决定着烤后叶片内部各种化学成分之间的协调程度[1]。淀粉在烘烤过程中分解的单糖可使烟叶弹性好、吃味佳,但是烤后烟叶的淀粉含量较高会使烟叶燃烧不良,产生难闻的气味,不仅影响烟气质量,而且使安全性下降[2]。不同烤烟品种的淀粉含量差异较大,邓云龙等[3]研究发现烤烟烟叶接近成熟时淀粉含量最高,红花大金元中部叶片淀粉含量为40%左右,K326为25%左右。丁金玲等[4]研究发现,在烘烤过程中,K326和红花大金元的淀粉含量变化规律基本上相同,淀粉含量在烘烤84 h后降至最低点(红花大金元降至7.33%,K326降至6.4%)。张会芳[5]研究认为,不同基因型烤烟的烤后烟叶淀粉含量差异达极显著水平。目前有关烤烟淀粉含量的遗传机制研究较少。本研究通过对烤煙烤后中部叶淀粉含量进行配合力分析与“主基因+多基因”多世代遗传分析,初步了解其遗传规律,以期为优质烤烟新品种选育提供理论依据。
1 材料与方法
1.1 材料
K326、NC89、翠碧一号和红花大金元,4个亲本的完全双列杂交组合(F1)共12个,K326×红花大金元组合F2代。
1.2 方法
2013年11月将4个亲本和12个完全双列杂交F1代在广东南雄市古市播种,2014年2月移栽,试验材料随机区组设计,3次重复,每小区种植50株,行株距1.2 m×0.6 m。每666.7平方米施纯氮量9.7 kg(N ∶ P2O5 ∶ K2O=1 ∶ 0.5 ∶ 1.6,下同)。田间栽培管理与当地优质烤烟生产一致。选取所有无病害的正常烟株第9~12叶位烟叶挂牌,在小区取样烘烤(密集式烤房,三段式烘烤工艺),采用流动分析法(YC/T216-2007)检测烤后9~12叶位烟叶淀粉含量。
2014年2月将K326(P1)、红花大金元(P2)及其F1和F2分别移栽种植于南雄市古市。亲本、F1各设3个重复,随机区组设计,每小区种植30株,F2种植300株,试验区四周设保护区。施肥量与上述田块相同,田间栽培管理与当地优质烤烟生产一致。选取无病害的正常烟株(P1、P2、F1每小区10株,F2代161株)的第9~12叶位烟叶挂牌,对单株进行取样并烘烤(密集式烤房,三段式烘烤工艺),用流动分析法(YC/T216-2007)检测烤后单株9~12叶位烟叶淀粉含量。
1.3 数据统计与分析
配合力分析按照Griffing方法I[6],应用DPS软件进行分析。采用数量性状“主基因+多基因”混合遗传分析windows版本软件包SEA对P1、P2、F1和F2等4个世代单株的烤后烟叶淀粉含量进行分析[7-9],根据AIC值和适合性检验的结果确定最优模型,估算最适合模型的主基因和多基因效应值、遗传率等一阶遗传参数和二阶遗传参数。
2 结果与分析
2.1 烤烟亲本及F1组合的烤后中部叶淀粉含量
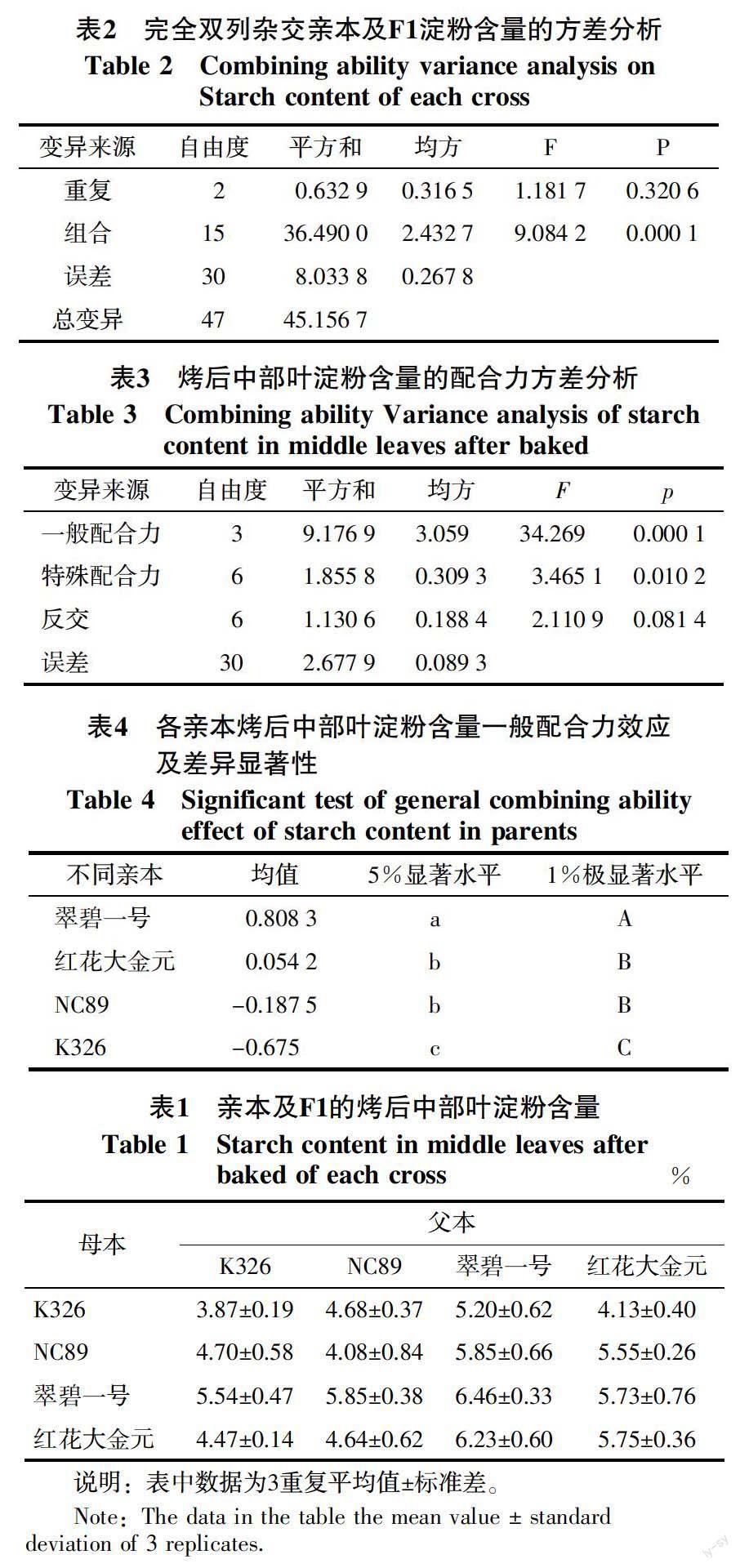
烤烟亲本及F1组合的烤后中部叶平均淀粉含量为3.87%~6.46%,标准差为0.19~0.84(表1)。4个亲本中,K326和NC89的淀粉含量较低,分别为3.87%和4.08%;翠碧一号和红花大金元的淀粉含量较高,分别为6.46%和5.75%。F1组合中,K326与NC89正反交F1的淀粉含量高于双亲,其他组合的淀粉含量位于双亲之间。方差分析(表2)表明,参试材料间的淀粉含量存在极显著差异,可以进一步进行配合力分析。
2.2 烤后中部烟叶淀粉含量的配合力分析
2.2.1 配合力方差分析 从表3中可以看出,一般配合力和特殊配合力方差达到极显著水平,一般配合力方差远大于特殊配合力方差,二者比值为9.9,表明参试品种烤后中部烟叶淀粉含量的遗传以加性效应为主,非加性效应处于次要地位。反交效应未达到显著水平,淀粉含量主要受细胞核效应的影响。
2.2.2 一般配合力效应分析 对各亲本的一般配合力效应值进行差异显著性分析(表4)。结果表明,NC89和红花大金元的效应值差异不显著,其他亲本的效应值差异均达极显著水平。4个亲本中,翠碧一号具有较高正值的一般配合力效应值, K326具有较高负值的一般配合力效应值,表明试验选用的亲本对于涉及淀粉含量品种的选育具有较高的育种价值,K326可作为选育低淀粉含量的烤烟亲本。
2.3 K326×红花大金元组合烤后中部叶淀粉含量的遗传分析
2.3.1 4个世代的淀粉含量表现 K326(P1)和红花大金元(P2)的平均淀粉含量分别为3.01和4.95,K326×红花大金元组合F1和F2的平均淀粉含量位于两亲本之间;K326淀粉含量的单株变异系数较小,红花大金元和F1淀粉含量的单株变异系数稍大。F2的淀粉含量表型分布呈现偏态分布,推测该组合的淀粉含量存在主效基因的作用(表5)。
2.3.2 淀粉含量的遗传模型及效应估计 对K326×红花大金元组合的亲本、F1和F2世代群体的烤后中部叶淀粉含量进行“主基因+多基因”混合遗传分析,各遗传模型的AIC值见表6,遗传模型E-1、E-0和D-0的AIC值相对较小。由适合性检验结果可知(表7),E-1、E-0和D-0均有1个显著偏离,因此选择AIC值最小的E-1为最佳遗传模型,即2对加性-显性-上位性主基因+加性-显性多基因模型。
根据遗传模型中成分的分布和相应的均值求得一阶遗传参数和二阶遗传参数(表8)。K326×红花大金元组合的2对主基因加性效应值分别为2.663 5和2.663 5,2对主基因加性效应均为正效应;二者的显性效应值分别为0.344 3和1.258 3,第2对主基因显性效应值是第一对主基因的3.65倍。2对主基因互作效应较明显,加性×加性效应(i)为-0.344 3,显性×显性效应值(l)为-1.258 8;第1对主基因加性效应与第2对主基因显性效应的互作效应(jab)为-1.749 4;第2对主基因加性效应与第1对主基因显性效应的互作效应(jba)为-2.663 5。加性效应值(da+db)、显性效应值(ha+hb)和上位性效应值(i+jab+jba+l)分别为5.327、1.602 6、-6.016。K326×红花大金元组合淀粉含量还存在多基因修饰,多基因加性效应值[d]和显性效应值[h]分别为-6.032 3和-0.559 1。该组合分离世代F2群体淀粉含量主基因遗传方差(σmg2)为1.205 8,主基因遗传率(hmg2)为49.53%,表明该组合的淀粉含量的主基因遗传率较小。
3 讨论
3.1 作物淀粉含量的遗传
关于淀粉含量的配合力与遗传机制,在水稻、玉米、麦类、高粱等作物上都有相关报道[10-18]。金正勋等[10]对水稻杂种后代稻米直链淀粉含量进行配合力分析,结果表明加性效应和非加性效应对稻米直链淀粉含量均起重要作用。国际上关于水稻直链淀粉含量的遗传模式有3种结论:1对基因和部分修饰基因;2对显性互补基因;数量性状的多基因[11]。普通玉米和超甜玉米籽粒淀粉含量主要受加性基因控制,同時也受非加性基因影响[12-14]。薛香等[15]认为小麦籽粒直链淀粉、支链淀粉和总淀粉含量的遗传受加性效应和非加性效应控制,且以加性效应为主。方先文等[16]研究认为小麦籽粒淀粉含量受2对主基因和多基因控制,支链淀粉含量受1对主基因和多基因控制。吕亮杰等[17]研究认为大麦直链淀粉含量符合2对连锁抑制作用主基因模型和两对连锁互补作用主基因+加性多基因遗传模型,支链淀粉含量符合2对抑制作用主基因+加性多基因遗传模型和2对连锁显性-上位性作用主基因+加性多基因遗传模型,支/直符合两对抑制作用主基因模型,总淀粉含量符合3对等比例加性作用主基因+加性多基因遗传模型。丁国祥等[18]研究认为,加性和非加性效应对糯高粱总淀粉含量、直链淀粉含量和支链淀粉含量的表型值均有重要作用。
在烤烟主要化学成分中,与淀粉密切相关的可溶性总糖和还原糖含量的遗传研究较多,大多数研究[19-23]认为,烤烟的总糖和还原糖含量受加性和非加性效应基因的共同控制,以加性效应为主。本研究通过遗传分析发现,烤烟烤后中部烟叶淀粉含量性状主要受细胞核效应的影响,加性效应和非加性效应对烤烟淀粉含量均起重要作用,以加性效应为主;不同亲本间一般配合力效应差异极显著,K326具有较高的一般配合力负向效应值,可以作为选育低淀粉含量的烤烟亲本;K326×红花大金元组合淀粉含量的遗传符合2对加性-显性-上位性主基因+加性-显性多基因模型(E-1)。张会芳[5]研究认为,烤烟淀粉含量的遗传以非加性效应为主,与本文结论不一致,可能与参试材料不同有关。
本研究选用了4个有代表性的烤烟生产品种进行淀粉含量的配合力分析,选用的亲本数量虽然偏少,但这些品种是目前国内烤烟新品种选育的主体亲本[24],而且在常规化学成分和致香物质等方面具有较大差异[25],本研究的遗传分析结果具有一定的育种参考价值。
3.2 环境对烤烟淀粉含量的影响
烤后烟叶淀粉含量除受控于基因型外,还受环境因子、栽培条件和烘烤措施等因素的影响[3,26-30]。本研究中,在K326×红花大金元组合世代群体中,红花大金元(P2)和K326×红花大金元 F1的单株淀粉含量间均存在较大差异,这种差异可能与品种适应性、大田肥力、栽培管理、采摘叶片成熟度、烤房中挂放的区域等差异有关,同时也表明,烤烟的淀粉含量受环境因素影响较大。
参考文献
[1] 官长荣, 王能如, 汪耀富. 烟叶烘烤原理[M]. 北京: 科学出版社, 1994: 46-49.
[2] 刘国顺. 烟草栽培学[M]. 北京: 中国农业出版社, 2003: 25.
[3] 邓云龙, 孔光辉, 武锦坤, 等. 氮素营养对烤烟叶片淀粉积累及SPS、 淀粉酶活性的影响[J]. 烟草科技, 2001(11): 34-37.
[4] 丁金玲, 杨焕文, 梁旭清, 等. K326和红花大金元在烘烤过程中烟叶淀粉含量和淀粉酶活性变化规律[J]. 云南农业大学学报, 2003, 18(4): 382-384.
[5] 张会芳. 烤烟叶片淀粉含量的遗传变异及农艺措施调控研究[D]. 郑州: 河南农业大学, 2005.
[6] 刘来福, 毛盛贤, 黄远樟. 作物数量遗传[M]. 北京: 农业出版社, 1984: 125-249.
[7] 章元明, 盖钧镒, 张孟臣. 利用P1F1P2和F2或F2∶3世代联合的数量性状分离分析[J]. 西南农业大学学报, 2000, 22(1): 6-9.
[8] 盖钧镒, 章元明, 王健康. 植物数量性状遗传体系[M]. 北京: 科学出版社, 2003: 169-219.
[9]曹锡文, 刘 兵, 章元明. 植物数量性状分离分析Windows软件包SEA的研制[J]. 南京农业大学学报, 2013, 36(6): 1-6.
[10] 金正勋, 崔成焕, 秋太权. 水稻杂种后代稻米直链淀粉含量的配合力分析[J]. 东北农业大学学报, 1999,30(2): 122-127.
[11] 张运锋, 谭学林. 水稻直链淀粉的影响因素、 直链淀粉对加工品质的影响及遗传[J]. 福建稻麦科技, 2004, 22(3): 9-11.
[12] 魏良明, 刘占先, 程泽强, 等. 普通玉米3个籽粒品质性状的配合力分析[J]. 中国农学通报, 2010, 26(15): 179-184.
[13]刘宗华, 王 琳, 汤继华, 等. 不同环境条件下玉米子粒淀粉含量的配合力效应及杂种优势分析[J]. 玉米科学, 2007, 15(1): 11-15.
[14] 刘鹏飞, 蒋 锋, 陈 趣,等. 超甜玉米淀粉含量的遗传特性与配合力分析[J]. 恺农业工程学院学报,2013, 26(2): 12-15.
[15] 薛 香, 粱云娟, 杨忠强, 等. 小麦淀粉品质性状的遗传及配合力分析[J]. 种子, 2009, 28(4): 18-20.
[16] 方先文, 姜 东, 戴廷波, 等. 小麦籽粒总淀粉及支链淀粉含量的遗传分析[J]. 作物学报, 2003, 29(6): 925-929.
[17] 吕亮杰, 郭元世, 杜丽杰, 等. 大麦籽粒淀粉含量的主基因+多基因遗传模型分析[J]. 麦类作物学报, 2014, 34(1): 13-22.
[18] 丁国祥, 赵甘霖, 张长伟, 等. 酿酒糯高梁亲本系籽粒淀粉含量及其组份的配合力分析[J]. 种子, 2011, 30(2): 10-11.
[19] 蔡长春, 邓 环, 程 玲, 等. 烤烟化学成分的遗传效应分析[J]. 烟草科技, 2015, 48(1): 1-6.
[20] 刘仁祥, 代 飞, 韩孝六, 等. 烤烟主要化学品质性状的遗传效应研究[J]. 江西农业学报, 2011, 23(4): 101-104.
[21] 肖炳光, 朱 军, 卢秀萍, 等. 烤烟几种主要化学成分的遗传分析[J]. 作物学报, 2005, 31(12): 1 557-1 561.
[22] 肖炳光, 卢秀萍, 焦芳蝉, 等. 烤烟几种化学成分的QTL初步分析[J]. 作物学报, 2008, 34(10): 1 762-1 769.
[23] 许 健, 杨 德, 张锦伟. 烤烟亲本配合力的双列杂交分析[J]. 烟草科技, 2004(1): 29-32.
[24] 常爱霞, 贾兴华, 冯全福, 等. 我国主要烤烟品种的亲源系谱分析及育种工作建议[J]. 中国烟草科学, 2013, 34(1): 1-6.
[25] 赵会纳, 卢秀萍, 许自成, 等. 不同基因型烤烟主要化学成分与香味物质的多变量分析[J]. 郑州轻工业学院学报(自然科学版), 2009, 24(1): 11-16.
[26] 宫长荣, 刘 霞, 郭 瑞, 等. 淀粉代谢及影响烤烟淀粉含量的因素[J]. 云南农业大学学报, 2006, 21(6): 742-745.
[27] 董维杰, 张忠锋, 窦玉青, 等. 烤烟烟叶淀粉含量影响因素及其与烟叶质量关系研究进展[J]. 广东农业科学, 2015(8): 11-16.
[28]王 劍, 李佳颖, 叶贤文, 等. 凉山烤烟淀粉含量状况及与感官质量的关系分析[J]. 山东农业科学,2013, 45(2): 45-49.
[29] 包可翔. 翠碧1号烟叶总糖、还原糖和淀粉含量的区域分布特征[J]. 安徽农业科学, 2011, 39(8): 4 486-4 488.
[30] 王建波, 黎摇娟, 周清明, 等. 湖南浓香型烤烟区淀粉含量变化及聚类分析[J]. 作物研究, 2013, 27(6): 558-560, 567.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
湖北农机化(2020年4期)2020-07-24
活力(2019年19期)2020-01-06
中国蔬菜(2019年5期)2019-06-04
猪业科学(2018年5期)2018-07-17
长江蔬菜(2017年5期)2017-05-10
河北农业科学(2016年2期)2016-09-09
长江蔬菜(2016年24期)2016-03-15
新疆农垦科技(2014年6期)2014-02-28
湖南农业科学(2014年18期)2014-02-27
