
旱涝交替胁迫对粳稻分蘖期叶片解剖结构的影响
2017-05-16 02:28:12陆红飞郭相平乔冬梅周新国王振昌刘春成
农业工程学报 2017年7期
陆红飞,郭相平,甄 博,乔冬梅,周新国,王振昌,刘春成,杨 泊
(1. 中国农业科学院农田灌溉研究所,新乡 453002; 2. 河海大学水利水电学院,南京 210098)
旱涝交替胁迫对粳稻分蘖期叶片解剖结构的影响
陆红飞1,2,郭相平2※,甄 博1,乔冬梅1,周新国1,王振昌2,刘春成1,杨 泊1
(1. 中国农业科学院农田灌溉研究所,新乡 453002; 2. 河海大学水利水电学院,南京 210098)
盆栽试验于2013年进行,在粳稻分蘖期分别设置涝-轻旱(LD)和涝-重旱(HD)2种处理(涝保持水深10 cm),以浅水勤灌(保持水深0~5 cm,CK)为对照,研究了旱涝交替胁迫对水稻叶片解剖结构的影响。结果表明,与CK相比,第1次旱5 d后(阶段I),LD处理主脉截面积减小了31.13%,LD处理侧叶大维管束面积和周长分别降低了28.87% 和15.79%,HD处理则分别降低了24.74%和13.16%(P<0.05);LD和HD处理侧叶小维管束面积和周长也显著减少。进入涝胁迫5 d后(阶段II),但LD处理小维管束周长较HD处理显著降低了0.13×102μm;HD处理除大维管束外其余指标均超过了CK,说明涝后重旱胁迫处理叶片发育表现出了一定的补偿效应。涝结束5 d后(阶段III),LD和HD处理主脉面积分别较CK显著减小了45.47%和53.79%,侧叶器官发育也落后于CK。第2次旱5 d后(阶段IV),LD和HD处理主脉大维管束面积分别较CK显著降低了18.45%和29.13%;HD处理侧叶大维管束面积和周长则显著低于CK和LD处理,HD处理侧叶面积较CK减少了34.49%,LD和HD处理泡面积在受旱时均低于CK,阶段III,泡面积则分别显著减少了0.38×102和0.57×102μm2。重旱处理对水稻叶片的迫害在第2次重旱后加重,而轻旱处理则减轻。研究结果可为制定水稻灌溉方案以及研究光合能力提供依据。
胁迫;干旱;细胞;水稻;维管束;叶片;旱涝交替胁迫
陆红飞,郭相平,甄 博,乔冬梅,周新国,王振昌,刘春成,杨 泊. 旱涝交替胁迫对粳稻分蘖期叶片解剖结构的影响[J]. 农业工程学报,2017,33(7):116-122.doi:10.11975/j.issn.1002-6819.2017.07.015 http://www.tcsae.org
Lu Hongfei, Guo Xiangping, Zhen Bo, Qiao Dongmei, Zhou Xinguo, Wang Zhenchang, Liu Chuncheng, Yang Bo. Effects of alternative stress of drought and waterlogging on rice leaf anatomical structure at tillering stage[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2017, 33(7): 116-122. (in Chinese with English abstract)doi:10.11975/j.issn.1002-6819.2017.07.015 http://www.tcsae.org
0 引 言
维管束是水稻植株的主要微观结构,茎、叶、叶鞘中都有维管束[1-2],其在植株体内进行长距离运输,承担了“源、库、流”中的“流”的功能[3]。穗颈节间、1次枝梗、叶片、2次枝梗、叶鞘等5个部位形态解剖性状关系紧密[4],且穗茎节部位的大维管束能通过分离和合并来调节生长[5]。水稻上部叶片、叶鞘中大维管束数均显著高于下部叶片[6],且叶鞘中的维管束数量稍多于叶片。叶脉是分布于叶肉组织中的维管束,是茎中维管束的分枝,起输导和支持的作用。一般而言,维管束越大、数目越多,养分供应就越好,有利于叶片的生长和光合速率的提高[7]。另外,叶片中大而多的气腔有利于叶部向下运送更多氧气至根部,以吸收更多的营养物质。叶中部和主脉一侧叶片发育状况也能够反映水稻光合以及运输养分的能力。
土壤水分对水稻各器官的生长及其功能起到重要作用。水稻的御旱性或受抑制程度与水稻根系、叶片形态结构的关系密切[8-9]。在受到旱胁迫之后,水稻叶片大、小维管束,泡状细胞数目均明显减少,可能引起产量降低[10-11],且维管束的大小能影响水分亏缺条件下水稻对水分的维持[12]。一般而言,在水稻幼苗期肥力对维管束的影响较为明显;与肥料全部基施相比,氮肥分开施用花序梗和叶片的导管总横截面积更大,并可增加分化阶段的导管数量和大小[13]。旗叶叶片和主茎鞘小维管束数目和横截面积均随着施氮水平的增加而增加[14]。水稻叶片主脉内的维管束、气腔的数目和大小或者叶肉细胞超微结构及叶绿体基粒片层结构差异可以作为水稻高产的评价指标。目前针对不同品种[15]、干旱和水作条件下[16-17]水稻叶片解剖结构的研究已取得诸多成果,也有学者从叶片解剖结构来研究旱稻抗旱特性,但对于旱-涝交替胁迫条件下水稻叶片解剖结构的研究较为少见。前人从根系解剖结构上已证明了旱涝交替胁迫可以增强水稻抗倒伏能力[18],针对茎解剖结构的研究[19]也表明短期旱胁迫可促进根系通气组织的发育,而不会造成水稻耐涝能力的降低。水稻分蘖期易受水肥等环境因素影响,完全展开叶的数目及发育情况对水稻干物质的积累起着重要作用。本文拟通过盆栽试验,研究旱涝交替胁迫对水稻分蘖期叶片解剖结构的影响,分析维管束、主脉等器官的发育状况,剖析水稻抗旱耐涝能力,以期为分析水稻光合能力、养分输送等提供依据。
1 材料与方法
1.1 试验地气象资料
试验地位于南京市江宁经济技术开发区,该地四季分明,属北亚热带季风气候区,年平均降水量为1 072.9 mm,平均年日照时数为2 017.2 h,年蒸发量为1 472.5 mm,年无霜期为 224 d,年平均气温为 15.7 ℃,年平均风速2.5 m/s。
1.2 试验设计
试验采用盆栽人工控水方式,于水稻分蘖期进行水分胁迫处理。水稻分蘖期试验方案为:设轻旱(LD)与重旱(HD)2个胁迫处理,均为旱-轻涝-旱连续胁迫各5 d,旱胁迫结束后立即转入涝胁迫,且涝之后待水分自然落干进入水分下限时第2次旱胁迫开始,CK水深维持在0~5 cm。参照文献[19-20]将田间持水率的60%作为水稻重度旱胁迫的水分下限,同时设置其水分上限为田间持水率的70%,而轻旱水分控制在田间持水率的70%~80%;轻涝为控制水深10 cm。水稻水分胁迫开始前与结束后水分管理同CK,除水分胁迫外,其他管理同大田。
试验采用的水稻品种为“南粳44”。2013年5月上旬育秧,待秧苗生长较为粗壮时(6月21日)选择大小相同的秧苗移栽(均为3叶1心),在每个盆里种3穴,共6株,每个处理6次重复。试验在河海大学南方地区高效灌排与农业水土环境教育部重点实验室玻璃温室内进行。5月初在试验地附件田地挖取供试土壤,经检测该处土壤为黏壤土,田间持水率为31.4%。将土平铺在温室平地上风干,经打碎、剔除石子并过0.5 mm筛后,装桶备用。
试验盆钵为塑料桶(从南京市润泰市场购置),其尺寸为:桶底、上口直径分别为18、24 cm,高25.5 cm。试验时每盆装干土7.5 kg(干土初始含水率为10.84%),干土中含全氮 60.1 mg/kg、速效氮 50.2 mg/kg、全磷39.4 mg/kg、速效磷9.58 mg/kg。施肥方案为:每盆施尿素(CO(NH2)2)、硫酸钾(K2SO4)、磷酸二氢钾(KH2PO4)和有机肥质量分别为1.63、0.65、1.9和12.5 g,有机肥含氮质量分数为3.75%,每盆总施氮质量为1.23 g(27.2 kg/hm2),所有肥料同时施入,将土与肥料搅拌均匀,并在大桶里摇匀后再装桶,轻微震荡,不压实,盆中土壤体积质量为1.29 g/cm3。采用称质量法控制土壤水分,上午、下午各称量1次。涝胁迫时,将6盆水稻一并放入水箱中,维持水桶水深高于盆中土壤表面高度10 cm。
1.3 叶片解剖结构观测方法和观测指标
叶片解剖结构观测方法:第一次旱5 d后(阶段I:7 月21日-7月25日)、涝5 d后(阶段II:7月26日-7月31日)、涝结束后5 d(阶段III:8月1日-8月5日)和第二次旱5 d后(阶段IV:8月7日-8月12日),取3盆长势一致的水稻,在每个盆中选取2片完全展开的倒二叶,共计 6片,用刀片切取叶片中部 8~10 mm叶段制作石蜡切片。叶片石蜡材料包埋、固定、切片、染色方法同文献[19]。采用奥林巴斯生产的OLYMPUS-BX41显微镜观察切片(20和40倍),通过Cell Sence Standard 软件量测大(小)维管束数目、大(小)维管束面积(μm2)和周长(µm)、主脉截面积(μm2)和周长(µm)、泡状细胞组面积(简称泡面积,μm2)和主脉一侧叶片(简称为侧叶)截面积(μm2),选择纹理清晰的切片拍照,大、小维管束数目、侧叶面积、主脉面积和周长均记录4组数据,主脉大、小维管束面积记录8组数据,侧叶大、小维管束和泡面积记录12组数据。
采用EXCEL和SPSS软件进行数据统计与分析。
2 结果与分析
2.1 水稻叶片解剖结构图片分析
不同处理水稻叶片20(主脉)和40倍(侧叶)解剖结构如图1所示。图1中绿色、蓝色部分分别为主脉大、小维管束,橙色部分为侧叶大维管束,紫色和红色部分分别为侧叶小维管束和泡状细胞组。从图1中可以看出,各处理叶片大小维管束、气腔、泡状细胞组均清晰可见,发育均比较完整,不同处理各器官外形不同,但无法判别处理间的差异性。因此,必须从周长、面积等指标进行细致分析。

图1 各处理叶片解剖结构Fig.1 Leaf anatomical structure of different treatments
2.2 分蘖期旱涝交替胁迫对水稻叶片主脉及大维管束面积、周长的影响
不同处理水稻叶片主脉和大维管束面积、周长见表1。由表1可知,第1次旱5 d后(阶段I),各处理叶片大维管束数目相同;LD处理大维管束面积和周长低于CK,但无显著差异,而HD处理则与CK相接近;与CK相比,LD处理主脉截面积显著减小了31.13%(P<0.05);LD 和HD处理主脉周长均较CK有所降低,但差异不显著。旱胁迫结束后,进入涝胁迫,5 d后(阶段II),各处理大维管束数目均为3个,但是LD和HD处理大维管束面积和周长均显著低于CK(P<0.05),其中LD和HD处理主脉大维管束面积较CK显著降低了22.22%和16.16% (P<0.05);LD和 HD处理主脉面积和周长也低于 CK (P>0.05);另外,LD处理大维管束和主脉的面积和周长与HD差异不显著(P>0.05),HD处理除大维管束外其余指标均超过了CK,说明涝后重旱胁迫处理叶片发育表现出了一定的补偿效应。
涝胁迫结束后5 d(土壤在逐渐干旱的过程中,此时盆中已无水层,阶段III),LD和HD处理大维管束数目与CK无显著差异;而LD和HD处理大维管束面积和周长相接近,但仍低于CK;而LD和HD处理主脉面积分别较CK显著减小了45.47%和53.79%(P<0.05),主脉周长则分别较CK减小了0.8×103和0.9×103μm(P<0.05),水分胁迫处理叶片主脉的发育状况远落后于CK。第二次旱5 d后(阶段IV),与CK相比,LD和HD处理大维管束面积分别降低了18.45%和29.13%(P<0.05),大维管束周长则分别降低了9.77%和16.71%(P<0.05),LD处理大维管束面积和周长均高于HD处理,但差异不显著;LD和HD处理主脉截面积和周长与CK差异不显著(P>0.05),此时,胁迫处理叶片主脉的发育状况接近于CK。
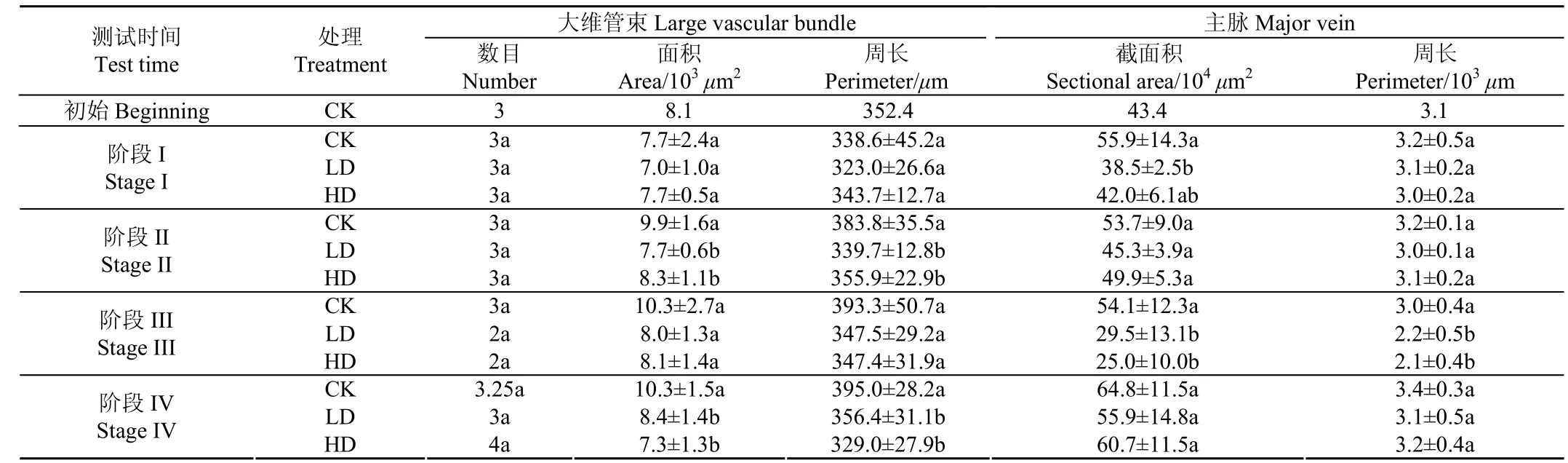
表1 水稻叶片主脉解剖结构统计结果Table1 Statistical results of anatomical structure of main veins in rice leaves
2.3 分蘖期旱涝交替胁迫对水稻主脉一侧解剖结构的影响
不同处理水稻叶片主脉一侧大小维管束发育状况、泡面积和侧叶面积如表2所示。由表2可知,阶段I,LD 和HD处理大维管束数目与CK无显著差异;与CK相比,LD处理大维管束面积和周长分别降低了 28.87%和15.79%(P<0.05),HD处理则分别降低了 24.74%和13.16%(P<0.05);而 LD和HD处理小维管束数目较CK分别减少了11.76%和4.81%(P>0.05),且二者小维管束面积和周长也显著低于 CK(P<0.05);LD和 HD处理泡面积和侧叶面积与CK无显著差异,而HD处理泡面积显著高于LD处理(P<0.05);此时,LD处理大(小)维管束面积和周长、侧叶面积均低于HD处理,且LD处理泡面积较HD处理显著降低了11.11%(P<0.05),说明水稻经历重旱胁迫(HD处理)后,叶片的发育状况优于轻旱胁迫(LD处理)。

表2 水稻叶片侧叶解剖结构统计结果Table2 Statistical results of anatomical structure of lateral leaf of rice leaves
阶段II,LD处理和HD处理大维管束数目与CK无显著差异;与CK相比,LD和HD处理大维管束面积分别显著降低了44.78%和39.55%(P<0.05),大维管束周长分别显著降低了21.43%和16.67%(P<0.05);LD和HD处理小维管束数目均低于CK,但差异不显著;各处理小维管束面积相接近;LD和HD处理小维管束周长与CK无显著差异,但 LD处理较 HD处理显著减少了0.13×102μm(P<0.05),二者泡面积和侧叶面积与CK也无显著差异;此时,LD处理侧叶解剖结构各项指标均低于CK和HD处理,而HD处理除大维管束及小维管束数目外其余指标均超过了CK,说明涝后重旱胁迫处理叶片发育表现出了一定的补偿效应。
阶段III,各处理大维管束数目与CK无显著差异;与CK相比,LD和HD处理大维管束面积分别减少了27.27%和31.82%(P<0.05),而周长则分别减少了13.89% 和16.67%(P<0.05);LD和HD处理小维管束数目和周长以及侧叶面积均低于 CK,但处理间无显著差异,而LD和 HD处理小维管束面积则分别较 CK显著减少了20.0%和 18.82%(P<0.05),泡面积则分别显著减少了0.38×102和0.57×102μm2(P<0.05),说明经过旱-涝胁迫后的一段时间内,水稻叶片的发育状况仍持续受到抑制;此时,LD处理叶片发育状况逐渐恢复,大维管束面积和周长、小维管束周长以及泡面积均超过了 HD处理,但LD侧叶面积低于HD处理。
阶段 IV,各处理大维管束数目表现与涝结束后 5 d时无显著差异;LD和HD处理大维管束面积和周长均低于 CK,其中 HD处理大维管束面积降低了 30.56% (P<0.05),但LD处理与CK无显著差异,而HD处理则显著低于CK和LD处理(P<0.05);不同于大维管束,LD和HD处理小维管束面积和周长接近CK,二者小维管束数目与CK无显著差异;LD处理泡面积较CK降低了10.33%(P<0.05),而HD处理则与CK无显著差异;HD处理侧叶面积较CK显著减少了34.49%(P<0.05);此时LD处理叶片发育状况得到加强,除侧叶面积及泡面积外,LD处理各指标与CK无显著差异。说明前期经历轻旱-涝胁迫后,水稻抗旱能力得到增强。
3 讨 论
水稻分蘖期对水分十分敏感,干旱和涝渍均会对水稻的生长产生不利影响[21-23]。已有研究表明水稻分蘖后期进行旱-轻涝不会引发叶片的衰老,却有助于根系活力的提高[20]。本试验研究发现,旱胁迫5 d后(阶段I),LD和HD处理主脉大维管束数目、主脉截面积和周长均低于常规灌溉(CK),这与张佳宇[16]的研究结果相似。旱-涝胁迫后(阶段II),LD和HD处理主脉大维管束面积和周长显著低于CK,这可能是因为旱涝双重胁迫对叶片的生长产生了明显的抑制。而在由涝转入第 2次旱的过程中(阶段III),LD和HD处理主脉大维管束的发育仍落后于 CK,且主脉面积和周长与 CK的差异显著(P<0.05),尤其是主脉截面积,水分胁迫处理仅为 CK的1/2左右,原因可能是经过旱涝双重胁迫后水稻植株的生长整体受到抑制,例如株高降低、分蘖减少等,叶片的发育也较慢,导致叶片比较瘦小。第2次旱胁迫后(阶段IV),LD和HD处理主脉大维管束面积和周长显著低于CK,但是HD处理大维管束数目超过了CK,此时主脉截面积和周长虽落后于CK,但差异不显著,说明水稻分蘖前期旱-涝胁迫并不会显著降低分蘖后期的抗旱能力。阶段I和阶段II,LD处理主脉解剖结构的发育均落后于HD处理,而进入阶段III时LD处理主脉截面积和周长,以及阶段IV LD处理大维管束面积和周长均超过HD处理,但是二者之间差异均不显著,说明重旱和轻旱对叶片主脉的影响差异并不明显,可能与主脉对干旱的敏感性有关。
旱涝交替胁迫减少了水稻叶片主脉一侧大小维管束数目,这与宋俊乔等[24]研究结果较为相似。阶段I至阶段II,LD和HD处理侧叶大维管束面积和周长均显著低于CK;阶段I时LD和HD处理侧叶小维管束面积和周长均显著低于CK,而阶段II时LD和HD处理小维管束发育得到恢复,尤其是HD处理小维管束面积和周长均超过了CK和LD处理,且周长显著高于CK,说明涝后水稻小维管束表现出一定的补偿效应。但是阶段I和阶段II,各处理泡面积和侧叶面积差异并不是十分显著,并且阶段II时HD处理侧叶面积较CK高10.94%,也表现出补偿效应。阶段III侧叶解剖结构的表现与主脉相似,大小维管束面积和周长、泡面积、侧叶面积均较CK有较大降幅,其中 LD处理小维管束面积和侧叶面积降幅分别为20%和23.20%,HD处理大维管束面积和泡面积分别降低了31.82%和25.00%,可能是由于叶片水分和养分得不到充分供给导致其发育不良。到阶段IV时,LD处理大维管束面积和周长虽仍低于CK,但差异不显著,HD处理则显著低于CK和LD处理,说明重旱对侧叶大维管束的抑制作用大于轻旱。阶段IV时LD和HD处理小维管束面积和周长与 CK无差异,但是泡面积和侧叶面积均较CK有较大差距,说明水稻叶片不同器官的抗旱特性表现不一。
综合水稻主脉和侧叶解剖结构的变化可以发现,从阶段I至阶段II,LD处理解剖结构相关指标先低于HD处理,阶段III至阶段IV,LD处理部分指标高于HD处理,且部分指标接近CK,说明水稻多次经历重旱不利于水稻叶片发育,而多次轻旱则对水稻叶片发育造成的不利影响相对较弱,同样轻旱对茎[18]和根[19]解剖结构的不利影响也弱于重旱。旱涝交替胁迫促进分蘖期根系的发育和通气组织的较早形成[19],对水稻根的生长发育产生了积极作用,而旱涝交替则限制了分蘖期叶片大小维管束的发育,不利于叶片生长,主要表现为叶面积的降低和叶片生长的萎缩[25]。这可能是因为干旱条件下,叶片失水较多,正常发育所需水分和养分得不到满足,而根系则不断调整以提高从干旱土壤中吸水的能力,并增强输水能力以维持水稻在逆境下的生长能力。
已有研究[26]表明分蘖期旱涝交替胁迫能显著降低光合速率、气孔导度,且分蘖期末轻旱和重旱处理叶片干物质量极显著低于CK[27],可能与叶片解剖结构的发育受限有关,尤其是与维管束面积的发育关系密切,这一点需要进一步进行光合指标和养分积累与解剖结构相关指标的相关性分析进行验证;另外,针对光合作用的研究也证明了前期受旱并不会降低后期水稻耐涝能力,前期轻度旱涝胁迫却能显著提高水稻后期抗旱能力。
分蘖期水稻叶片位置尚未完全固定(剑叶未展开),且不同叶位叶片解剖结构差异较大[28],这一点需要加强研究。徐静等[29]研究发现水稻窄叶基因主要通过调控生长素的合成与极性运输、维管组织的发育和分布,影响叶片维管束的数目。另外,分析叶片解剖结构的变化,可以反映水稻运输养分的能力,主要从“流”的角度来分析其对水稻光合作用的影响,但光合还受气孔[30]、叶绿体等器官的影响,因此需要加强叶片电镜扫描、化学微观成分、超微结构[7,15]、分子遗传学、大维管束和泡状细胞面积比以及泡状细胞下陷深度等方面的研究,并考察分蘖期旱涝交替胁迫结束后复水时水稻叶片在拔节期、抽穗期等的变化,以更加全面地评价旱涝交替胁迫下水稻叶片的发育。
4 结 论
1)水稻分蘖期旱胁迫能够显著限制侧叶大、小维管束的发育,但对主脉大维管束的影响较弱;旱-涝胁迫后主脉和侧叶大维管束的面积和周长显著降低,其中LD和HD处理主脉大维管束面积较CK显著降低了22.22%和16.16%(P<0.05),但侧叶小维管束、泡状细胞组、侧叶截面积的变化不明显;受旱后水稻仍然有一定的耐涝能力。
2)在水稻进入第2次干旱胁迫的过程中,前期叶片所受迫害仍未消除,主要表现在主脉和侧叶大、小维管束的面积和周长以及泡状细胞组面积显著降低,其中LD 和HD处理主脉面积较CK显著降低了45.47%和53.79%,而泡面积则分别减少了 0.38×102和 0.57×102μm2(P<0.05)。
3)旱-涝-旱胁迫后,主脉大维管束发育仍受到抑制,而轻旱处理(LD)侧叶除泡状细胞组面积显著降低了10.33%(P<0.05),其余叶片器官发育较为正常,但重旱处理(HD)侧叶大维管束和侧叶截面积显著降低了30.56%和34.49%。前期轻旱与涝胁迫并不会降低水稻后期的耐旱能力。
综合来看,水稻分蘖期经历 2次轻旱后叶片发育仍然比较良好,但应避免水稻经历多次重旱。
[1] Smillie I R, Pyke K A, Murchie E H. Variation in vein density and mesophyll cell architecture in a rice deletion mutant population[J]. Journal of Experimental Botany, 2012, 63(12): 4563-4570.
[2] 张英. 水稻维管束性状及与其相关性状分析[D]. 沈阳:沈阳农业大学,2005.
[3] 库尔萨诺夫. 植物体内同化物的运输[M]. 韩锦峰译. 北京:科学出版社,1986.
[4] 荆彦辉,徐正进. 水稻维管束性状的研究进展[J]. 沈阳农业大学学报,2003,34(6):467-471. Jing Yanhui, Xu Zhengjin. Research progress of rice vascular bundle characters[J]. Journal of Shenyang Agricultural University, 2003, 34(6): 467-471. (in Chinese with English abstract)
[5] 付娟,李霞,丁君辉,等. 水稻幼穗形态发生与维管束生长变化的研究[J]. 江西农业学报,2015,27(8):25-29. Fu Juan, Li Xia, Ding Junhui, et al. Study on morphogenesis of young panicles and growth change of vascular bundle in rice[J]. Acta Agriculturae Jiangxi, 2015, 27(8): 25-29. (in Chinese with English abstract)
[6] 徐正进,陈温福,曹洪任,等. 水稻穗颈维管束数与穗部性状关系的研究[J]. 作物学报,1998,24(1):47-54. Xu Zhengjin, Chen Wenfu, Chao Hongren, et al. Relation between the characters of panicle and vascular bundle in neck-panicle of rice[J]. Acta Agronomica Sinica, 1998, 24(1): 47-54. (in Chinese with English abstract)
[7] 王维金,徐珍秀. 不同品种水稻剑叶和穗颈大维管束和扫描电镜观察[J]. 华中农业大学学报,1994,13(5):518-520. Wang Weijin, Xu Zhenxiu. Examination to large vascular bundle of flag leaf and ear neck in different rice varieties by SEM[J]. Journal of Huazhong Agricultural University, 1994, 13(5): 518-520. (in Chinese with English abstract)
[8] 李芳兰,包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学报,2005,22(增刊):118-127. Li Fanglan, Bao Weikai. Responses of the morphological and anatomical structure ofthe plant leaf to environmental change[J]. Chinese Bulletin of Botany, 2005, 22(Supp.): 118-127. (in Chinese with English abstract)
[9] 陈展宇. 旱稻抗旱解剖结构及其生理特性的研究[D]. 长春:吉林农业大学,2008. Chen Zhanyu. Research Oil Anatomic Structure of Drought Resistance and Physiological Characteristic of Upland Rice[D]. Changchun: Jilin Agricultural University, 2008. (in Chinese with English abstract)
[10] 宋俊乔. 水稻叶片形态、解剖结构与抗旱性的关系研究[D].武汉:华中农业大学,2010. Song Junqiao. Research of the Relationship Between Morphologic and Anatomic Structure and Drought Resistance of Rice[D]. Wuhan: Huazhong Agricultural University, 2010. (in Chinese with English abstract)
[11] 戴常军. 土壤水分含量对水稻农艺性状及维管束系统分化的影响[J]. 吉林农业大学学报,2003,25(2):124-127. Dai Changjun. Effects of edaphic water content on agricultural characters and vascular bundle systems of rice[J]. Journal of Jilin Agricultural University, 2003, 25(2): 124-127. (in Chinese with English abstract)
[12] Sibounheuang V, Basnayake J, Fukai S. Genotypic consistency in the expression of leaf water potential in rice (Oryza sativaL.)[J]. Field Crops Research, 2006, 97(2/3): 142-154.
[13] Lee D J, Chae J C. Effect of nitrogen split application methods on development of vascular bundle and yield components of rice cultivars[J]. Korean Journal of Crop Science, 2000, 45(4): 237-240.
[14] Dong J L, Vergara B S, Zamora O B, et al. Development of small vascular bundles at different tillers as affected by nitrogen levels in rice plants[J]. Korean Journal of Crop Science, 1994, 39(3): 366-372.
[15] 杨艳华,王才林. 不同水稻品种叶片显微结构和超微结构的比较研究[J]. 植物研究,2010,30(2):152-156. Yang Yanhua, Wang Cailin. Comparative study on the microstructure and ultrastructure for three cultivated varieties of rice[J]. Bulletin of Botanical Research, 2010, 30(2): 152-156. (in Chinese with English abstract)
[16] 张佳宇. 水作和旱作水稻形态解剖结构及叶片气体交换特性的比较研究[D]. 长春:吉林农业大学,2008. Zhang Jiayu. Comparative Study of Rice Anatomical Structure and Leaves Gas Exchange Characteristics in Water Cultivation and in Dry Cultivation[D]. Changchun: Jilin Agricultural University, 2008. (in Chinese with English abstract)
[17] 胡雪琴,王三根. 6-BA对干旱胁迫水稻幼苗叶片显微结构的影响[J]. 西南大学学报:自然科学版,1999(6):505-509. Hu Xueqin, Wang San’gen. Effects of 6-BA on the microstructure of the leaf of rice seedings under water stress[J]. Journal of Southwest Agricultural University, 1999(6): 505-509. (in Chinese with English abstract)
[18] 郭相平,甄博,王振昌. 旱涝交替胁迫增强水稻抗倒伏性能[J]. 农业工程学报,2013,29(12):130-135. Guo Xiangping, Zhen Bo, Wang Zhenchang. Increasing lodging resistance performance of rice by alternating drought and flooding stress[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2013, 29(12): 130-135. (in Chinese with English abstract)
[19] 甄博,郭相平,陆红飞. 旱涝交替胁迫对水稻分蘖期根解剖结构的影响[J]. 农业工程学报,2015,31(9):107-113. Zhen Bo, Guo Xiangping, Lu Hongfei. Effects of alternative stress of drought and waterlogging at tillering stage on rice root anatomical structure[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2015, 31(9): 107-113. (in Chinese with English abstract)
[20] 郭相平,袁静,郭枫,等. 旱涝快速转换对分蘖后期水稻生理特性的影响[J]. 河海大学学报:自然科学版,2008,36(4):516-519. Guo Xiangping, Yuan Jing, Guo Feng, et al. Effects of rapid shift from drought to waterlogging stress on physiological characteristics of rice in late tillering stage[J]. Journal of Hohai University: Natural Sciences, 2008, 36(4): 516-519. (in Chinese with English abstract)
[21] 邵玺文,阮长春,赵兰坡,等. 分蘖期水分胁迫对水稻生长发育及产量的影响[J]. 吉林农业大学学报,2005,27(1):6-10. Shao Xiwen, Ruan Changchun, Zhao Lanpo, et al. Effects of water stress on growth and yield of rice in tillering stage[J]. Journal of Jilin Agricultural University, 2005, 27(1): 6-10. (in Chinese with English abstract)
[22] 赵正宜,迟道才. 水分胁迫对水稻生长发育影响的研究[J].沈阳农业大学学报,2000,31(2):214-217. Zhao Zhengyi, Chi Daocai. Effect of soil water stress on rice growing and rice water production function[J]. Journal of Shenyang Agricultural University, 2000, 31(2): 214-217. (in Chinese with English abstract)
[23] 杨伟利,俞双恩,赵方波. 不同受涝模式下水稻抽穗开花期荧光参数的日变化规律[J]. 灌溉排水学报,2011,30(2):27-31. Yang Weili, Yu Shuang’en, Zhao Fangbo. The impact of varying farmland water logged of rice to the parameters of diurnal chlorophyll fluorescence kinetics in heading and flowering stage[J]. Journal of Irrigation and Drainage, 2011, 30(2): 27-31. (in Chinese with English abstract)
[24] 宋俊乔,陈亮,楼巧君,等. 水稻叶片形态解剖结构在水旱两种条件下的多样性分析[C]//全国生物遗传多样性高峰论坛会刊,2012.
[25] 陆红飞,郭相平,王振昌,等. 分蘖期旱涝交替胁迫对水稻叶片性状的影响[J]. 灌溉排水学报,2017,36(1):47-51. Lu Hongfei, Guo Xiangping, Wang Zhenchang, et al. Effects of alternative stress of drought and waterlogging on leaf character[J]. Journal of Irrigation and Drainage, 2017, 36(1): 47-51. (in Chinese with English abstract)
[26] 陆红飞,郭相平,甄博,等. 旱涝交替胁迫条件下粳稻叶片光合特性[J]. 农业工程学报,2016,32(8):105-112. Lu Hongfei, Guo Xiangping, Zhen Bo, et al. Photosynthetic characteristics of Japonica rice leave under alternative stress of drought and waterlogging[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2016, 32(8): 105-112. (in Chinese with English abstract)
[27] 郭相平,李小朴,陆红飞,等. 水稻分蘖期旱涝交替胁迫对干物质累积及氮素吸收的影响[J]. 灌溉排水学报,2015,34(2):20-24. Guo Xiangping, Li Xiaopu, Lu Hongfei, et al. Study on dry matter accumulation and nitrogen uptake under alternating drought and flooding stress in rice tillering stage[J]. Journal of Irrigation and Drainage, 2015, 34(2): 20-24. (in Chinese with English abstract)
[28] 林长茂. 大穗型水稻叶片形态解剖的初步研究[J]. 江西农业大学学报,1988(增刊2):73-84.
[29] 徐静,王莉,钱前,等. 水稻叶片形态建成分子调控机制研究进展[J]. 作物学报,2013,39(5):767-774. Xu Jing, Wang Li, Qian Qian, et al. Research advance in molecule regulation mechanism of leaf morphogenesis in rice (Oryza sativaL.) [J]. Acta Agronomica Sinica, 2013, 39(5): 767-774. (in Chinese with English abstract)
[30] 单提波,徐正进. 不同气孔密度水稻叶片光合及叶绿素荧光参数的日变化特征[J]. 沈阳农业大学学报, 2015, 46(2):129-134. Shan Tibo, Xu Zhengjin. Daily dynamics of photosynthesis and chlorophyll fluorescence parameters in rice with different stomata densities[J]. Journal of Shenyang Agricultural University, 2015, 46(2):129-134.
Effects of alternative stress of drought and waterlogging on rice leaf anatomical structure at tillering stage
Lu Hongfei1,2, Guo Xiangping2※, Zhen Bo1, Qiao Dongmei1, Zhou Xinguo1, Wang Zhenchang2, Liu Chuncheng1, Yang Bo1
(1.Farmland Irrigation Research Institute, Chinese Academy of Agricultural Sciences, Xinxiang453002,China; 2.College of Water Conservancy and Hydropower Engineering, Hohai University, Nanjing210098,China)
Vascular bundle is the main microstructure of rice plants, which is conducive to the growth of leaf and photosynthetic rate, and the greater the vascular bundle, the more the number, the better the supply of nutrients. Pot experiments were performed to research the effects of alternative stress of drought and waterlogging at tillering stage on rice leaf anatomical structure, including the number, area and perimeter of large (small) vascular bundles, sectional area and perimeter of major vein, area of bulliform cell group and sectional area of side blade, and 2 treatments were set up in 2013: waterlogging - light drought (LD) and waterlogging - severe drought (HD), in which the water depth of waterlogging was 10 cm, using regular irrigation (CK) as the control. The experiments were conducted in the greenhouse of Key Laboratory of Efficient Irrigation-Drainage and Agricultural Soil-Water Environment in Southern China (Hohai University), Ministry of Education. The results showed that alternative stress of drought and waterlogging decreased the number of large (small) vascular bundles, but there was no significant difference compared to the CK; after the first drought for 5 d (Stage I), the sectional area of major vein of LD treatment was reduced by 31.13% compared to the CK (P<0.05), and that of HD treatment was reduced by 13.9×104μm2, which yet had no significant difference with the CK; the area and perimeter of large vascular bundle of side blade were reduced by 28.87% and 15.79% for LD treatment, respectively, and by 24.74% and 13.16% for HD treatment, respectively, and the area and perimeter of small vascular bundle of side blade for LD and HD treatments were also lower than that of the CK. After waterlogging for 5 d (Stage II), the perimeter of small vascular bundle of side blade for LD and HD treatments had no significant difference with the CK, and that of LD treatment was reduced by 0.13×102μm compared to HD treatment. Except large vascular bundle, other indices of HD treatment were all over than the CK, indicating that leaf development showed a certain compensation effect. Five days after waterlogging (Stage III), the sectional area of major vein of LD and HD treatments was significantly reduced by 24.6×104and 29.1×104μm2respectively compared to the CK, and the development of side blade organ was also behind the CK. After the second drought for 5 d (Stage IV), the area of large vascular bundle of major vein for LD and HD treatments was significantly reduced by 19.42% and 29.13% respectively compared to the CK; the area and perimeter of large vascular bundle of side blade under LD treatment had no significant difference with the CK, and those under HD treatment were significantly lower than LD treatment and CK. The areas of bulliform cell group of LD and HD treatments were lower than the CK when the rice was in drought, and especially in Stage III, the areas of bulliform cell group were significantly reduced by 0.38×102and 0.57×102μm2, respectively; the sectional areas of side blade of LD and HD treatments in Stage I, III and IV were lower than the CK, while in Stage II, the sectional area of side blade of HD treatment was higher than the CK, but the difference was not significant. After the water stress, the development of the main vein’s large vascular bundle was inhibited, and the development of side blade was also subject to certain persecution, but the persecution of rice leaves under severe drought treatment was increased after the second drought stage, while it was reduced under light drought treatment, indicating that the rice drought tolerance was enhanced, and after the drought stress, the waterlogging tolerance of rice was still strong. The results of this study can provide the basis for the development of appropriate water management plan and the research of rice photosynthesis.
stresses; drought; cells; rice; vascular bundle; leaf; alternative stress of drought and waterlogging
10.11975/j.issn.1002-6819.2017.07.015
S274.1
A
1002-6819(2017)-07-0116-07
2016-10-09
2017-04-01
国家自然科学基金项目(51070942、51309080);“十二五”公益性行业(农业)科研专项经费资助项目(201203077、201203032);中央级科研院所基本科研业务费专项 (中国农业科学院农田灌溉研究所) 资助项目(1610262016028);江苏省水利科技项目
陆红飞,江苏淮安人,助理研究员,主要从事节水灌溉理论与技术研究。新乡 中国农业科学院农田灌溉研究所,453002。
Email:gofeigo@sina.com
※通信作者:郭相平,山东人,教授,博士,主要从事农业水土工程研究。南京 河海大学水利水电学院,210098。
Email:xpguo@hhu.edu.cn
猜你喜欢
河北农业大学学报(2021年5期)2021-11-10 13:11:28
南方农业学报(2021年2期)2021-06-30 01:56:03
新疆农业科学(2020年9期)2020-10-13 12:37:42
成都信息工程大学学报(2020年2期)2020-08-24 08:05:50
湖南文理学院学报(自然科学版)(2020年2期)2020-05-08 08:00:14
河南农业科学(2019年9期)2019-09-24 09:10:48
新课程·小学(2019年7期)2019-09-17 12:58:34
江苏农业科学(2019年21期)2019-01-03 02:05:58
水利科技与经济(2016年5期)2016-04-22 03:43:40
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23 00:59:48
