
日本和牛Y染色体USP9Y基因多态性与父系起源研究
2017-05-24 14:42:57马志杰黄永震党瑞华陈生梅刘善斋蓝贤勇雷初朝
中国牛业科学 2017年1期
马志杰,黄永震,党瑞华,陈生梅,刘善斋,曹 晖,林 清,蓝贤勇,张 琪,陈 宏,雷初朝*
(1.西北农林科技大学动物科技学院,陕西 杨凌712100;2.青海大学畜牧兽医科学院,青海 西宁810016;3.国家肉牛牦牛产业技术体系亳州综合试验站,安徽 亳州 236800;4.陕西秦宝牧业股份有限公司,陕西 宝鸡 722300)
科学试验
日本和牛Y染色体USP9Y基因多态性与父系起源研究
马志杰1,2,黄永震1,党瑞华1,陈生梅2,刘善斋3,曹 晖4,林 清1,蓝贤勇1,张 琪1,陈 宏1,雷初朝1*
(1.西北农林科技大学动物科技学院,陕西 杨凌712100;2.青海大学畜牧兽医科学院,青海 西宁810016;3.国家肉牛牦牛产业技术体系亳州综合试验站,安徽 亳州 236800;4.陕西秦宝牧业股份有限公司,陕西 宝鸡 722300)
【目的】为了分析日本和牛Y染色体USP9Y基因遗传多态性与父系起源。【方法】利用PCR扩增与限制性酶酶切方法。【结果】对8头日本和牛纯种及19头和秦F1代杂种牛进行Y染色体USP9Y基因多态性及父系起源研究,结果表明:日本和牛与其杂种牛都具有USP9Y基因多态性,USP9Y基因的PCR产物均显示471 bp和552 bp两种带型,其中552 bp带型不能被SspI酶酶切,则这2种带型对应Y1和Y2两种普通牛单倍型组,表明日本和牛含有Y1和Y2两个父系支系,有2个普通牛父系起源。日本和牛和杂种牛单倍型多样度分别为0.5714±0.0945和0.1988±0.1121,提示日本和牛具有较高的父系遗传多样性。【结论】日本和牛具有较高的父系遗传多样性及2个普通牛父系起源。
日本和牛;Y染色体;USP9Y;多态性;父系起源
日本和牛是日本十分珍贵的优质肉牛品种资源,是当今世界公认的品质最优秀的良种肉牛之一,其大理石花纹明显、细嫩多汁、肌肉脂肪中饱和脂肪酸含量很低,风味独特,肉用价值极高,在日本被视为“国宝”[1]。 近年来,随着胚胎移植和体细胞克隆技术的发展,和牛胚胎移植、体细胞克隆及自体繁育已在我国获得成功[2,3]。同时,和牛冻精杂交改良我国本地黄牛品种等育种实践也取得一定进展[4,5]。在分子水平上,基于mtDNA D-loop区序列变异,王建英等[6]研究表明,日本和牛和渤海黑牛在mtDNA D-loop区具有丰富的遗传多样性,二者亲缘关系较近,日本和牛属普通牛起源,欧洲原牛、朝鲜牛和我国北方的部分牛种对日本和牛的形成可能起到了丰富种群的作用。
动物Y染色体遵循父系遗传,单倍型完整,突变率低,不易受重组和回复突变的影响,是研究动物父系遗传多样性与起源的理想工具[7]。近年来,Y染色体SNPs标记(Y-SNPs)已广泛用于世界家牛的父系起源进化研究,发现家牛有3种Y染色体单倍型组,即Y1、Y2和Y3[8-16]。值得关注的是,自Bonfiglio等[15]发现通过USP9Y基因内含子26上的一个81 bp片段插入,利用一个天然的限制性酶切位点进行琼脂糖电泳,能够直接对这3种Y染色体单倍型组进行分型,不用测序,只需PCR和酶切电泳分析就可以鉴别不同的Y染色体单倍型组,大大降低了分型成本和工作量。上述这一省时省力、成本低廉的检测技术得到较为广泛的应用[16]。然而,目前尚未见日本和牛及其杂种牛的父系遗传多样性及遗传背景的相关研究报道。鉴于此,本研究利用USP9Y基因PCR扩增和酶切电泳技术对日本和牛及其和秦F1代杂种进行Y染色体单倍型组分析及父系起源研究,以期弄清日本和牛及其杂种牛的父系遗传背景,为我国黄牛改良奠定理论基础。
1 材料和方法
1.1 样品采集及基因组DNA提取
在国家肉牛牦牛产业技术体系亳州综合试验站和陕西秦宝牧业公司分别采集8头纯种日本和牛和19头和秦F1代杂种牛(以日本和牛为父本,秦川牛为母本)血样或耳组织带回实验室,常规酚-氯仿法提取基因组DNA,TE稀释其浓度至10~20 ng/μL保存备用。
1.2 引物合成和PCR扩增
参照Bonfiglio等[15]报道的普通牛USP9Y 基因序列合成1对引物,其中上游引物(PF):5′-AAACCCTTCAAGGTAATAAAACAAAA-3′,下游引物(PR):5′- CACAGCTCCTCAAAACCAGA -3′,由生工(上海)有限公司合成。PCR体系(25 μL):2×PrimeSTAR Max Premix (TakaRa 大连公司) 10.5 μL,上、下游引物 (10 pmol·L-1) 各0.5 μL,模板DNA (10~20 ng/μL) 1μL,超纯水12.5 μL。
PCR反应程序:98℃变性10 sec,57℃退火15 sec,72℃延伸10 sec,35个循环,冷却至4℃保存。PCR产物经1.5% 琼脂糖凝胶(含GoldView核酸染料)电泳后,凝胶成像系统分析检测。
1.3 PCR产物的酶切分析
对PCR产物先进行凝胶电泳检测,以确保扩增的目的片段特异性较高而不影响酶切结果分析。后对PCR产物直接用SspI限制性内切酶进行酶切,用2%琼脂糖凝胶电泳检测。并根据其带型进行酶切分析,判定单倍型组。其中SspI酶切体系(20 μL)为:SspI酶(12 U/μL)1 μL,10×SspI Buffer 2 μL,PCR产物10 μL,超纯水7 μL。37℃酶切8~10 h,后对酶切结果进行电泳检测。
2 结果与分析
2.1 牛USP9Y 基因扩增多态性与SspI 酶切鉴定
通过对和牛及其杂种牛USP9Y 基因部分序列进行PCR扩增和电泳,结果显示,8头纯种日本和牛和19头杂种牛PCR产物均获得预期大小,有471 bp和552 bp两种产物(图1)。8头和牛中,有4头牛的PCR产物为471 bp,而另外4头牛的PCR产物为552 bp。然而,在19头杂种牛中,只有2头牛的PCR产物为471 bp,而另外17头牛的PCR产物均为552 bp。依据Bonfiglio等[15]的判定标准,确定4头和牛与2头杂种牛(471 bp)为普通牛Y1单倍型组,剩余4头和牛与17头杂种牛(552 bp)尚不能确定是普通牛Y2单倍型组还是瘤牛Y3单倍型组,有待SspI酶酶切才能鉴定。对不能确定是普通牛Y2单倍型组还是瘤牛Y3单倍型组的4头和牛与17头杂种牛(552 bp)的PCR产物进行SspI酶酶切分析,结果表明,这21头牛的PCR产物都不能被SspI酶切开(图1),与其PCR产物带型、大小完全相同。依据Bonfiglio等[15]的判定标准,则确定这21头牛均为普通牛Y2单倍型组。
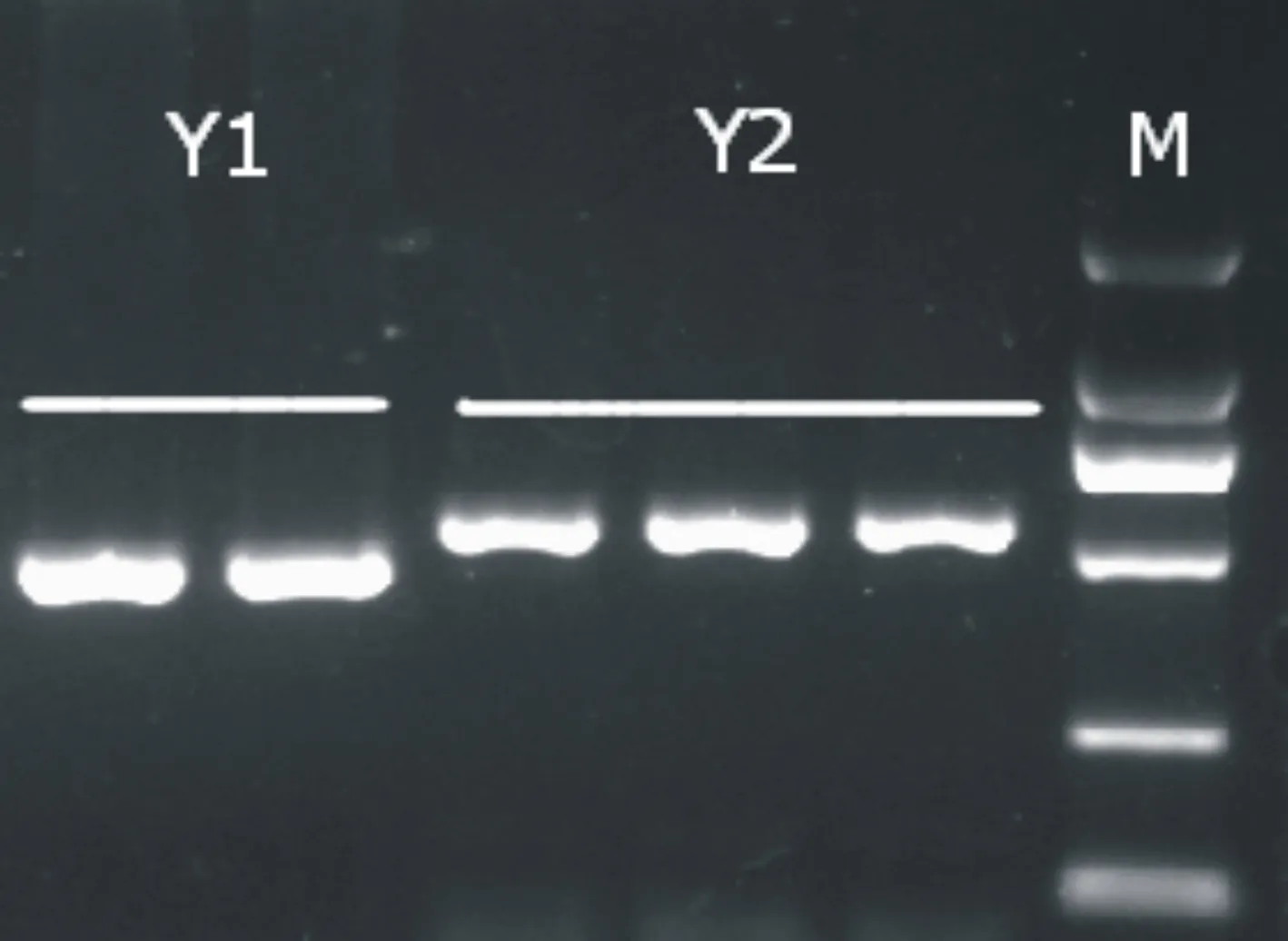
图1 日本和牛及和秦杂种牛USP9Y基因PCR产物扩增多态性
(M泳道为D 2000 marker,自上而下依次为2000、1000、750、500、250、100bp)
2.2 牛不同Y染色体单倍型组频率
对8头日本和牛和19头杂交牛中各自所占Y1和Y2单倍型组个体进行频率统计,结果显示:8头日本和牛中所含Y1和Y2个体分别为4个个体,频率均为0.50,单倍型多样度为0.5714±0.0945;而19头杂交牛中,含Y1和Y2单倍型组个体分别为2头和17头,所占频率分别为0.12和0.88,单倍型多样度为0.1988±0.1121(表1)。相比而言,日本和牛和杂种牛都含有两个Y染色体单倍型组,但日本和牛的Y染色体单倍型多样度相对较高。
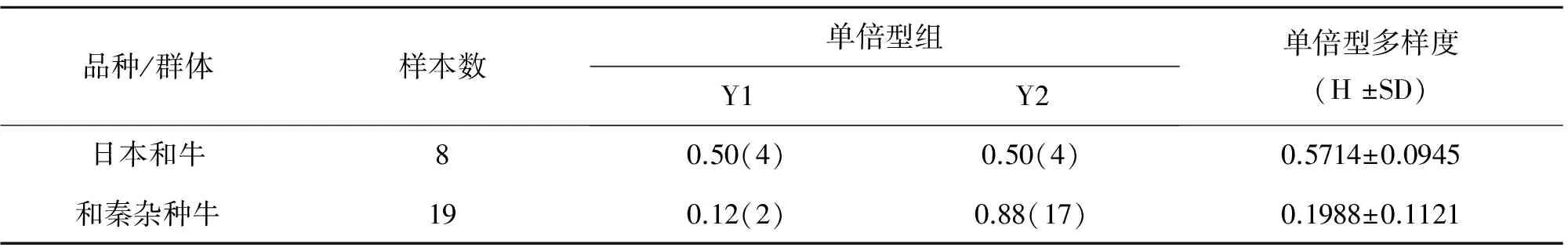
表1 日本和牛及和秦杂种牛Y1与Y2单倍型组的频率与多样度
3 讨论
近年来,围绕Y染色体分子标记(Y-SNPs和Y-STRs),国内外研究者对家牛进行了广泛深入的父系遗传研究,研究一致表明:Y染色体分子标记是追溯家牛起源、驯化历史和迁徙路线的重要工具,可在一定程度上揭示家牛的父系遗传多样性及群体间父系介导的杂交情况,家牛有3种父系起源(即普通牛Y1和Y2单倍型组以及瘤牛Y3单倍型组),Y-SNPs可以区分这3种单倍型组,而Y-STRs标记可将Y1、Y2和Y3所具有的丰富的单倍型进行精细区分[9-16]。然而,对日本和牛的父系遗传多样性及遗传背景的研究报道相对较少。
在本研究中,我们使用Bonfiglio等[15]发展的检测技术对8头纯种日本和牛和19头和秦F1代杂种进行了单倍型组分析,结果表明该方法快速有效,省时省力。4头日本和牛为普通牛Y1单倍型组,其余4头确定为普通牛Y2单倍型组,频率均为0.50,表明日本和牛有两个父系支系,有两个父系起源,拥有普通牛血统;而19头F1代杂种中,确定2头为普通牛Y1单倍型组,其余17头确定为普通牛Y2单倍型组,频率分别为0.12和0.88,表明杂种牛也有两个父系支系,有两个父系起源,拥有普通牛血统而无瘤牛血统。
在前期基于mtDNA D-loop区遗传变异分析中,研究表明日本和牛具有丰富的母系遗传多样性,属普通牛类型[6]。本文的结果表明日本和牛的父系起源与母系起源是一致的,表明日本和牛是普通牛起源。在本研究中,8头日本和牛和19头杂种牛单倍型多样度分别为0.5714±0.0945和0.1988±0.1121,表明日本和牛具有相对较高的父系遗传多样性。这表明从母系和父系遗传角度来看,日本和牛具有相对较高的分子遗传多样性,拥有丰富的遗传变异信息,遗传基础丰富,是改良中国地方黄牛,提高其大理石花纹的最佳终端父本。
[1] Hirooka H. Marbled Japanese Black cattle [J]. Journal of Animal Breeding and Genetics, 2014, 131(1):1-2.
[2] 王立平. 中国首例日本和牛在秦皇岛克隆成功[J]. 农业知识, 2009,60(21):9.
[3] 丁进东. 日本和牛胚胎移植的试验研究[J]. 中国牛业科学, 2009,35(4):28-30.
[4] 马平. 引进日本和牛冻精改良本地黄牛效果观察[J]. 中国牛业科学, 2014, 40(6):18-19.
[5] 李儒仁, 余群力, 韩玲, 等. 日本和牛与秦川牛杂交牛肝脏营养特性分析[J]. 营养学报, 2013, 35(5):502-504.
[6] 王建英, 张秀美, 兰邹然,等. 渤海黑牛和日本和牛mtDNA D-loop区遗传变异研究[J]. 中国农业大学学报, 2009, 14(3):75-82.
[7] Cortez D, Marin R, Toledo-Flores D, et al. Origins and functional evolution of Y chromosomes across mammals[J]. Nature, 2014, 508(7497), 488-493.
[8] Edwards C., Gaillard C., Bradley D, et al. Y-specific microsatellite polymorphisms in a range of bovid species[J]. Animal Genetics, 2000, 31:127-130.
[9] Gtherstrm A., Anderung C., Hellborg L, et al. Cattle domestication in the Near East was followed by hybridization with aurochs bulls in Europe[J]. Proceedings of the Royal Society B:Biological Sciences, 2005, 272(1579):2345-2350.
[10] Li M H, Zerabruk M, Vangen O, et al. Reduced genetic structure of north Ethiopian cattle revealed by Y-chromosome analysis[J]. Heredity, 2007, 98(4):214-221.
[11] Ginja C, Telo da Gama L. Penedo M C T. Y chromosome haplotype analysis in Portuguese cattle breeds using SNPs and STRs[J]. Journal of Heredity, 2009, 100(2):148-157.
[12] Kantanen J, Edwards C J, Bradley D G, et al. Maternal and paternal genealogy of Eurasian taurine cattle (Bos taurus) [J]. Heredity, 2009, 103(5):404-415.
[13] Pérez-Pardal L, Royo L J, Beja-Pereira A, et al. Y-specific microsatellites reveal an African subfamily in taurine (Bos taurus) cattle[J]. Animal Genetics, 2010, 41(3):232-241.
[14] Cortés O, Tupac-Yupanqui I, García-Atance M A, et al. Paternal genetic variability into the Lidia bovine breed [J]. Archivos De Zootecnia, 2011, 60(231):417-420.
[15] Bonfiglio S, De G A, Tesfaye K, et al. A novel USP9Y polymorphism allowing a rapid and unambiguous classification of Bos taurus Y chromosomes into haplogroups[J]. Animal Genetics, 2012, 43(5):611-613.
[16] Li R, Xie W M, Chang Z H, et al. Y chromosome diversity and paternal origin of Chinese cattle[J]. Molecular Biology Reports, 2013, 40(12):6633-6636.
Paternal Origins and Polymorphism of Y ChromosomeUSP9YGene in Japanese Black Cattle
MA Zhi-jie1,2, HUANG Yong-zhen1, DANG Rui-hua1, CHEN Sheng-mei2, LIU Shan-zhai3, CAO Hui4, LIN Qing1, LAN Xian-yong1, ZHANG Qi, CHEN Hong1, LEI Chu-zhao1
( 1.CollegeofAnimalScienceandTechnology,NorthwestA&FUniversity,Yangling,Shaanxi712100; 2.AcademyofAnimalScienceandVeterinaryMedicine,QinghaiUniversity,Xining810016; 3.BozhouComprehensiveExperimentalStation,NationalBeefcattleandYakIndustrialTechnologySystem,Bozhou,Anhui, 236800; 4.ShaanxiQinbaoAnimalhusbandryLimitedcompany,Shaanxi,Baoji, 722300)
[Objective] To explore the genetic polymorphism of Y chromosomeUSP9Ygene and paternal origins in Japanese black cattle. [Method] Using PCR and restriction enzyme digestion method. [Result] The polymorphism of Y chromosomeUSP9Ygene and paternal origins of eight Japanese black cattle and 19 F1 hybrids from the hybridization between male Japanese black cattle and female Qinchuan cattle were analyzed. The results showed that Japanese black cattle and the hybrids indicatedUSP9Ygene polymorphism with PCR products of 471 bp and 552 bp respectively, and the 552 bp PCR products can't be digested bySspIenzyme. Therefore, the above two band types represented Y1 and Y2 cattle haplogroups, respectively, indicating that Japanese black cattle had two patrilineal lineages with two taurine paternal origins. The haplotype diversities of Japanese black cattle and its hybrids were 0.5714 ± 0.0945 and 0.1988±0.1121 respectively, suggesting that Japanese black cattle has relatively high paternal genetic diversity. [Conclusion] Japanese black cattle had high paternal genetic diversity and two taurine paternal origins.
Japanese black cattle;Y chromosome;USP9Y;polymorphism;paternal origin
2016-12-20 接收日期:2017-01-05
国家肉牛牦牛产业技术体系(CARS-38)。
马志杰(1978-),男,甘肃省张家川县人,在读博士,副研究员,主要从事牦牛遗传育种研究。
*通讯作者:雷初朝(1968-),男,湖南常宁人,教授,博导,主要从事黄牛遗传资源研究。
S823
A
1001-9111(2017)01-0001-03
猜你喜欢
环球时报(2022-10-11)2022-10-11 17:16:53
中国饲料(2022年5期)2022-04-26 13:42:38
现代畜牧科技(2021年11期)2021-12-21 06:11:16
快乐语文(2021年30期)2021-11-24 13:21:50
贵州文史丛刊(2021年1期)2021-03-19 20:46:09
金桥(2021年2期)2021-03-19 08:34:16
中国牛业科学(2021年6期)2021-03-05 06:45:26
意林(2021年23期)2021-01-16 21:43:25
饲料博览(2016年2期)2016-04-05 22:42:40
植物营养与肥料学报(2012年1期)2012-10-26 02:49:46
