
流行性2型PRRSV强毒株的广谱中和抗体研究
2017-05-13 02:43
猪业科学 2017年4期
(勃林格殷格翰国际贸易有限公司,北京 100027)
流行性2型PRRSV强毒株的广谱中和抗体研究
陈 锴,黄亚平(译);徐大为,朱连德(校)
(勃林格殷格翰国际贸易有限公司,北京 100027)
由于疫苗需要提供有效的交叉保护力对抗变异较大的猪繁殖与呼吸综合征病毒(PRRSV)毒株,获得可预测PRRSV免疫保护力的评估指标可促进群体管理改善和疫苗研发项目的开展。中和抗体在动物和人的抗病毒免疫中起着关键作用,故通过特异性抗体中和病毒感染性的中和效力是免疫保护的常见预测因子。然而,由于病毒中和作用不是抗PRRSV免疫应答的标志性和共同特征,它们抗PRRSV的保护作用被打了折扣。例如,众多关于生长猪的研究表明,PRRSV感染后数周至数月无中和抗体产生或以低于1/8滴度的低效价存在。目前,在PRRSV强毒暴发或有重复血清接种史的母猪群体中发现了具有广谱反应能力的高滴度中和抗体。进一步的研究显示,这些具有交叉反应能力的中和抗体是所有受检猪场母猪的共同特征,这些现象出现在免疫场或野毒感染场,与胎次无关,交叉反应谱差异大,且在相同猪场的不同母猪个体中存在较大变化。例如,在一个常规接种疫苗猪场的所有母猪(N=20)均对VR2332具有同源血清中和活性,滴度范围为15~256;90%的母猪对MN184具有滴度范围为4~60的异源中和活性;20%的母猪对1型PRRSV SDEU具有滴度范围为4~8的异源中和活性。在使用中和反应检测高毒力毒株PRRSV174时,发现其生长特征不同于所有其他病毒。在细胞系中加入相同拷贝RNA的情况下,使用TCID50检测显示毒株MW174的感染活性比其他毒株低约1 000倍。由于TCID50是基于细胞病变效应,提示这种高毒力毒株家族可延迟或阻断细胞裂解从而获得更好的生长。来自两个MW174毒株病毒暴发群体中的所有10头母猪的血清均具有针对VR2332、MN184、MW174、NC134和1型SDEU的中和活性。在22%的样品(N=50,每种病毒10个血清样品)中和反应滴度效价大于256,由于广谱中和反应血清可显著降低异源PRRSV的感染,交叉抗体中和反应活性非常重要。这些发现支持了PRRSV交叉保护的理论,表明免疫应答相关的保守表位存在于1型和2型PRRSV毒株中。
1 实验动物
血清样品收集于美国明尼苏达州和北卡罗来纳州的独立商品代母猪群,这些种群被PRRSV毒株多次感染,感染途径为疫苗免疫、自然感染或血清接种。血清样品采集自:被多次不同毒力毒株感染的淘汰母猪、免疫过疫苗但未被强毒力毒株感染过的不同胎次母猪;使用强毒力PRRSV毒株对后备母猪进行可控而系统性的驯化,实验性感染VR2332毒株后202 d,及来自具有较高病毒中和滴度的商品母猪群。
2 细胞培养
通过灌洗各日龄动物肺脏,但主要从5~6周龄保育猪收集猪肺泡巨噬细胞(PAM)。 用含有50 μg/mL庆大霉素的磷酸盐缓冲液(PBS)进行灌洗,并将红细胞在无菌水中洗脱。使用70 μm过滤器过滤分离物,并重悬于采用40%RPMI培养基,50%胎牛血清(FBS)和10%二甲基亚砜(Fisher Scientific,Pittsburgh,PA)配置的溶液中,冷冻保存。用RPMI培养基(CellgroMediatech,Manassas,VA)培养PAM,并添加丙酮酸钠、硫酸庆大霉素、非必需氨基酸、HEPES缓冲液和5%FBS(Sigma,St.Louis,MO)。使用MEM培养基(Gibco,Grand Island NY)培养猿猴肾上皮MARC145细胞,并添加碳酸氢钠、非必需氨基酸、HEPES缓冲液(Sigma,St.Louis,MO)、硫酸庆大霉素(CellgroMediatech,Manassas,CA)和10%FBS(Sigma,St.Louis,MO)。
3 PRRSV毒株
PRRSV分离株VR2332,MN184,SDEU,MW174,NC134接种并收获于MARC145细胞,通过监测96孔板中MARC145细胞的病变效应,使用Reed&Muench方法计算每个分离株的半数感染量(TCID50)。从两个强毒力PRRSV流行毒株MN0111和MN1211上获得了其ORF5的核苷酸序列信息(编码糖蛋白5),但是并未分离到病毒。使用CLC Sequence Vie wer version 6.8.2(CLC bio/Qiagen,Cambridge,MA)对实验室改编毒株VR2332,疫苗株,MN184,1型PRRSV病毒 LelystadVirus (LV)株和1型田间分离株SDEU进行序列比对分析,并在使用Geneious R6Tree Builder(Biomatters Ltd.,Auckland,New Zealand,版本6.1.7)构建系统发生树,bootstrap设置为100,不使用外参基因。同时使用Geneious R6分析不同毒株之间同源性差异百分比。
4 基于ELISA的血清中和抗体测定
以如下方法进行PRRSV血清中和(SN)测定。将96孔板用25 μg/mL聚乙烯亚胺(Sigma-Aldrich,St.Louis,MO)处理,以6×104个细胞/孔接种PAM并培养48 h,或以6×103个细胞/孔接种MARC145细胞。将血清样品热灭活(56 ℃,30 min)并使用不含FBS的培养基进行连续稀释。将稀释后的血清与等体积的5×104TCID50/ mL PRRSV病毒在非贴壁96孔板中混合,并孵育1 h(37 ℃,5%CO2)。将含有5 000 TCID50的血清-病毒混合物转移至细胞孔中,在37℃下吸附1 h。PBS洗涤细胞,加入培养基,并进一步温育24~48 h。使用含有3.7%甲醛(Sigma-Aldrich,St.Louis,MO)的PBS固定,随后洗涤,透化(PBS中的0.1%Triton X-100(Sigma-Aldrich,St。Louis,MO)),并封闭1 h(5%脱脂奶粉PBS)。使用含5%脱脂奶粉的PBS以1:10 000稀释抗PRRSV核衣壳蛋白的单克隆抗体SR30-A(Rural Technologies,Brookings,SD),并以100 μL /孔加入细胞孔,孵育1 h。以100 μL/孔加入,以1:10 000稀释羊抗鼠HRP(H+L),反应1 h。加入100 μLTMB(KirkegaardPerry Laboratories,Gaithersburg,MD)过氧化物酶底物进行显色,作用15 min。最后用100μL的1M磷酸终止反应,平板使用Millermax酶标仪(Molecular Devices,Sunnyvale,CA)于450nm进行读数。在每个稀释度下的病毒抑制比例与扣除空白背景吸光度的阳性病毒感染对照进行对比。计算出能保护50%细胞孔不产生细胞病变的血清稀释度,该稀释度即为该份血清的中和抗体效价。
5 结果与讨论
通常认为,PRRSV感染后体液免疫应答较弱,中和抗体水平效价低且出现晚,出现在感染后期急性病毒血症已经减弱时,或者根本不出现。目前这些类型的研究主要基于幼龄动物的攻毒实验。这些研究还发现,中和抗体在淋巴组织的持续性感染过程中效价也未提高至更高水平。但是,这些中和反应实验并未在成年猪上进行研究,成年猪经历多次感染或抗原接触,并随着动物年龄的增长和时间推移可能会发生免疫变化。我们假设有PRRSV感染史的成年猪的中和抗体反应是不同的,首先需要从有PRRSV暴发史的商品代母猪场中分离并检测PRRSV中和抗体。
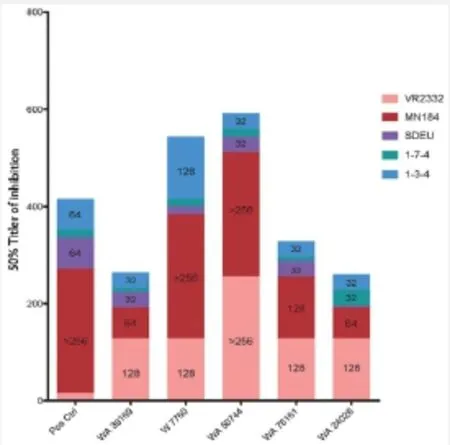
图1 5个从174 RFLP型PRRSV暴发场随机抽取血清样品的广谱中和反应滴度
在两个场的多胎次母猪中发现了高均匀度的高效价中和抗体,此中和抗体可与2型毒株VR2332反应,未知是否接触相关疫苗株Ingelvac MLV。2个群体间最大中和作用是相同的,群体1 中50%中和效力的滴度为512,最高个体值达2 187,群体2中50%中和效力滴度为81。中和活力与剂量相关,曲线结构在两个群体间也类似。
南达科他州立大学Eric Nelson应用目前广泛使用的荧光聚焦中和实验(F lurescent Focus Neutralization, FFN)对来自一个群体的5个血清样品使用多个1型和2型PRRSV分离株进行检测。血清与90%的2型PRRSV毒株发生反应,滴度为16~56;并意外发现可与90%的I型PRRSV发生反应,滴度大于80。广谱中和活性与胎次并无相关性。高滴度中和抗体也在每月使用PRRSV强毒株驯化的后备母猪上发现。结果表明:广谱中和抗体可存在于各种不同生产日龄和不同PRRSV感染群体,包括生长猪、成年猪、强毒力感染和弱毒疫苗免疫。
中和抗体在同一猪场和有感染史猪群间变异系数均较大。例如,在一个常规接种疫苗猪场的所有母猪(N=20)均对VR2332具有同源血清中和活性,滴度范围为15~256;90%的母猪对MN184具有滴度范围为4~60的异源中和活性;20%的母猪对MN184具有滴度范围为4~60的异源中和活性;20%的母猪对1型PRRSV SDEU具有滴度范围为4~8的异源中和活性。在使用中和反应检测强毒株MW174时,发现其生长特征不同于所有其他病毒。在细胞系中加入相同拷贝RNA的情况下,使用TCID50检测显示毒株MW174的感染活性比其他毒株约低1 000倍。由于TCID50是基于细胞病变效应,提示这种高毒力毒株家族可延迟或阻断细胞裂解从而获得更好的生长。从两个174毒株暴发场收集的10母猪样品显示,对VR2332、MN184、MW174、NC134和1型毒株SDEU均有中和反应活性;22%的样品(N=50,每种病毒10个血清样品)中和反应滴度大于256。图1显示了同一猪场中5头不同母猪血清对上述病毒的中和反应滴度。
由于广谱中和反应血清可显著降低异源PRRSV的感染,因此交叉中和反应活性非常重要。这些发现支持了PRRSV交叉保护的基础理论,表明免疫相关的保守表位存在于1型和2型PRRSV中。
注:本文选自2017-AASV论文集
2017-03-08)
猜你喜欢
动物医学进展(2022年9期)2022-11-26
昆明医科大学学报(2022年2期)2022-03-29
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
华侨大学学报(自然科学版)(2021年4期)2021-07-30
中国医药科学(2021年7期)2021-05-11
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
中国抗生素杂志(2019年6期)2019-07-06
中国人兽共患病学报(2017年11期)2017-12-13
