
运动员大脑皮层的神经效率:来自一般视觉识别任务ERP与ERD/S的证据
2017-05-29 02:37:45李安民
天津体育学院学报 2017年5期
魏 瑶 ,李安民
大量研究显示,练习不仅提升任务绩效,还能引起大脑皮层可塑性变化[1,2]。一些研究表明,高任务绩效者的任务相关皮层激活降低,表现出大脑皮层加工的高效率,提出了大脑皮层神经效率假说[3-5]。运动领域研究发现,长期高强度训练,不仅提高运动员任务绩效,也诱发运动员大脑皮层结构功能的可塑性改变。依据神经效率假说,运动员的高绩效可能伴随神经功能的增强,并表现任务皮层激活的降低[6]。
但是,横向研究来看,一些脑电图(EEG)或神经功能影像学(fMRI)的证据显示,与控制组相比,运动员的任务脑区激活变化趋势并不一致。如与控制组相比,高尔夫运动员的感觉运动区[7-8]、空手道选手的额-颞-枕腹侧通路[9]、射击和射箭运动员的额-颞-枕皮层[10-12]、职业舞蹈家的额-颞叶皮层[13-14]、乒乓球选手的额-颞-枕皮层[15-16]、职业赛车的视觉空间和运动皮层[17],在任务认知加工过程,呈现更低的激活水平;但是,在篮球预测投篮[18]、网球和排球发球识别[19-20]、芭蕾舞蹈动作观察[21-22]、乒乓球动作表象[23]、高尔夫推杆[24]、击剑动作控制[25]等任务加工过程,运动员皮层激活程度更高。一些事件相关电位(ERP)的横向研究也显示,与控制组相比,运动员诱发的ERP成分潜伏期或波幅的变化趋势也不一致。如与控制组相比,射击[26]、棒球[27]、散打[28]、乒乓球[29]运动员一般任务诱发的ERP成分(如P3)平均波幅更低;但是,在乒乓球专项图形识别[30]、羽毛球动作预测[31]、击剑决策[32]等任务中,与控制组相比,运动员诱发的ERP成分(如P3)平均波幅更高。ERP成分的平均波幅,被作为个体认知加工注意资源的消耗程度,与控制组相比,运动员诱发的ERP平均波幅更高或更低证据都存在,表明运动员的大脑皮层资源消耗程度的趋势并不一致。
并且,从纵向研究来看,被试练习前后的皮层激活变化趋势也不一致。如一些视觉认知[33]、记忆[34]、电游[4]等技能学习的研究显示,与练习初期相比,随着技能熟练化,被试的大脑额-顶皮层的激活程度逐渐降低,呈现皮层的神经效率。但是,另一些复杂动作学习[35-36]和多维感知整合训练[37-38]显示,随着认知绩效提高与自动化加工形成,被试的任务皮层激活变化趋势并不一致[39],在动作学习结束后初期,练习者的初级运动区(M1)激活逐渐降低,但当熟练掌握后5~10天左右,被试的M1区、辅助运动区(SMA)、动作皮层、皮下中枢激活增强[40-41]。
上述横向或纵向研究的不一致表明,运动员大脑皮层的神经效率,可能存在条件性限制。有研究者认为,练习绩效水平与大脑皮层激活程度的趋势不一致,可能受到任务特异性与复杂性、认知策略与经验、神经激活通路、皮层激活重组等因素的影响[1,5,42]。运动领域相关研究也表明,与对照组相比,运动员的大脑皮层神经激活的变化趋势可能受到运动类型、训练方向、大脑半球等因素的影响[43]。因此,通过练习绩效与大脑激活变化的相关性,来解释是否因长期练习导致的大脑皮层神经效率,需要考虑调节变量效应[44]。有研究者提出,加工策略与皮层环路是否一致,是探讨个体间任务皮层神经效率的必要条件,只有采用相同加工策略和皮层神经环路[2,45],才可以将皮层激活程度的差异性,解释为大脑皮层的神经效率。因此,验证长期训练是否对运动员大脑皮层的神经效率产生影响,一般应采用被试较熟悉的任务,确保运动员与控制组的任务加工策略和神经环路相同[45-46],才能判断运动员与控制组的任务脑区激活差异,是任务皮层的神经效率,而不是认知策略或皮层区域的变化所致。
此外,评价指标选择的差异性,也可能影响神经效率的评价。目前,研究有关皮层神经激活评价指标主要包括血氧含量(fMRI)和脑电指标,前者通过血氧含量间接反映脑功能激活,空间分辨率高;后者直接反映脑神经皮层激活特征,时间分辨率高。因此,在强调时间精确度的大尺度脑皮层功能活动分析中,脑电指标具有明显优势。其中,脑电包括EEG频域和ERP时域分析技术,两者的理论依据和分析方法都不同。其中,ERP基于诱发模型,反映皮层短暂神经认知电位,主要是锁时锁相慢节律脑波,波幅表示认知资源消耗程度。而EEG节律频谱分析基于神经震荡理论,通过分析诱发脑波节律振荡能量分布,探讨认知加工的神经机制。有研究认为[47],大脑皮层间(或皮层与中间神经元间)节律波同步震荡,是大脑皮层认知加工的生理机制,与静息态相比,任务过程皮层alpha节律值下降,表现为alpha节律去同步化;恢复过程皮层alpha节律值增强,表现为alpha节律波同步化,可以用alpha节律去同步化/同步化水平,反映个体任务加工皮层的激活程度。此外,EEG和ERP的优势各异,其中,ERP时间分辨率高,但不能提供频域能量分布特征;EEG频域分析可以提供大脑神经的脑波能量特征,但无法表现出神经能量分布的时间特征。EEG时-频分析(如小波分析)能够提供节律波时域和能量分布特征,但对低频节律波的解析不理想[48-49]。因而,脑电信号探讨认知过程的神经机制,如果将ERP和EEG节律分析技术结合,可能更全面地解释认知神经机制。
根据大脑可塑性理论[50],长期多年的专项训练,可能诱发运动员大脑皮层结构和功能的适应性变化。运动领域一般采用横向比较方法,通过运动员和控制组皮层激活差异性,揭示长期运动训练对大脑皮层的可塑性。由于运动项目的特异性,不同训练任务对大脑皮层环路的影响可能存在差异,因此,通过专项任务探讨运动员皮层功能高效性。但是,被试对专项任务熟悉性或加工方式可能存在差异,这不利于皮层功能激活差异性的解释。运动员的大脑激活差异性,可能是专项策略或经验导致皮层区域的变化,而不是相同任务皮层功能水平。因此,探讨运动员大脑皮层神经效率,需要控制任务熟悉性和加工皮层通路等变量,使2组被试的试验任务过程使用一样的加工方式,诱发相同的皮层神经环路。前期音乐[51]、运动领域[52]相关研究显示,采用简单或熟悉性任务,能诱发相同的皮层环路,这是皮层神经效率比较的前提。
乒乓球运动属于典型的快速对抗性开式技能项目,需要个体较强的视-动认知能力,承受较高视觉注意负荷。长期乒乓球训练,可能促进运动员运动绩效,并提高视觉空间与动作控制能力,进而优化视觉-动作皮层网络,提高皮层的神经效率。但是,现有研究显示,无论是一般性任务,还是专项任务,与控制组相比,乒乓球运动员皮层表现的激活(或注意资源)程度的差异性并不一致[15-16,23,30,53]。这也反映试验任务选择不同,诱发不同的大脑皮层神经网络,不同的诱发皮层通路干扰对运动员皮层神经效率的评价。为此,本研究随机选择乒乓球运动员,通过横向对比研究,采用一般性简单视-空图形认知任务,既能确保诱发专项相关的皮层通路(激活相同的皮层环路),又能避免专项经验和加工策略使用差异性(采用相同的策略加工)。基于脑电对神经元活动反映的直接性,以及时间分辨率高等优势,本研究采用脑电技术,探讨长期乒乓球训练如何影响运动员视觉-动作加工皮层功能,进而表现运动员大脑皮层的神经效率。并且,为了从综合评价加工时间过程和神经能量消耗特征,探讨运动员大脑任务皮层功能变化特征,本研究将ERP时域和EEG频域分析相结合,进行任务诱发脑电信号分析。根据本研究目的和前期研究结果,随机抽取8年以上训练年限的国家二级乒乓球运动员为试验组,普通专业大学生为控制组,试验任务为简单一般性的带缺口的特征圆任务,主要选择视觉背侧通路和动作皮层等兴趣区(如OZ、PZ、CZ、FZ电极点),比较乒乓球运动员与控制组脑电指标的差异性,揭示运动员大脑神经效率的生理机制。并提出以下假设:(1)运动员兴趣区电极点诱发的ERP成分平均波幅低于控制组,表现为认知注意资源的节省化;(2)运动员的兴趣区电极点诱发Alpha节律ERD/S水平低于控制组,呈现视觉-动作环路皮层的神经效率;(3)任务诱发ERP成分与Alpha节律时-频能量分布是一致的。
1 研究对象与方法
1.1 被 试
被试由随机抽取的大学生运动员和普通大学生组成,其中,试验组为大学乒乓球二级运动员19名,平均年龄(19.68±2.062)岁,平均训练年限10年以上;对照控制组为普通专业大学生20名,平均年龄(19.75±1.746)岁。全部被试身体健康,无精神器质性疾病,瑞文推理测验显示智力水平中等以上,2组被试学历、年龄、智力水平等组别差异不显著(P>0.05)。通过中国人利手评定调查结果显示,全部被试均为右利手。招募的被试都自愿参与试验,试验前签订自愿协议,然后熟悉试验流程与任务,积极配合完成试验全过程,试验结束后支付一定的报酬。
1.2 试验任务与步骤
运动员一般性任务通常采用简单线、圆、点等简单图形[28,32],有关乒乓球运动员的研究显示,在一般认知加工任务(如特征圆)中,运动员与控制组的行为绩效或脑电存在差异[29,54-55]。为了保证被试加工策略和诱发皮层神经通路的一致性,避免认知加工策略和诱发环路的不同,对皮层激活与绩效相关性结果解释的干扰效应,采用一般性的带缺口特征圆为试验材料,其中,缺口10°,依据缺口中心与水平夹角的不同角度,分为8种不同空间朝向的特征,角度分别为0°、45°、90°、135°、180°、225°、270°和315°。其中,缺口朝右下45°的特征圆为靶刺激,全部类型的带缺口的圆,通过E-Prime2.0编程随机呈现,流程见图1。其中,被试只对缺口朝右下45°的特征圆(靶刺激)进行按键辨别反应,其余类型的特征圆(非靶刺激)不做按键反应。靶刺激和非靶刺激随机呈现,其中,靶刺激为60次,非靶刺激60次,共120次。试验开始前,被试要进行试验前准备工作,如填写被试基本信息、洗头、带电极帽、打脑电膏降电阻、调节视距位置和头部U型托架固定等。试验分练习和正式试验2部分,要求被试双眼与屏幕中心呈水平,距离保持60 cm,尽量不晃动身体,控制眨眼,以降低脑电伪迹。被试先进行练习任务,熟悉任务要求后,进入正式试验阶段。

图1 任务刺激呈现的时间进程Figure1 Time Process of the Stimulus Presentation
1.3 试验仪器与数据采集
采用2台计算机,一台呈现E-Prime 2.0编程的刺激任务和记录行为数据,一台记录德国BP脑电64导测试系统同步EEG信号,并记录垂直眼电(ECG)和水平眼电(EOG)。要求被试的头皮电极电阻<10 KΩ,脑电采样率为500 Hz,滤波带通为0.5~30 Hz。2台试验所用的显示器均为19英寸液晶显示器,分辨率为1 024×768,刷新频率为100 Hz。
1.4 数据处理
1.4.1 行为数据 采用E-prime2.0软件包中E-Run运行程序来采集行为数据,记录被试的反应时和反应正确率。通过E-Data Aid初步筛选和处理数据,并采用E-Merge合并不同组被试的行为数据。依据3个标准差法则,筛选所有行为数据,剔除加减3个标准差之外的数据。行为数据的统计分析过程:先采用K-S法进行行为数据分布的正态性检验,然后采用t检验,比较2组被试的平均正确率、反应时的差异显著性(α=0.05)。
1.4.2 脑电ERP数据 将BP Recorder2.0软件采集的脑电数据,采用BP网络版Analyzer2.1软件对脑电数据进行离线预处理。(1)通过双侧乳突转换参考电极,并采用独立成分分析法去除眼电(Ocular Correction),半自动分析去除伪迹;(2)将预处理数据以刺激锁时方式分段EEG数据,每个片段时长1 000 ms(-200~800 ms),并进行基线校正,再次去伪迹,自动剔除波幅大于±100 μv的伪迹波;(3)对每个被试的靶刺激诱发的有效EEG片段进行叠加,得到每个被试任务加工的ERP波形;(4)通过波峰检测,确定被试的ERP各成分(如P1、N1、P2、N2、P3等)的潜伏期与波幅;(5)分别对2组被试的波形进行叠加,获得2组被试的总平均(Average)波形图;(6)依据以往研究结果和本研究目的,依据“前中后”和“左中右”选取规则,选取FZ、F3、F4,CZ、C3、C4,PZ、P3、P4,OZ、O1、O2等电极点,重点考察枕区(OZ)、顶区(PZ)、中央区(CZ)和额区(FZ)等主要电极点诱发的ERP成分。本研究采用2(运动员组、控制组)×4(枕、顶、中央、额)重复测量方差分析,检验不同兴趣区(枕、顶、中央、额)和组别(运动员组、控制组)各成分的平均波幅和潜伏期组别差异显著性(α=0.05)。
1.4.3 脑电数据的时-频分析 采用小波时-频分析技术,能同时呈现脑电信号的时间和频率特征,反映大脑不同时间阶段加工的节律波能量变化特征[56]。小波分析(Wavelet Analysis)是采用某一函数族,将原始信号与小波权函数进行卷积,将原始信号分解成不同频率的块信号,提供信号的时间-频率能量分布的方法。因此,小波分析被广泛使用EEG信号分析中。本研究首先采用BP自带Analyzer2.1处理软件,对EEG数据的去眼电、伪迹和分段等预处理,然后采用复Morlet小波基函数,对单个trail的EEG片段进行小波变换,转换公式:Wχ(τ,f)=,参数选择为,频率范围为1~100 HZ,频率进步为1,f/σf设定为7,输出值选择Spectral Power(μV2),归一化输出(Normed Output)时间为 0~600 ms,基线矫正(Based on Time)时间为-200~0 ms。然后,将每个被试正确反应trail的EEG时-频小波能量分布进行叠加平均。最后,将全部被试有效数据总叠加平均,获得主要兴趣区电极点EEG信号的时-频能量分布图。
前期研究显示,任务期大脑皮层alpha节律震荡功率值降低,而任务结束后皮层alpha节律震荡功率值增强[47],研究者常采用alpha节律去同步化/同步化(ERD/ERS)评价大脑皮层激活程度。为了比较运动员和对照控制组大脑激活差异,进一步提取(-200~600ms)事件相关alpha(8~13HZ)节律的功率值,以-200~0 ms平均功率为基线值(R),0~600 ms的平均功率为事件期(A),运用公式ERD/ERS=[(A-R)/R]×100%,计算2组被试兴趣区电极点alpha节律(8~13 Hz)的去同步化/同步化水平,通过比较2组alpha节律波去同步化/同步化水平,评价皮层激活组间差异性。其中,ERS为正值,显示alpha节律同步化,皮层激活降低(钝化现象);ERD为负值,显示alpha节律去同步化,皮层激活增强。本研究兴趣区为枕叶(OZ)、顶叶(PZ)、中央区(CZ)和额叶(FZ)等背侧视觉-动觉皮层通路,采用2(运动员组、控制组)×4(OZ、PZ、CZ、FZ)两因素重复测量方差分析方法,以alpha节律的ERD/ERS水平为因变量,检验2组被试这些脑区的激活差异显著性(α=0.05)。
2 结果
2.1 行为数据
将2组被试反应时(RT)和正确率(ACC)等行为数据,分别进行独立样本t检验和χ2检验,比较组间差异显著性(α=0.05)。检验结果显示,运动员组[(363.8±38.1)ms]和对照组[(379.4±36.3)ms]的反应时(RT)差异不显著(t=1.31,P>0.05),运动员组(98.6%±2.4)和对照组(97.1%±3.7)的正确率(ACC)差异也不显著(χ2=0.115,P>0.05),表明本研究的运动员与控制组任务绩效相同。
2.2 脑电ERP数据
从事件相关电位(ERP)的总平均波形图(见图2)可知,2组被试的枕区(Oz)、顶区(Pz)、中央区(Cz)和额区(Fz)等脑区诱发明显的ERP成分,表明任务主要诱发视觉空间认知的大脑皮层背侧通路。根据本研究目的和假设,主要分析兴趣区诱发明显的N1、N2和P3等成分潜伏期与波幅。
(1)对4个脑区的N1成分进行2(组别:运动员、控制组)×4(脑区:枕、顶、中央、额区)重复测量方差分析。N1成分潜伏期统计结果显示,脑区主效应显著[F(3,111)=167.332,P<0.001,η2p=0.819],基于估算边际均值(α=0.05)的成对比较显示,全部被试不同脑区的平均潜伏期大小依次为额区(115.811 ms)=中央区(112.608 ms)<顶区(162.179 ms)=枕区(169.272 ms),组别主效应不显著[F(1,37)=0.132,P>0.05,η2p=0.007],脑区和组别交互效应显著[F(3,111)=2.605,P>0.05,η2p=0.071],表明 4个脑区N1潜伏期组间差异不显著。N1成分平均波幅统计结果显示,脑区主效应显著[F(3,111)=15.856,P<0.01,η2p=0.301],组别主效应不显著[F(1,37)=1.374,P>0.05,η2p=0.0363],组别与脑区交互效应显著[F(3,111)=19.740,P<0.01,η2p=0.346]。进一步简单效应分析显示,枕区[F(1,37)=13.536,P<0.01,η2p=0.268]、额区[F(1,37)=12.471,P<0.01,η2p=0.251]和中央区[F(1,37)=9.843,P<0.01,η2p=0.210]组别效应显著,其中,运动员枕区的平均波幅小于控制组(-1.322 μVVS-2.601 μV,P<0.01),运动员额区(-2.803 μV VS-1.291 μV,P<0.01)和中央区(-2.367 μV VS-1.768 μV,P<0.01)的平均波幅大于控制组,顶区的组别效应不显著[F(1,37)=0.095,P>0.05,η2p=0.003](见图3)。
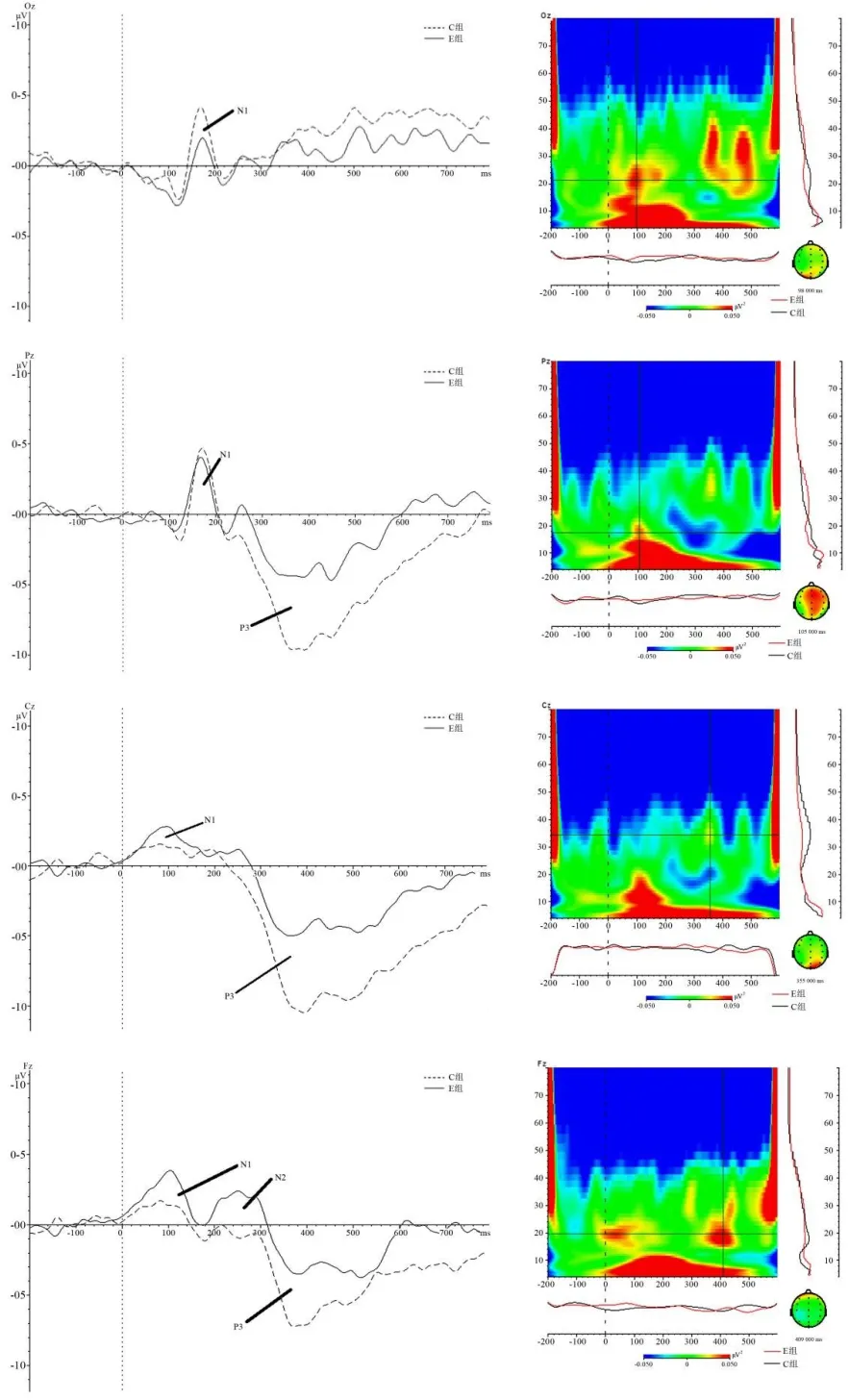
图2 主要电极点ERP和EEG时-频能量分布图Figure2 ERP and EEG Time-Frequency Energy Distribution Map of the Main Electrode Sites
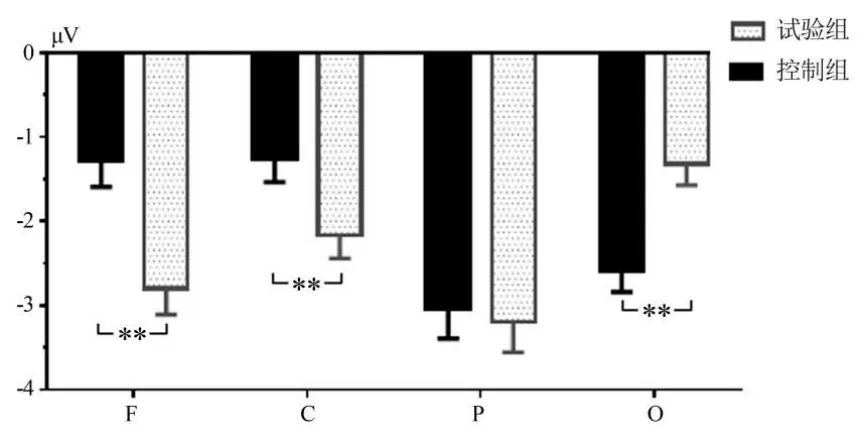
图3 主要脑区N1成分的平均波幅Figure3 Average Peak of N1 of the Main Area Cortex
(2)对4个脑区N2成分的平均潜伏期和波幅,进行2×4重复测量方差分析。潜伏期的统计结果显示,脑区主效应不显著[F(3,111)=1.380,P>0.05,η2p=0.036],组别主效应不显著[F(3,111)=0.057,P>0.05,η2p=0.002],脑区和组别交互效应不显著[F(3,111)=0.920,P>0.05,η2p=0.024],表明N2成分潜伏期的脑区和组别差异都不显著。N2 波平均波幅的统计结果显示,脑区[F(3,111)=0.208,P>0.05,η2p=0.006]和组别[F(1,37)=3.342,P>0.05,η2p=0.083]主效应不显著,脑区和组别交互效应显著[F(3,111)=3.461,P<0.05,η2p=0.086]。简单效应分析表明,额区[F(1,37)=8.516,P<0.01,η2p=0.187]、顶区[F(1,37)=6.005,P<0.05,η2p=0.140]的组别差异显著,中央区[F(1,37)=2.217,P>0.05,η2p=0.057]、枕区[F(1,37)=0.590,P>0.05,η2p=0.016]的组别差异不显著。事后比较显示,运动员的额区(-1.815 μV VS0.553 μV,P<0.01)、顶区(-0.663 μV VS2.296 μV,P<0.01)平均波幅大于控制组(见图4)。
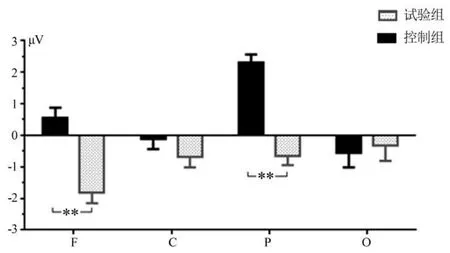
图4 主要脑区N2成分的平均波幅Figure4 Average Peak of N2 of the Main Area Cortex
(3)对不同脑区P3成分的平均潜伏期和波幅,进行2×4重复测量方差分析。P3波的平均潜伏期统计结果显示,脑区主效应显著[F(1,37)=8.516,P<0.01,η2p=0.187],组别主效应不显著[F(1,37)=1.670,P>0.05,η2p=0.043],脑区和组别交互效应不显著[F(1,37)=2.173,P>0.05,η2p=0.055]。脑区间潜伏期均值的多重比较显示,额区=中央区=顶区>枕叶,并且,4个脑区平均潜伏期的组间差异不显著(P>0.05)。P3波的平均波幅统计结果显示,脑区主效应显著[F(3,111)=153.301,P<0.01,η2p=0.806],组别主效应显著[F(1,37)=17.132,P>0.05,η2p=0.007],脑区和组别交互效应显著[F(3,111)=7.733,P<0.01,η2p=0.173]。简单效应分析显示,额区[F(1,37)=11.688,P<0.01,η2p=0.240]、中央区[F(1,37)=19.271,P<0.01,η2p=0.342]和顶区[F(1,37)=29.118,P<0.01,η2p=0.440]的组别效应显著,枕区[F(1,37)=2.493,P>0.05,η2p=0.063]组别效应不显著。事后检验可知,运动员额区(3.13μV VS 6.42μV,P<0.01)、中央区(5.86μV VS 9.56μV,P<0.01)和顶区(3.26μV VS 7.89μV,P<0.01)平均波幅均小于控制组(见图5)。
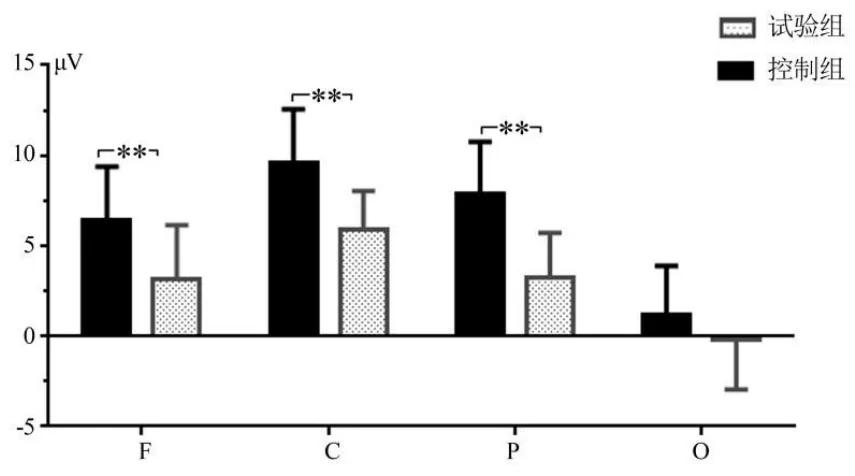
图5 主要脑区P3成分的平均波幅Figure 5 Average Peak of P3 of the Main Area Cortex
2.3 脑电EEG时-频能量分布和ERD/S数据
根据本研究目的和假设,重点关注任务诱发的ERP波形与EEG的alpha(8~13 HZ)节律能量变化趋势,因此,主要比较兴趣区电极点(Oz、Pz、Cz、Fz)ERP和EEG时频能量分布图。首先,重点关注基线期(-200~0 ms)、早期(0~200 ms)和晚期(200~600 ms)的alpha节律(8~13 HZ)的能量变化,分析任务认知过程,被试的主要兴趣区(Oz、Pz、Cz、Fz)早期和晚期的皮层激活的变化趋势(见图2)。将刺激呈现早期(0~20 0ms)和晚期(200~600 ms)的alpha节律功率图谱,分别与基线期(-200~0 ms)alpha节律波图谱比较。结果显示:早期(0~200 ms)alpha节律的功率值增强,呈现同步化(alpha-ERS),反映大脑皮层抑制加工[57];晚期(200~600 ms)alpha节律功率低于基线,呈现alpha节律去同步化(alpha-ERD),反映大脑皮层执行加工。并且,这种图谱显示的去同步化现象,在顶区(Pz)和中央区(Cz)更明显。
为了比较2组被试任务脑区皮层的激活差异性,计算2组被试兴趣区(Fz、Cz、Pz、Oz)的alpha节律ERD/ERS水平,以组别为被试间变量,脑区为被试内变量,alpha节律的ERD/ERS水平为因变量,进行2(运动员组,控制组)×4(额区、中央区、顶区、枕区)重复测量方差分析。结果显示,脑区主效应显著[F(3,111)=4.978,P<0.01,η2p=0.119],组别主效应显著[F(1,37)=12.025,P<0.01,η2p=0.245],脑区与组别的交互效应不显著[F(3,111)=1.178,P>0.05,η2p=0.031]。不同脑区的组别差异性单变量检验显示,中央区[F(1,37)=5.942,P<0.05,η2p=0.138]和顶区[F(1,37)=6.110,P<0.05,η2p=0.142]的组别效应显著,额区[F(1,37)=2.463,P>0.05,η2p=0.081]和枕区[F(1,37)=1.699,P>0.05,η2p=0.044]的组别效应不显著。事后比较显示,运动员组的中央区、顶区的alpha去同步化水平低于控制组(P<0.05)(见图6)。
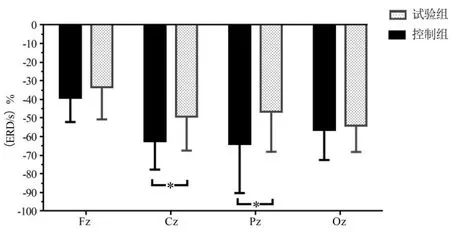
图6 主要脑区alpha ERD/S的平均值Figure6 Alpha ERD/S Average of the Main Area Cortex
3 讨论
本研究采用简单的一般性刺激辨认任务,目的保证被试采用相同的认知策略和神经环路,最大限度减少被试的认知策略和工作记忆容量限制,对神经活动与绩效关系解释的干扰效应,揭示被试皮层神经效率的差异性[46]。因此,尽管本研究结果显示行为绩效组别差异不显著,但是,并不影响研究评价个体的神经皮层效率的差异性。只要保证2组被试认知策略和神经通路一致性,在相同绩效前提下,神经皮层的激活程度不同,也反映个体任务脑区的神经效率差异性[17]。个体间神经效率评价,包括更高绩效被试的皮层资源更低消耗,或者是相同绩效条件下个体资源消耗的差异性(激活水平)。行为绩效相同的被试,表现出神经激活的差异性,也能反映皮层资源消耗的高效率。如2个耐力水平不同的跑步者,在相同时间和负荷条件下,完成2 000 m跑过程中,高水平耐力选手的消耗能量相对少、感觉更轻松,而低水平选手消耗更多能量、感觉更累一样,高耐力选手的体能储备更多,能够适应更强的运动负荷。
3.1 ERP成分的差异性分析
前期研究认为,ERP诱发成分的潜伏期与信息加工速度相关,而波幅则反应个体注意资源加工投入的程度[58]。既然ERP波幅反映了被试注意资源的消耗程度,那么,通过比较运动员和对照组的皮层诱发ERP波幅,可以探讨长期训练对运动员皮层中枢神经资源消耗的影响。由于采用相对简单的任务,工作记忆负荷需求低,加工过程不受工作记忆容量的限制,因此,诱发ERP成分潜伏期组别差异不显著,表明2组被试对该任务的加工速度相同。如果2组被试诱发的ERP波幅差异显著,说明中枢注意资源消耗是不同的,相同绩效投入的中枢神经资源少,同样是大脑皮层神经效率的体现。对兴趣区电极点诱发ERP波幅组别比较显示,ERP早成分(如N1)和晚成分(如P3)都存在组间差异(P<0.05),且受脑区位置和加工进程影响。
(1)运动员枕区ERP成分N1平均波幅低于控制组,表明运动员视觉辨别加工消耗的神经资源呈现节约化趋势,这可能与顶-枕皮层神经元激活数量或程度更低有关。有趣的是,运动员的额区和中央区的N1波幅高于控制组,与假设不一致。枕区的视觉诱发N1成分,潜伏期160 ms左右,与刺激辨别加工相关,运动员更低枕叶N1波幅,表明运动员在视觉任务加工早期,消耗的枕区资源更低。额区、中央区诱发的N1成分,与早期反应准备加工相关[59]。运动员额-中央区诱发的N1波幅更高,表明行为反应早期运动员消耗注意资源更多,这可能与运动员长期从事专项训练相关,在乒乓球等快速反应类项目运动情境中,运动者更多通过早期注意加工,才能快速应答,既快又准击球,满足运动情境动作快速响应需要,可能为晚期的大脑执行加工节约资源,促进大脑皮层神经效率。根据注意容量模型[60],个体中枢注意资源是有限的,通过分配方案完成认知加工。研究认为,早期注意属于无意识、内隐的、无限制性,晚期注意属于意识性、外显的、容量有限性[61]。长期运动训练可能提高运动员自上而下加工通路连接性,有利于额区早期无意识资源对动作加工的影响,提高快速任务的应答绩效,并且,节约有限的晚期注意资源,满足运动任务加工注意的高需求,反映了运动员大脑神经资源利用的高效率。乒乓球[62]、网球[63]等研究也显示,运动员的额区诱发的N1波幅也高于对照组,与本研究结果一致。
(2)本研究结果显示,运动员额区诱发N2成分的平均波幅高于对照组,表明运动员保持较高的认知控制加工水平,其原因可能与运动员长期从事对抗性项目相关,比赛过程需要保持高度动作认知控制,保证快速准确的动作应答反应。在简单的视觉空间特征加工任务中,也保持较高认知控制状态,调整自己动作反应。额-顶诱发的N2成分,反应个体的行为监控、动作控制、抑制加工,运动员较强的抑制控制能力是运动员对动作快速应答与激烈对抗的适应,运动员通过动作抑制皮层网络加工,促进后期动作认知顺利完成,与运动员运动认知任务完成过程的抑制控制相适应,表明长期训练促进运动员皮层的动作控制能力[64]。前期乒乓球[62]、网球[63]等研究也显示,运动员额叶诱发更大的N2波,反映了运动员大脑更强的抑制控制能力,与本研究结果一致。
(3)运动员的中央区(CZ)和顶区(PZ)诱发P3成分平均波幅低于对照组,表明运动员背侧视觉神经通路相关皮层在完成相同任务绩效过程中,消耗的神经加工资源更少,体现任务相关皮层的神经效率。研究表明,P300波幅随着个体对刺激任务注意程度的变化而改变[65],诱发脑源可能位于颞顶联合区[66]或海马回[67],与物体辨别和工作记忆刷新相关,反映注意、辨别、决策应答等认知加工过程[68]。本研究结果显示,运动员诱发P3成分波幅低于控制组,表明运动员大脑中央区和顶区神经资源消耗更低,验证了神经效率假说。注意双加工模型认为,控制加工向自动化加工转化,中枢注意资源需求逐渐降低[69]。运动员视觉动作加工的自动化,节约更多的晚期意识性注意资源,适应快速准确加工任务。因此,长期训练促进运动认知和动作加工自动化,通过早期无意识激活,减少后期有意注意资源的消耗,将运动员有限的意识注意资源,用于其他运动信息领域加工,提高运动任务绩效。长期训练对运动员皮层的神经注意资源消耗的转变,提高了运动员视觉-动作皮层的加工效率。
然而,一些专项识别任务研究显示,运动员额-顶区诱发的P3等晚成分波幅高于对照组,与中枢皮层资源节省化的高效率不一致,这可能是不同被试组,对任务熟悉程度不同有关,运动员和对照组可能采用的加工策略或皮层通路不同,或者被试主观难度认知水平太高,影响个体努力程度和任务自信,这样,运动员复杂任务消耗神经资源反而高于对照组。
3.2 Alpha节律ERD/S的差异性分析
本研究显示,运动员的顶区和中央区的alpha节律ERD水平小于控制组,表明运动员的皮层神经能量的消耗更低,体现运动员的视觉空间与动作控制皮层的神经效率,这与本研究假设一致。前期相关研究显示,被试皮层的alpha节律波ERD/ERS水平与fMRI血氧信号变化密切相关,皮层激活增强与alpha-ERD相关,皮层活动抑制与alpha-ERS相关[70-73]。
根据视觉注意加工的背腹侧双通路理论[61],个体对视觉空间特征加工主要涉及顶枕额背侧通路。本研究简单任务加工期间,运动员的顶叶和中央皮层较低的激活,显示运动员消耗视觉空间和动作加工的神经资源更少。表明,长期训练提高运动员视觉空间和动作控制能力,呈现顶区和中央皮层的神经效率,这些结果与大脑可塑性理论和神经效率假说具有一致性。乒乓球运动需要个体快速视觉识别和合理动作执行能力,长期专项训练提高运动者的视觉空间认知和动作加工能力,促进皮层结构与功能适应性变化,表现运动员大脑任务皮层的神经效率。有关研究认为,高绩效者大脑皮层激活降低,可能是激活的皮层神经环路或者神经元数量减少相关[3]。根据神经网络(节点)连接理论,高绩效者的任务脑区激活降低,可能是皮层功能区(节点)之间连接有效性提高,或者连接数量下降相关[46]。心理模块理论认为[74],大脑神经系统是一种高度模块化表征方式,模块内外具有不同节点连接特性,功能模块内的节点连接比较紧密,而功能模块之间的节点比较稀疏。如果功能模块之间(如功能脑区皮层间)节点有效性较低,需要激活更多的节点连接,才能完成认知任务,皮层激活更强。此外,突触修剪理论认为[75],大脑发育过程,个体皮层神经元连接(突触)数量随着年龄增加而下降,但突触连接强度有提高,运动训练可能诱发皮层-皮层或皮层-皮下效连接加强,皮层神经连接通过“削枝强干”,减少低效连接,提高皮层(节点)有效连接,降低神经元激活数量,减少神经元资源消耗,任务相关皮层激活下降,表现皮层神经效率[11]。本研究运动员顶叶、中央区等皮层激活降低,可能是长期专项训练强化功能连接有效性减少了低效率的视觉通路等皮层的连接数量,神经元激活数量下降,节约更多视觉中枢资源,将视觉注意资源用于运动更高的注意需求。在简单认知和动作任务条件下,射击、射箭、足球、赛车选手相关皮层激活降低[9,11,25,76],与本研究结果一致。
但是,一些复杂性视觉空间、视觉搜索和动作观察任务的研究[19-23,30]显示,运动员的顶区、中央区等任务皮层激活更强,与神经效率假设不一致。其可能原因:(1)对照组专项经验和策略缺乏,任务完成自信心与努力投入较少,相关神经皮层资源投入更少,诱发任务相关脑区激活更低;(2)可能与默认网络(Default Network)有关,运动员控制加工下降,默认网络显示激活水平更高[77];(3)长期专项训练可能导致皮层激活环路重组,诱发皮层环路不同所致[1]。
本研究还显示,任务诱发枕区激活水平组间差异不显著,与预期假设不同。可能是因为本研究是一般性简单视觉任务,视觉皮层对简单任务的敏感性低于专项任务,因而,诱发个体皮层程度低,并未体现训练效应。运动员在复杂性或专项视觉任务加工过程中,枕叶激活程度低于普通组,这可能因为复杂任务需要动员更多的皮层连接,提高视觉皮层对外部刺激的响应加工速度。同时,额叶皮层激活(alpha-ERD)组间差异不显著,与假设不一致,其原因可能是运动员早期注意资源(如N1、N2波幅更大)消耗能量大,晚期注意资源消耗低,但是从总体来看,运动员的额叶神经激活(alpha-ERD)平均值不显著,说明简单任务条件下,2组被试对额叶执行控制加工需求基本一致。
3.3 ERP成分与EEG频谱变化的相关性分析
EEG时-频能量分布图,直观反映EEG不同节律波能量分布的时间变化趋势,本研究重点关注背侧通路和动作皮层的电极点。数据显示,无论早期时间窗(0~200 ms),还是晚期时间窗(200~600 ms),alpha节律(8~13 Hz)同步化(ERS)与ERP成分负电位(如N1、N2成分)变化一致,alpha节律去同步化(ERD)与ERP的正电位(如P3)相关。依据大脑皮层节律振荡研究[47],皮层神经网络通过EEG节律活动,实现神经信息加工,其中,低频节律波(如alpha波)的同步化反映皮层神经元抑制,高频节律波(如beta、gamma)同步化反映皮层激活加工,反映感知(视觉)皮层神经元与其他皮层(或中间神经元)的神经信息交流。由于高频锁时不锁相,叠加获得的ERP成分,高频beta、gamma等节律波被抵消,并且,ERP只是直接叠加,并不是节律能量功率值,而只是短时电位平均振幅。但对比EEG时频能量分布和ERP成分原始波,还是不难发现两者变化趋势的一致性。
前期相关研究显示,皮层ERP成分P3潜伏期和波幅变化,与alpha节律变化高度关联[71]。听觉Oddball任务研究显示,P3波幅与alpha活动的抑制相关[57]。有关听觉、视觉、躯体感觉和痛觉刺激Oddball任务研究发现,即使P300和alpha-ERD的皮层诱发源不同,但P300与alpha-ERD水平高度相关[70]。但是,另一些研究却显示,刺激诱发alpha节律波震荡活动与ERP波幅变化趋势并不完全一致,既有呈反比关系的,也有呈正比关系[57,71-72]。其原因可能与两者的锁时时间窗差异性相关,或者是alpha节律ERD/ERS,不但反映皮层-皮层,还反映皮层-皮层下的神经网络信息流。因此,本研究将ERP高时间分辨率优势和EEG振荡能量特征相结合,能够更好地解释运动员大脑皮层的神经效率,揭示运动员视觉任务认知加工的神经机制。此外,被试枕叶皮层高频节律波(如gamma)同步化现象表明,任务诱发被试枕叶皮层。已有研究证实,高频节律波震荡与认知加工密切相关[73]。后期考虑进一步分析gamma节律和alpha节律的振荡特征,将可能更好地揭示运动员大脑神经效率机制。同时,脑区间神经元集群的同步化,反映大脑神经网络信息加工特征,因此,后期研究需要考虑脑区激活与脑区间功能连接程度,从全脑网路视角,揭示运动员大脑的神经加工特征,揭示运动员神经效率的生理机制。此外,EEG所面临的空间分辨率不足,也制约振荡信号诱发源的精确定位,需要考虑将EEG和fMRI技术相结合,进一步探讨运动员大脑神经效率。
4 结论
本研究以乒乓球运动员为研究对象,采用一般性视觉认知任务,将ERP和EEG频域分析相结合,探讨运动员大脑的神经效率。ERP结果显示,运动员枕叶的早期神经资源,以及额、顶、中央区晚期神经资源节约化明显,但额-顶皮层神经资源早期消耗更多,表明长期训练提高运动员大脑神经资源消耗效率,促进运动员快速加工的适应性。EEG频域分析显示,运动员的顶区和中央区的激活程度更低,呈现大脑皮层的神经效率。EEG时频能量分布与ERP波幅变化趋势的一致性表明,可以分别从节律能量和时间进程视角,更全面揭示大脑认知加工的神经电生理机制。
[1]KELLY A M,GARAVAN H.Human functional neuroimaging of brain changes associated with practice[J].Cereb Cortex,2005,15(8):1089-1102.
[2]POLDRACK R A.Imaging brain plasticity:conceptual and methodologi⁃cal issues-a theoretical review[J].Neuroimage,2000,12(1):1-13.
[3]HAIER R J,SIEGEL JR B V,NUECHTERLEIN K H,et al.Cortical glu⁃cose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography[J].Intelligence,1988,12(2):199-217.
[4]HAIER R J,SIEGEL JR B V,MACLACHLAN A,et al.Regional glu⁃cose metabolic changes after learning a complex visuospatial/motor task:a positron emission tomographic study[J].Brain Res,1992,570(1-2):134-143.
[5]NEUBAUER A C,FINK A.Intelligence and neural efficiency[J].Neuro⁃sci Biobehav Rev,2009,33(7):1004-1023.
[6]徐立彬,李安民,刘玉.优秀运动员大脑神经效率及可塑性特点[J].首都体育学院学报,2013,25(6):566-572.
[7]BABILONI C,DEL PERCIO C,IACOBONI M,et al.Golf putt outcomes are predicted by sensorimotor cerebral EEG rhythms[J].J Physiol,2008,586(1):131-139.
[8]BAUMEISTER J,REINECKE K,LIESEN H,et al.Cortical activity of skilled performance in a complex sports related motor task[J].Eur J Appl Physiol,2008,104(4):625-631.
[9]DEL PERCIO C,ROSSINI P M,MARZANO N,et al.Is there a“neural efficiency”in athletes?A high-resolution EEG study[J].Neuroimage,2008,42(4):1544-1553.
[10]BELL M A,FOX N A.Crawling experience is related to changes in cor⁃tical organization during infancy:evidence from EEG coherence[J].De⁃velopmental Psychobiology,1996,29(7):551-561.
[11]HATFIELD B D,HAUFLER A J,HUNG T M,et al.Electroencephalo⁃graphic studies of skilled psychomotor performance[J].Journal Clincal Neurophysiology,2004,21(3):144-156.
[12]DEL P C,BABILONI C,BERTOLLO M,et al.Visuo-attentional and sensorimotor alpha rhythms are related to visuo-motor performance in athletes[J].Hum Brain Mapp,2009,30(11):3527-3540.
[13]BABILONI C,DEL PERCIO C,ROSSINI P M,et al.Judgment of ac⁃tions in experts:a high-resolution EEG study in elite athletes[J].Neuro⁃image,2009,45(2):512-521.
[14]ORGS G,DOMBROWSKI J H,HEIL M,et al.Expertise in dance modu⁃lates alpha/beta event-related desynchronization during action observa⁃tion[J].Eur J Neurosci,2008,27(12):3380-3384.
[15]郭志平,李安民,王积福.乒乓球运动员的大脑功能非对称性及功能耦合性研究[J].天津体育学院学报,2015,30(3):256-261.
[16]王丽岩,李安民,顾楠,等.乒乓球运动员发球动作识别时的神经效率:来自EEG的证据[J].中国运动医学杂志,2013,32(5):430-436.
[17]BERNARDI G,RICCIARDI E,SANI L,et al.How skill expertise shapes the brain functional architecture:an fMRI study of visuo-spa⁃tial and motor processing in professional racing-car and naive drivers[J].PLoS One,2013,8(10):e77764.
[18]AGLIOTI S M,CESARI P,ROMANI M,et al.Action anticipation and motor resonance in elite basketball players[J].Nat Neurosci,2008,11(9):1109-1116.
[19]BALSER N,LOREY B,PILGRAMM S,et al.The influence of expertise on brain activation of the action observation network during anticipa⁃tion of tennis and volleyball serves[J].Front Hum Neurosci,2014,8:568.
[20]BALSER N,LOREY B,PILGRAMM S,et al.Prediction of human ac⁃tions:expertise and task-related effects on neural activation of the ac⁃tion observation network[J].Hum Brain Mapp,2014,35(8):4016-4034.
[21]CALVO-MERINO B,GLASER D E,GREZES J,et al.Action observa⁃tion and acquired motor skills:an FMRI study with expert dancers[J].Cereb Cortex,2005,15(8):1243-1249.
[22]CALVO-MERINO B,GREZES J,GLASER D E,et al.Seeing or doing?Influence of visual and motor familiarity in action observation[J].Cur⁃rent Biology,2006,16(19):1905-1910.
[23]WOLF S,BROELZ E,SCHOLZ D,et al.Winning the game:brain pro⁃cesses in expert,young elite and amateur table tennis players[J].Front Behav Neurosci,2014,8:e370.
[24]MILTON J,SOLODKIN A,HLUŠT K P,et al.The mind of expert motor performance is cool and focused[J].Neuroimage,2007,35(2):804-813.
[25]DI RUSSO F,TADDEI F,APNILE T,et al.Neural correlates of fast stimulus discrimination and response selection in top-level fencers[J].Neuroscience Letters,2006,408(2):113-118.
[26]ROSSI B,ZANI A,TADDEI F.Chronometric aspects of information pro⁃cessing in high level fencers as compared to non-athletes:an ERPs and RT study[J].Journal Human Movement Studies,1992,23(1):17-28.
[27]RADLO S J,JANELLE C M,BARBA D A,et al.Perceptual Decision Making for Baseball Pitch Recognition:Using P300 Latency and Am⁃plitude to Index Attentional Processing[J].Research Quarterly for Exer⁃cise and Sport,2001,72(1):22-31.
[28]赵洪朋.优秀散打运动员知觉预测过程认知特征与神经机制研究[D].上海:上海体育学院,2010:60-75.
[29]赵思,李安民,张大为.乒乓球运动经验对视觉注意、认知控制和识别过程影响的ERP研究[J].天津体育学院学报,2014,29(1):33-37.
[30]徐立彬,李安民,陈爱国.乒乓球运动员单一特征信息模式识别的神经机制研究[J].西安体育学院学报,2014,31(4):488-499.
[31]WRIGHT D J,HOLMES P,DI RUSSO F,et al.Reduced Motor Cortex Activity during Movement Preparation following a Period of Motor Skill Practice[J].PLoS One,2012,7(12):e51886.
[32]黄琳.击剑运动员线索化决策特点及ERP研究[D].上海:上海体育学院,2014:14-70.
[33]BUEICHEKU E,MIRO-PADILLA A,PALOMAR-GARCIA M A,et al.Reduced posterior parietal cortex activation after training on a visual search task[J].Neuroimage,2016,135(4):204-213.
[34]GRABNER R H,NEUBAUER A C,STERN E.Superior performance and neural efficiency:the impact of intelligence and expertise[J].Brain Res Bull,2006,69(4):422-439.
[35]KARNI A,MEYER G.Functional MRI evidence for adult motor cortex plasticity during motor skill learning[J].Nature,1995,377(6545):155.
[36]KARNI A,MEYER G,REY-HIPOLITO C,et al.The acquisition of skilled motor performance:fast and slow experience-driven changes in primary motor cortex[J].Proc Natl Acad Sci U S A,1998,95(3):861-868.
[37]SEIDLER R D,PURUSHOTHAM A,KIM S G,et al.Neural correlates of encoding and expression in implicit sequence learning[J].Exp Brain Res,2005,165(1):114-124.
[38]WALDSCHMIDT J G,ASHBY F G.Cortical and striatal contributions to automaticity in information-integration categorization[J].Neuroim⁃age,2011,56(3):1791-1802.
[39]DAYAN E,COHEN L G.Neuroplasticity subserving motor skill learn⁃ing[J].Neuron,2011,72(3):443-454.
[40]FLOYER-LEA A,MATTHEWS P M.Changing brain networks for vi⁃suomotor control with increased movement automaticity[J].Journal of Neurophysiology,2004,92(4):2405-2412.
[41]XIONG J,MA L,WANG B,et al.Long-term motor training induced changes in regional cerebral blood flow in both task and resting states[J].Neuroimage,2009,45(1):75-82.
[42]MILTON J,SOLODKIN A,HLUSTIK P,et al.The mind of expert motor performance is cool and focused[J].Neuroimage,2007,35(2):804-813.
[43]VECCHIO F,DEL PERCIO C,BABILONI C.Is there a neural efficien⁃cy in the athlete's brain?[J].International Journal of Psychophysiology,2012,85(3):296-297.
[44]RYPMA B,BERGER J S,PRABHAKARAN V,et al.Neural correlates of cognitive efficiency[J].NeuroImage,2006,33(3):969-979.
[45]POLDRACK R A.Is“efficiency”a useful concept in cognitive neurosci⁃ence[J].Developmental Cognitive Neuroscience,2015,11(6):12-17.
[46]RYPMA B,PRABHAKARAN V.When less is more and when more is more:The mediating roles of capacity and speed in brain-behavior effi⁃ciency[J].Intelligence,2009,37(2):207-222.
[47]PFURTSCHELLER G,LOPES D S F H.Event-related EEG/MEG syn⁃chronization and desynchronization:basic principles[J].Clin Neuro⁃physiol,1999,110(11):1842-1857.
[48]ROSSO O A,MARTIN M T,PLASTINO A.Brain electrical activity analysis using wavelet-based informational tools(II):Tsallis non-ex⁃tensivity and complexity measures[J].Physica A,2003,497-511.
[49]QUIAN QUIROGA R,SAKOWITZ O W,BASAR E,et al.Wavelet Transform in the analysis of the frequency composition of evoked po⁃tentials[J].Brain Research Protocols,2001,8(1):16-24.
[50]POLDRACK R A.Imaging brain plasticity:conceptual and methodologi⁃cal issues--a theoretical review[J].Neuroimage,2000,12(1):1-13.
[51]HYDE K L,LERCH J,NORTON A,et al.Musical training shapes struc⁃tural brain developmen[J].The Journal of Neuroscience,2009,29(10):3019-3025.
[52]BABILONI C,PERCIO C D,MARZANO N,et al.Neurophysiologic mechanism of neural efficiency in humans:Can it explain performanc⁃es of athletes and patients with neurodegenerative diseases?[J].Clinical Neurophysiology,2015,126(3):32.
[53]郭志平.乒乓球运动员在经验相关图形识别中的神经效率一项EEG研究[J].首都体育学院学报,2017,29(1):78-84.
[54]郭志平,李安民,陆青,等.乒乓球运动员在经验相关图形识别中的神经效率:一项EEG研究[J].首都体育学院学报,2017,29(1):78-84.
[55]郭志平,李安民,金晨曦,等.乒乓球运动员在经验相关图形识别中的脑电相干性分析[J].天津体育学院学报,2016,31(3):233-239.
[56]赵晶晶,牟书,舒华,等.基于Morlet小波变换的EEG时频分析[J].中国医学物理学杂志,2006,23(1):56-59.
[57]YORDANOVA J,KOLEV V.Event-related alpha oscillations are func⁃tionally associated with P300 during information processing[J].Neurore⁃port,1998,9(14):3159-3164.
[58]POLICH J.Updating P300:an integrative theory of P3a and P3b[J].Clin Neurophysiol,2007,118(10):2128-2148.
[59]VOGEL E K,LUCK S J.The visual N1 component as an index of a dis⁃crimination process[J].Psychophysiology,2000,37(2):190-203.
[60]张淑华.认知科学基础[M].北京:科学出版社,2007:340.
[61]沈政,林庶芝.生理心理学[M].北京:开明出版社,2012:250.
[62]赵思,李安民,张大为.乒乓球运动经验对视觉注意、认知控制和识别过程影响的ERP研究[J].天津体育学院学报,2014,29(1):33-37.
[63]陆颖之,周成林.网球运动员反应抑制能力及其最佳时区的ERP特点[J].中国运动医学杂志,2013,32(11):974-979.
[64]PATEL S H,AZZAM P N.Characterization of N200 and P300:selected studies of the Event-Related Potential[J].International Journal Of Medi⁃cal Sciences,2005,2(4):147-154.
[65]KEY A P F,DOVE G O,MAGUIRE M J.Linking brainwaves to the brain:an ERP primer[J].Developmental Neuropsychology,2005,27(2):183-215.
[66]HEGERL U,FRODL-BAUCH T.Dipole source analysis of P300 com⁃ponent of the auditory evoked potential:a methodological advance?[J].Psychiatry Research:Neuroimaging,1997,74(2):109-118.
[67]TARKKA I M,STOKID S,BASILE L F H,et al.Electric source lo⁃calization of the auditory P300 agrees with magnetic source localization[J].Electroencephalography and Clinical Neurophysiology/Evoked Po⁃tentials Section,1995,96(6):538-545.
[68]LAMME V A F.Why visual attention and awareness are different[J].Trends in Cognitive Sciences,2003,7(1):12.
[69]SCHNEIDER R M S W.Controlled and Automatic Human Information Processing:II.Perceptual Learning,Automatic Attending,and a Gener⁃al Theory[J].Psychological Review,1977,84(2):123-190.
[70]PENG W,HU Y,MAO Y,et al.Widespread cortical α-ERD accompa⁃nying visual oddball target stimuli is frequency but non-modality spe⁃cific[J].Behav Brain Res,2015,295(4):71-77.
[71]PENG W,HU L,ZHANG Z,et al.Causality in the Association between P300 and Alpha Event-Related Desynchronization[J].PLoS One,2012,7(4):341-363.
[72]YORDANOVA J.P300 and alpha event-related desynchronization(ERD)[J].Psychophysiology,2001,38(1):143-152.
[73]BAŞAR E,BAŞAR-EROGLU C,KARAKAŞ S,et al.Gamma,alpha,delta,and theta oscillations govern cognitive processes[J].International Journal of Psychophysiology,2001,39(2):241-248.
[74]ZAMORA-L PEZ G,ZHOU C,KURTHS J.Cortical hubs form a mod⁃ule for multisensory integration on top of the hierarchy of cortical net⁃works[J].Frontiers In Neuroinformatics,2010,19(4):1-11.
[75]PAOLICELLI R C,BOLASCO G,PAGANI F,et al.Synaptic Pruning by Microglia Is Necessary for Normal Brain Development[J].Science,2011,333(6048):1456-1458.
[76]BERNARDI G,RICCIARDI E,SANI L,et al.How Skill Expertise Shapes the Brain Functional Architecture:An fMRI Study of Visuo-Spatial and Motor Processing in Professional Racing-Car and Naïve Drivers[J].PLoS One,2013,8(10):e77761.
[77]JUTTA S,MAYER A R,KONRAD MAURER,et al.Specialization in the default mode:Task-induced brain deactivations dissociate between visual working memory and attention[J].Hum Brain Mapp,2010,31(1):126-139.
猜你喜欢
癫痫与神经电生理学杂志(2022年6期)2022-02-10 13:55:46
青少年科技博览(中学版)(2019年7期)2019-10-11 04:13:55
文教资料(2019年31期)2019-01-14 02:32:05
价值工程(2018年25期)2018-09-26 10:14:36
校园英语·中旬(2016年8期)2016-07-09 21:25:53
西南交通大学学报(2016年4期)2016-06-15 20:29:36
华北电力大学学报(自然科学版)(2016年3期)2016-04-25 05:06:56
广西林业科学(2016年3期)2016-03-16 05:43:23
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国实用医药(2015年24期)2015-05-08 09:42:17
