
短季棉生育期性状的遗传分析
2017-04-13 06:56田锋范术丽魏恒玲王寒涛赵树琪庞朝友胡守林喻树迅
新疆农业科学 2017年2期
田锋,范术丽,魏恒玲,王寒涛,赵树琪,3,庞朝友,胡守林,喻树迅
(1.塔里木大学植物科学学院,新疆阿拉尔 843300;2.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳 455000;3.黄冈市农业科学院,湖北黄冈 438000)
短季棉生育期性状的遗传分析
田锋1,2,范术丽2,魏恒玲2,王寒涛2,赵树琪2,3,庞朝友2,胡守林1,喻树迅2
(1.塔里木大学植物科学学院,新疆阿拉尔 843300;2.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳 455000;3.黄冈市农业科学院,湖北黄冈 438000)
【目的】研究短季棉生育期的遗传规律,为棉花早熟性状QTL定位和分子标记辅助育种提供理论基础。【方法】利用四个陆地棉品种构建两个杂交群体,采用主基因+多基因联合世代分析,对棉花早熟性相关的播种-开花(Flowering time,FT),花铃期(Flowering and boll-setting period, FBP),全生育期(Whole growing period,WGP)这三个性状进行遗传分析。【结果】两个组合,除组合Ⅱ中F1的花铃期和F2的播种-开花期偏向晚熟亲本外,其他表型性状均偏向早熟亲本,但总体偏早熟亲本;组合Ⅰ及组合Ⅱ的相同性状均符合同一个遗传模型,且均受两对主基因控制。【结论】三个表型性状是由主基因和多个微效基因共同作用,且主要由主基因控制;三个表型性状的遗传力高,且加性效应大。
短季棉;生育期;主基因+多基因;遗传分析
0 引 言
【研究意义】棉花一直是我国重要的经济作物,棉花种植业的发展关系到纺织工业的兴衰,同时也涉及到我国近一亿棉农及两千多万纺织工人的就业、再就业问题[1-2]。人多地少,粮棉争地的问题一直存在,这使棉花种植面积一再缩减,导致国内棉花产量无法满足内需,还需要向外大量进口[3]。缓解粮棉争地矛盾,解决我国棉花供需以及粮食需求的一个有效方法就是提高粮棉复种指数,实现粮棉由一熟向两熟发展[4]。因此,选育生育期较短,品种优良的短季棉品种势在必行[5]。棉花早熟相关性状是复杂的数量性状,受到多个数量性状位点和环境的共同影响,了解棉花关于早熟的遗传规律,对培育短季棉品种具有的指导意义。【前人研究进展】前人根据数量遗传学的方法,从整体上分析了某一性状的多个微效基因的遗传效应,但无法解析单一基因的遗传效应[6-7]。在相关棉花早熟性的遗传分析中,范术丽[8]利用两个组合的四个世代群体,对棉花早熟相关性状主基因-多基因的遗传规律进行了分析,揭示了早熟相关性状主基因存在的普遍性。董娜[7]根据六个世代,利用了主基因-多基因混合遗传模型分析方法,说明早熟相关性状的基因表达很容易受到环境的影响。【本研究切入点】喻树迅等[9]在对所推广的早熟棉品种进行的遗传相关分析中提出,生育期可以作为早熟性的指示性状。艾先涛等[10]认为,通过品种遗传多样性的分析可以减少育种过程中的盲目性与重复性。利用多世代联合遗传分析,对棉花早熟性状进行分析,确定控制早熟性基因的作用方式。【拟解决的关键问题】研究采用主基因-多基因混合遗传模型对组合Ⅰ(中棉所60×中棉所50)和组合Ⅱ(中棉所41×中棉所74)的四个世代群体P1、P2、F1、F2,对棉花生育期三个早熟相关性状的遗传规律进行分析,明确棉花早熟性状的遗传模型及基因作用方式,估测出两组合的主基因遗传效应和遗传力,为棉花早熟性状QTL定位及分子标记辅助育种提供理论基础[10]。
1 材料与方法
1.1 材 料
选用早熟棉品种和中熟棉品种为亲本构建两个分离群体(组合Ⅰ:中棉所60×中棉所50;组合Ⅱ:中棉所41×中棉所74)。2014年在安阳配置两个杂交组合,获得F1代种子,同年将获得的F1代种子在海南进行加代,自交得到F2代种子。2015年5月25日在河南安阳种植两个杂交组合的亲本及其F1各3行,种植组合Ⅰ的F2分离群体414株,种植组合Ⅱ的F2分离群体468株。
1.2 方 法
1.2.1 性状调查
对两杂交组合的亲本、F1及F2群体进行单株调查。亲本及F1代植株各调查30个单株,其F2分离群体单株全部调查。调查性状包括:播种-开花(Flowering time,FT)、花铃期(Flowering and boll-setting period, FBP)、全生育期(Whole growing period,WGP)3个表型性状。
1.2.2 遗传模型及参数估计
采用章元明等[11]提出的利用P1、P2、F1、F2四个世代的主基因+多基因混合遗传分析法对单株表型数据进行分析。根据所计算出的各遗传模型AIC值(Akaike's Information Criterion,AIC)来确定候选基因型,进一步对候选基因模型进行均匀性检验、Smirnov检验以及Kolmogorov检验,最终根据检验结果选定最适模型,并对估算出的各项遗传参数进行分析。
2 结果与分析
2.1 表型性状特性
研究表明,组合Ⅰ中的母本相对于父本为晚熟品种,并且两个亲本间播种-开花时间相差11.57 d,花铃期相差13 d,全生育期相差24.57 d;组合Ⅱ的母本为晚熟品种,父本为早熟品种,调查数据显示,两个亲本间播种-开花相差7.44 d,花铃期相差8.16 d ,全生育期相差15.6 d;两个组合亲本之间的差异有统计学意义。从表型数据可以看出,组合Ⅰ中F1、F2表型偏向早熟亲本中棉所50,由此可以肯定早熟亲本的早熟性在后代群体中部分呈显性遗传;组合Ⅱ中除F1的花铃期和F2的播种-开花期偏向晚熟亲本中棉所41,其它性状的都偏向早熟亲本,对两个组合的总体而言,群体表型偏向早熟亲本,在后代群体中,早熟性部分呈显性遗传。表1
表1 各世代表型特征
2.2 遗传模型的选择和适合性检测
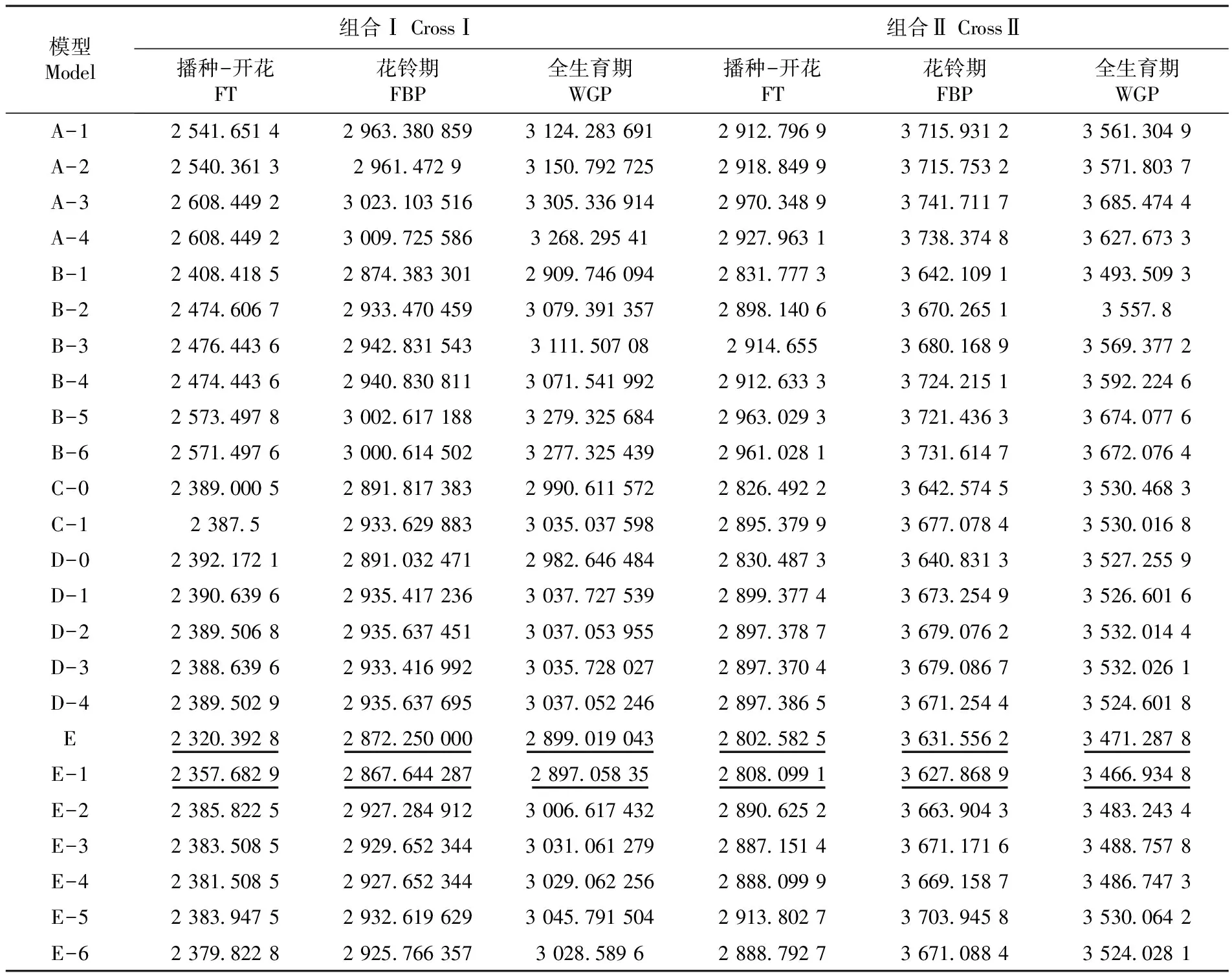
采用四世代联合分析的方法,根据两个组合的播种-开花、花铃期、全生育期的田间表型数据估算出24种遗传模型的AIC值,并根据最小AIC值的原则在每个指标中选取两个候选模型。结果显示两个组合的播种-开花、全生育期、花铃期的候选模型全都为E和E-1。通过对以上候选的遗传模型进行适合性检验,结合检验结果确定最适遗传模型。表2
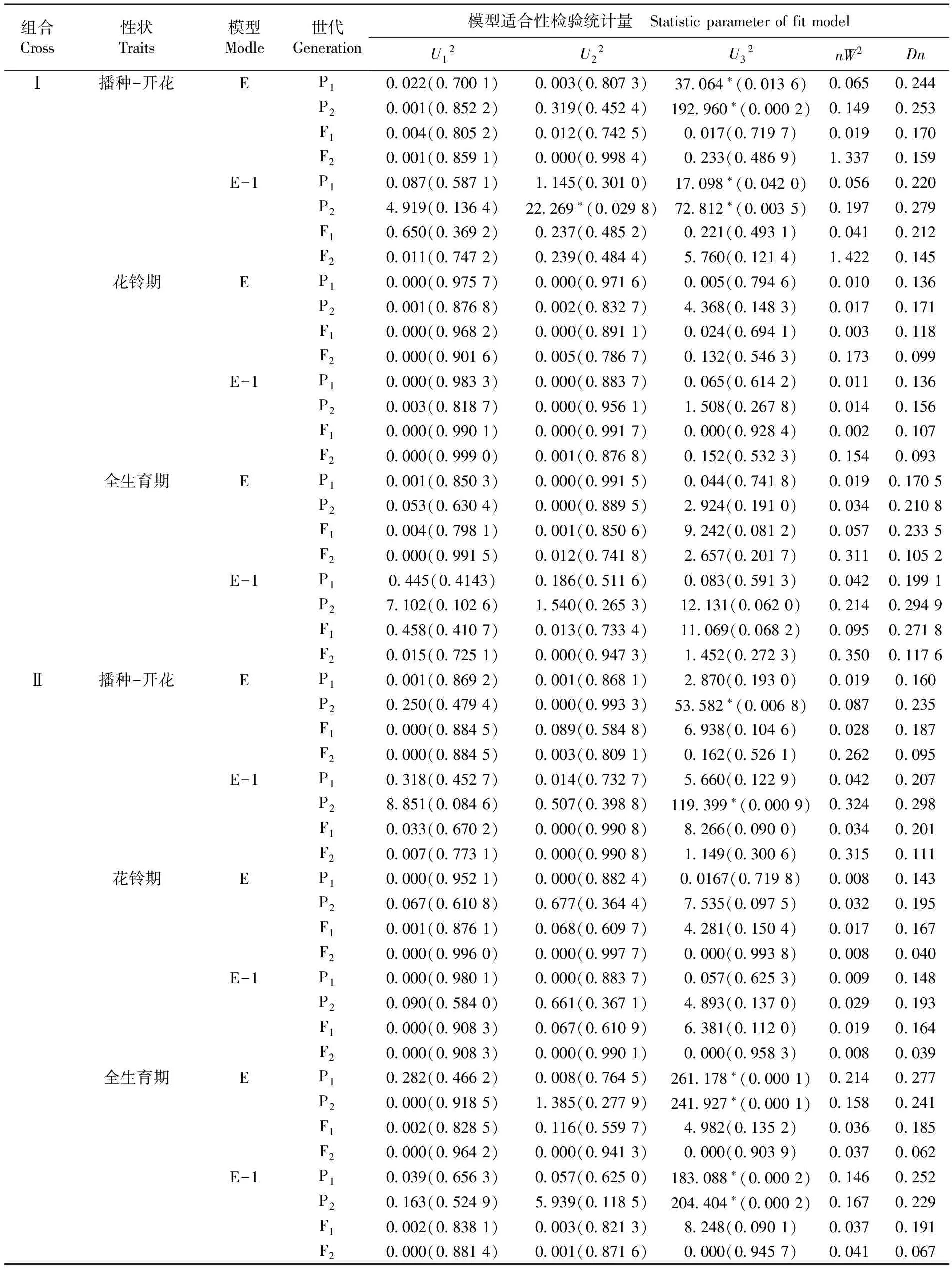
适合性检验结果所示,根据两个组合不同生育期性状模型的统计量达到的显著水平及最小AIC的原则,确定组合Ⅰ、组合Ⅱ不同生育期的最适遗传模型。两个杂交组合的播种-开花及全生育期的最适遗传模型均符合模型E(两对加性-显性-上位性主基因+加性-显性-上位性多基因模型);花铃期的最适遗传模型也都符合模型E-1(即两对加性-显性-上位性主基因+加性-显性多基因模型)。根据最适模型可以看出,组合Ⅰ及组合Ⅱ各生育期的最适遗传模型是相一致的,两个杂交组合在生育期的遗传性状稳定,这三个生育期性状可以作为早熟性的指示性状。表3
2.3 遗传参数的估计
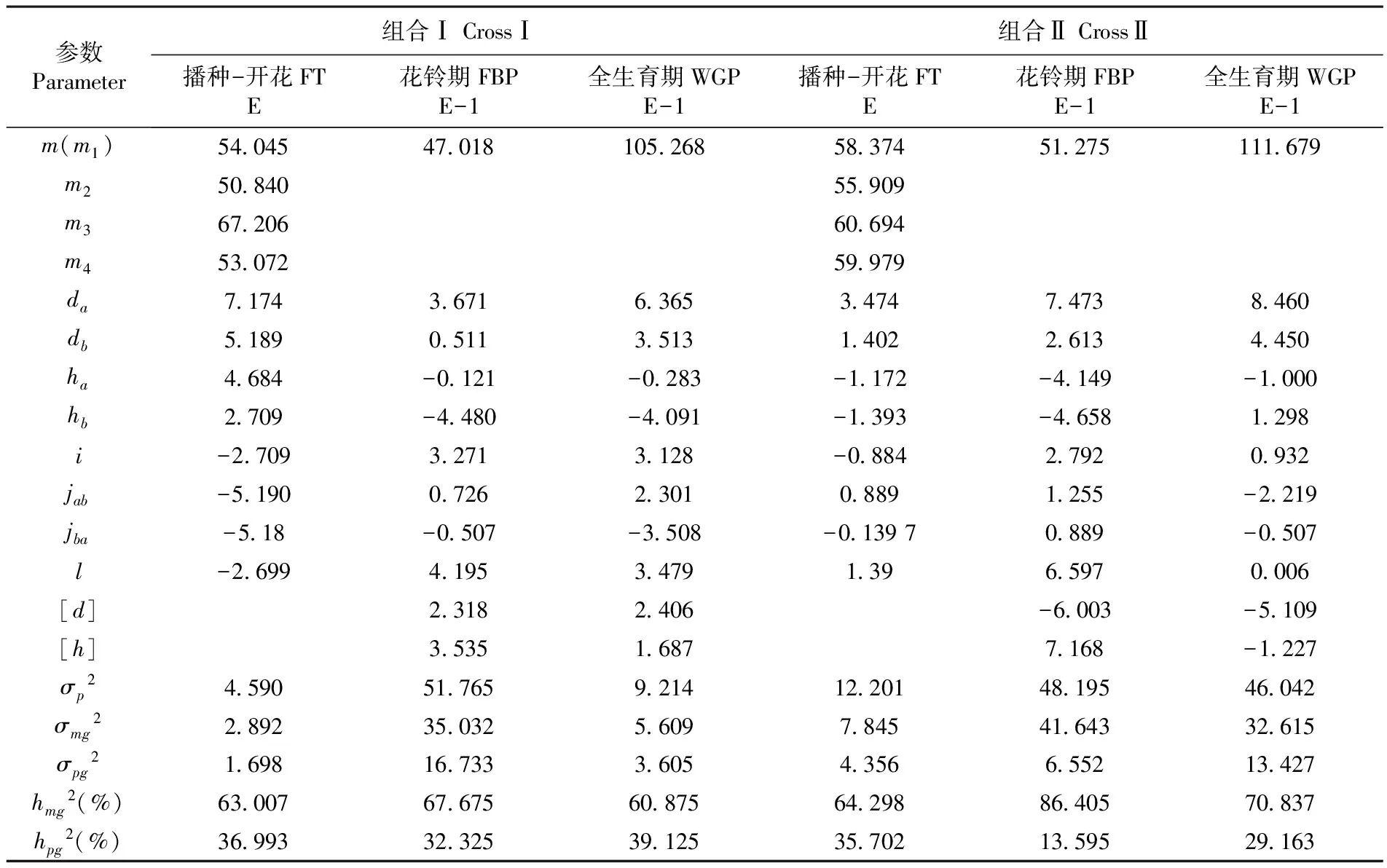
利用最小二阶乘法分别估计组合Ⅰ及组合Ⅱ播种-开花、花铃期、全生育期最适遗传模型的一阶参数和二阶参数,分析两个杂交组合各个生育时期的遗传规律。表4
2.3.1 播种-开花主-多基因的遗传分析
通过最优遗传模型E(两对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型)对两个组合播种-开花的性状进行遗传参数的估计,研究表明,组合Ⅰ、组合Ⅱ控制开花的两对主基因的加性效应分别为7.174、5.189和3.474、1.402,显性效应分别为4.684、2.709和-1.172、-1.393,两个杂交组合的第一对主基因的显性程度高于第二对主基因的显性程度;两对主基因间的上位效应分别为-2.709和-0.884;两个杂交组合控制播种-开花的主基因的遗传率分别为63.007%和64.289%,多基因的遗传率分别为36.993%和35.702%,主基因的遗传率高于多基因的遗传率,组合Ⅰ、组合Ⅱ播种-开花性状主要是由主基因控制。表4
表2 四世代不同模型的AIC值
Table 2 The AIC values of different genetic models
模型Model组合ⅠCrossⅠ组合ⅡCrossⅡ播种-开花FT花铃期FBP全生育期WGP播种-开花FT花铃期FBP全生育期WGPA-12541651429633808593124283691291279693715931235613049A-225403613296147293150792725291884993715753235718037A-32608449230231035163305336914297034893741711736854744A-4260844923009725586326829541292796313738374836276733B-12408418528743833012909746094283177733642109134935093B-22474606729334704593079391357289814063670265135578B-324764436294283154331115070829146553680168935693772B-42474443629408308113071541992291263333724215135922246B-52573497830026171883279325684296302933721436336740776B-62571497630006145023277325439296102813731614736720764C-02389000528918173832990611572282649223642574535304683C-12387529336298833035037598289537993677078435300168D-02392172128910324712982646484283048733640831335272559D-12390639629354172363037727539289937743673254935266016D-22389506829356374513037053955289737873679076235320144D-32388639629334169923035728027289737043679086735320261D-42389502929356376953037052246289738653671254435246018E2320392828722500002899019043280258253631556234712878E-1235768292867644287289705835280809913627868934669348E-22385822529272849123006617432289062523663904334832434E-32383508529296523443031061279288715143671171634887578E-42381508529276523443029062256288809993669158734867473E-52383947529326196293045791504291380273703945835300642E-623798228292576635730285896288879273671088435240281
注:“__”表示候选模型
Note: “__”Means candidate model
表3 早熟性状遗传模型适合性检验
Table 3 Tests for goodness-of-fit about genetic models of earliness traits
组合Cross性状Traits模型Modle世代Generation模型适合性检验统计量 StatisticparameteroffitmodelU12U22U32nW2DnⅠ播种-开花EP10022(07001)0003(08073)37064∗(00136)00650244P20001(08522)0319(04524)192960∗(00002)01490253F10004(08052)0012(07425)0017(07197)00190170F20001(08591)0000(09984)0233(04869)13370159E-1P10087(05871)1145(03010)17098∗(00420)00560220P24919(01364)22269∗(00298)72812∗(00035)01970279F10650(03692)0237(04852)0221(04931)00410212F20011(07472)0239(04844)5760(01214)14220145花铃期EP10000(09757)0000(09716)0005(07946)00100136P20001(08768)0002(08327)4368(01483)00170171F10000(09682)0000(08911)0024(06941)00030118F20000(09016)0005(07867)0132(05463)01730099E-1P10000(09833)0000(08837)0065(06142)00110136P20003(08187)0000(09561)1508(02678)00140156F10000(09901)0000(09917)0000(09284)00020107F20000(09990)0001(08768)0152(05323)01540093全生育期EP10001(08503)0000(09915)0044(07418)001901705P20053(06304)0000(08895)2924(01910)003402108F10004(07981)0001(08506)9242(00812)005702335F20000(09915)0012(07418)2657(02017)031101052E-1P10445(04143)0186(05116)0083(05913)004201991P27102(01026)1540(02653)12131(00620)021402949F10458(04107)0013(07334)11069(00682)009502718F20015(07251)0000(09473)1452(02723)035001176Ⅱ播种-开花EP10001(08692)0001(08681)2870(01930)00190160P20250(04794)0000(09933)53582∗(00068)00870235F10000(08845)0089(05848)6938(01046)00280187F20000(08845)0003(08091)0162(05261)02620095E-1P10318(04527)0014(07327)5660(01229)00420207P28851(00846)0507(03988)119399∗(00009)03240298F10033(06702)0000(09908)8266(00900)00340201F20007(07731)0000(09908)1149(03006)03150111花铃期EP10000(09521)0000(08824)00167(07198)00080143P20067(06108)0677(03644)7535(00975)00320195F10001(08761)0068(06097)4281(01504)00170167F20000(09960)0000(09977)0000(09938)00080040E-1P10000(09801)0000(08837)0057(06253)00090148P20090(05840)0661(03671)4893(01370)00290193F10000(09083)0067(06109)6381(01120)00190164F20000(09083)0000(09901)0000(09583)00080039全生育期EP10282(04662)0008(07645)261178∗(00001)02140277P20000(09185)1385(02779)241927∗(00001)01580241F10002(08285)0116(05597)4982(01352)00360185F20000(09642)0000(09413)0000(09039)00370062E-1P10039(06563)0057(06250)183088∗(00002)01460252P20163(05249)5939(01185)204404∗(00002)01670229F10002(08381)0003(08213)8248(00901)00370191F20000(08814)0001(08716)0000(09457)00410067
注:*:差异显著(P<0.05);U12、U22、U32:均匀性检验;nW2:Smirnov检验;Dn:Kolmogorov 检验
Note:*: Significant difference at 0.05 levels;U12,U22andU32: were statistics of uniformity test;nW2was the statistic of Smirnov test;Dnwas the statistic of Kolmogorov test
2.3.2 花铃期主-多基因的遗传分析
通过最优遗传模型E-1(两对加性-显性-上位性主基因+加性-显性多基因混合遗传模型)对花铃期这一性状进行遗传参数估计,结果可以看出,组合Ⅰ、组合Ⅱ控制花铃期的两对主基因的加性效应分别为3.671、0.511和7.473、2.613;显性效应分别为-0.121、-4.480和-4.149、-4.658,两个杂交组合控制花铃期主基因的显性效应均为负值,控制花铃期性状的两对主基因呈负向不完全显性;组合Ⅰ控制花铃期的多基因的加性效应和显性效应分别为2.318、3.535,组合Ⅱ控制花铃期的多基因的加性效应和显性效应分别为-6.003、7.168;两个杂交组合控制花铃期主基因的遗传率分别为67.675%、86.405%;多基因的遗传率分别为32.325%、13.595%,主基因遗传率相比多基因遗传率要高,花铃期性状主要是由主基因来控制。表4
2.3.3 全生育期主-多基因的遗传分析
通过最优遗传模型E-1(两对加性-显性-上位性主基因+加性-显性多基因混合遗传模型)对全生育期性状进行遗传参数估计,研究表明,组合Ⅰ、组合Ⅱ控制全生育期的两对主基因的加性效应分别为6.365、3.513和8.460、4.450;显性效应分别为-0.283、-4.091和-1.000、1.298;组合Ⅰ、组合Ⅱ的两对主基因间均存在上位性效应,上位性效应分别为3.128和0.932;组合Ⅰ多基因的加性效应和显性效应分别为2.406、1.687,组合Ⅱ多基因的加性效应和显性效应分别为-5.109、-1.227;两个杂交组合主基因的遗传率分别为60.875%和70.837%;多基因的遗传率分别为39.125%和29.163%,主基因遗传率较高,全生育期性状是由主基因控制。表4
表4 棉花早熟性状遗传参数估计
Table 4 The estimates of genetic parameters of fit model of earliness characters
参数Parameter组合ⅠCrossⅠ组合ⅡCrossⅡ播种-开花FTE花铃期FBPE-1全生育期WGPE-1播种-开花FTE花铃期FBPE-1全生育期WGPE-1m(m1)54045470181052685837451275111679m25084055909m36720660694m45307259979da717436716365347474738460db518905113513140226134450ha4684-0121-0283-1172-4149-1000hb2709-4480-4091-1393-46581298i-270932713128-088427920932jab-51900726230108891255-2219jba-518-0507-3508-013970889-0507l-26994195347913965970006[d]23182406-6003-5109[h]353516877168-1227σp24590517659214122014819546042σmg2289235032560978454164332615σpg216981673336054356655213427hmg2(%)630076767560875642988640570837hpg2(%)369933232539125357021359529163
注:m、m1、m2、m3、m4:中亲值;da、ha:主基因a的加性效应和显性效应;db、hb:主基因b的加性效应和显性效应:[d]、[h]:多基因加性效应和显性效应;i、l:两个主基因的加×加效应和显×显效应;jab:加性(a)×显性(b)效应;jba:加性(b)×显性(a)效应;σp2:表型方差;σmg2:主基因方差;σpg2:多基因方差;hmg2:主基因遗传率;hpg2:多基因遗传率性
Note:m,m1,m2,m3,m4:Mid-parent value;da、ha: Additive effect and dominant effect of the first pair major gene;db、hb: Additive effect and dominant effect of the second pair major gene; [d]、[h]: Additive effective value and dominant effective value of polygene;i、l: Additive effect plus additive effect and dominant effect plus dominant effect of the two major genes;jab: Additive effect plus dominant effect of the two major genes;jba: Dominant effect plus additive effect of the two major genes;σp2: Phenotypic variance;σmg2: Major gene variance;σpg2: Polygenic variance;hmg2: Major gene heritability;hpg2: Polygene heritability
3 讨 论
棉花的早熟性是复杂的数量性状极易受到环境因素干扰[12-13],研究利用两个杂交组合的四个世代群体对棉花生育期的三个性状进行的联合分析,可以使试验的精准度有所增加,也可以使主基因遗传参数方面估计值的精度有所提高,确保了分析结果的准确度和可靠性[14-15]。在前人的研究结果中总结发现,数量性状是一种混合遗传模型,是由主基因和多基因共同来控制的,因此,控制数量性状的基因在其遗传效应大小上存在较大的差异[16]。棉花的早熟性是由多基因控制的复杂的数量性状,同时存在加性效应与显性效应[17-19]。播种-开花、花铃期、全生育期等性状是短季棉早熟的重要指标性状,Godoy[20]和吴吉祥等[21]的研究中指出生育期、始花期、铃期和果枝始节都是以加性效应为主,而显性效应则在次之;在研究中,组合Ⅰ各性状第一对主基因的加性效应分别为4.174、3.671和6.365,显性效应分别为4.684、-0.121和-0.283,各性状第一对主基因的加性效应均大于其显性效应;第二对主基因的加性效应分别为5.189、0.511、3.513,显性效应分别为2.709、-4.480、-4.091,加性效应大于显性效应。组合Ⅱ各性状的第一对主基因和第二对主基因的加性效应分别为3.474、7.473、8.460和1.402、2.613、4.450,显性效应分别为-1.172、-4.149、-1.000和-1.393、-4.658、1.298,加性效应大于显性效应,可见两组合各时期主基因的加性效应均大于显性效应,结果与其相符,且董娜[7]的研究结果也是如此。而周有耀等[22]则认为早熟性状的遗传应为显性效应;范术丽[8]在对播种-开花,全生育期和开花-吐絮研究结果的数据中表示,播种-开花和全生育期的加性效应分别为-6.257和-9.697 2,显性效应分别为-3.451 0和-6.527 58,加性效应小于显性效应,开花-吐絮的第一对主基因的加性效应为-12.443,显性效应为-11.751,加性效应小于显性效应;第二对主基因的加性效应和显性效应分别为12.488和-6.474,加性效应大于显性效应,两人结果基本一致。董承光[15],宋美珍等[23]均认为这些结果与研究的遗传材料、环境条件及当时年份有关。研究结果中花铃期的遗传模型和范术丽[8]的研究结果相符均为E-1模型,且主基因遗传率均大于多基因遗传率,说明花铃期是以主基因控制的可以在多环境下稳定遗传的性状,可以作为短季棉育种选择的重要标志。
4 结 论
研究以生育期差异显著的四个陆地棉品种为亲本,联合其构建的F1、F2为材料,利用多世代混合遗传分析的方法,明确了两个杂交组合播种-开花性状的遗传同时符合E模型,花铃期和全生育期性状均符合E-1模型,播种-开花,花铃期,全生育期三个性状可以稳定遗传,且都受两对主基因和多基因控制。组合Ⅰ播种-开花、花铃期、全生育期的主基因的遗传率分别为63.007%、67.675%、60.875%,多基因的遗传率分别为36.993%、32.325%、39.125%;组合Ⅱ播种-开花、花铃期、全生育期的主基因遗传率分别为64.298%、86.405%、70.837%,多基因遗传率分别为35.702%、13.595%、29.163%,数据显示,两个杂交组合控制生育期性状的主基因的遗传力要高于多基因的遗传率,两个杂交组合的三个生育期性状主要由主基因控制,同时还受到微效多基因的调控。联合两个杂交组合的表型数据来看,组合Ⅰ分离后代的三个表型数据都偏向早熟亲本,组合Ⅱ分离后代中除F1的花铃期和F2的播种-开花偏向晚熟,其他分离后代的三个表型数据均偏向早熟,两个杂交组合的表型总体偏向早熟;且两个杂交组合播种-开花、花铃期、全生育期的主基因的加性效应均要高于显性效应。遗传力高,加性效应大的生育期性状可以做为选育早熟棉品种的重要指标[20],并且,存在主基因遗传的生育期性状可以作为数量性状分子标记定位的主攻方向。
References)
[1] 喻树迅,张雷,冯文娟.快乐植棉-中国棉花生产的发展方向[J].棉花学报,2015,(3):283-290.
YU Shu-xun, ZHANG Lei, FENG Wen-juan. (2015). Easy and Enjoyable Cotton Cultivation: Developments in China's Cotton Production [J].CottonScience, (3):283-290.(in Chinese)
[2] 喻树迅,张雷,冯文娟.棉花生产规模化、机械化、信息化、智能化和社会服务化发展战略研究[J].中国工程科学,2016,(1):137-148.
YU Shu-xun, ZHANG Lei, FENG Wen-juan. (2016). Study on Strategy of Large Scale, Mechanization, Informationization, Intelligence and Social Services for Cotton Production [J].EngineeringSciences, (1):137-148. (in Chinese)
[3] 喻树迅.我国棉花生产现状与发展趋势[J].中国工程科学,2013,(4):9-13.
YU Shu-xun. (2013). Present situation and development trend of cotton production in China [J].EngineeringSciences, 15(4): 9-13. (in Chinese)
[4] 杨伟华,王延琴,周大云,等.植物修复重金属污染土壤研究进展[J].湖南农业科学,2014,(18):41-44.
YANG Wei-hua, WANG Yan-qin, ZHOU Da-yun, et al. (2014). Research Progress in Remediation of Heavy-Metal Contaminated Soils by Planting Cotton [J].HunanAgriculturalSciences, (18):41-44. (in Chinese)
[5] 喻树迅,范术丽.我国棉花遗传育种进展与展望[J].棉花学报,2003,15(2):120-124.
YU Shu-xun, FAN Shu-li. (2003). The Evolutions and Prospect of Cotton Genetics and Breeding in China [J].CottonScience, 15(2):120-124. (in Chinese)
[6] 袁有禄,张天真,郭旺珍,等.棉花高品质纤维性状的主基因与多基因遗传分析[J].遗传学报,2002,29(9):827-834.
YUAN You-lu, ZHANG Tian-zhen, GUO Wang-zhen, et al. (2002). Major-polygene effect analysis of super quality fiber properties in upland cotton (GossypiumhirsutumL.)[J].ActaGeneticaSinica, 29(9): 827-834. (in Chinese)
[7] 董娜.短季棉早熟及相关性状的混合遗传与QTL定位研究[D].新乡:河南科技学院硕士论文,2010.
DONG Na. (2010).MixedInheritanceandQTLLocationStudyonEarlinessanditsReiatedTraitsofShortSeasonCotton[D]. Master Dissertation. Henan Institute of Science and Technology, Xinxiang. (in Chinese)
[8] 范术丽.短季棉早熟性相关性状的遗传及QTL 定位研究[D].北京:中国农业科学院博士论文, 2004.
FAN Shu-li. (2004).StudyoninheritanceofearlinessanditsrelativetraitsofshortseasoncottonandQTLsmapping[D].PhD Dissertation. The Chinese Academy of Agricultural Sciences, Beijing. (in Chinese)
[9] 喻树迅,黄祯茂.短季棉品种早熟性构成因素的遗传分析[J].中国农业科学,1990,23(6):48-54.
YU Shu-xun, HUANG Zhen-mao.(1990). Inheritance analysis on earliness components of short season cotton varieties in G.hirsutum[J].ScientiaAgriculturaSinica, 23(6): 48-54. (in Chinese)
[10] 艾先涛,李雪源,王俊铎,等.北疆陆地棉育成品种表型性状遗传多样性分析[J].分子植物育种,2011,9(1):113-122.
Ai Xian-tao, Li Xue-yuan, Wang Jun-duo, et al. Genetic Diversity on Agronomic Phenotypes in Upland Cotton Varieties of North Xinjiang Area[J].MolecularPlantBreeding, 2011,9(1):113-122. (in Chinese)
[11] 章元明,盖均镒,张孟臣.利用P1、F1、P2和F2或F2∶3世代联合的数量性状分离分析[J].西南农业大学学报,2000,22(1):6-9.
ZHANG Yuan-ming, GAI Jun-yi, ZHANG Meng-chen. (2000). Jointly segregating analysis of P1P2F1and F2or F2∶3families [J].JournalofSouthwestAgriculturalUniversity, 22(1): 6-9. (in Chinese)
[12] 林忠旭,冯常辉,郭小平,等. 陆地棉产量、纤维品质相关性状主效 QTL 和上位性互作分析[J]. 中国农业科学,2009,42(9):3 036-3 047.
Lin Zhong-xu, Feng Chang-hui, Guo Xiao-ping, et al. (2009). Genetic Analysis of Major QTLs and Epistasis Interaction for Yield and Fiber Quality in Upland Cotton[J].ScientiaAgriculturaSinica, 42(9):3,036-3,047. (in Chinese)
[13] 柳宾.棉花早熟性、产量和纤维品质性状的遗传分析和QTL定位[D]. 山东:山东农业大学,2010.
Liu Bin. (2010).GeneticAnalysisandQTLMappingforMaturity,YieldandFiberQualityTraitsofCotton[D]. Shandong: Shandong Agricultural University.(in Chinese)
[14] 殷剑美,武耀廷,朱协飞.陆地棉产量与品质性状的主基因与多基因遗传分析[J].棉花学报,2003,15(2): 67-72.
YIN Jian-mei, WU Yao-ting, ZHU Xie-fei, et al. (2003). Genetic analysis of yield traits and fiber qualities by using major gene plus polygene mixed inheritance model in upland cotton (G.hirsutumL.)[J].CottonScience, 15(2):67-72. (in Chinese)
[15] 董承光,王娟,周小凤,等.新疆早熟陆地棉早熟性状的遗传分析[J].西北农业学报,2014,23(12):96-101.
DONG Cheng-guang, WANG Juan, ZHOU Xiao-feng, et al. (2014). Inheritance of earliness traits in Xinjiang early maturityupland cotton (GossypiumhirsutumL.)[J].ActaAgriculturaeBoreali-OccidentalisSinica, 23(12):96-101. (in Chinese)
[16] 杜雄明,汪若海,刘国强,等.棉花纤维相关性状的主基因-多基因混合遗传分析[J].棉花学报,1999,11(2):73-78.
DU Xiong-ming, WANG Ruo-hai, LIU Guo-qiang, et al. (1999). Inheritance Analysis of the Characters Related with Fiber by Using Mixed Major Gene and Polygene Model [J].CottonScience, 11(2):73-78. (in Chinese)
[17] 范术丽,喻树迅,原日红,等.短季棉早熟性的遗传效应及其与环境互作研究[J].西北植物学报,2006,26(11):2 270-2 275.
FAN Shu-li, YU Shu-xun, YUAN Ri-hong, et al. (2006). Genetic effects and environmental interactions of early maturity in short-season cotton [J].ActaBotanicaBoreali-OccidentaliaSinica, 26(11): 2,270-2,275. (in Chinese)
[18] 詹有俊,杨涛,孙建船,等.特早熟陆地棉的遗传效应及杂种优势分析[J].农业现代化研究,2012,33(4):943-947.
Zhan You-jun, Yang Tao, Sun Jian-chuan, et al. Analysis of Genetic Effect and Heterosis of Special-early MatureGossypiumhirsutumL. [J].Researchofagriculturalmodernization, 2012,33(4):943-947. (in Chinese)
[19] 詹有俊,杨涛,孙建船,等.特早熟陆地棉熟性产量品质的遗传相关分析[J].农业现代化研究,2013,34(1):118-121.Zhan You-jun, Yang Tao, Sun Jian-chuan, et al. (2013). Genetic Correlation Analysis of Early Maturity Yield and Quality in Special-early MatureGossypiumhirsutumL. [J].Researchofagriculturalmodernization, 34(1):118-121. (in Chinese)
[20] Godoy, A. S., & Palomo, G. A. (1999). Genetic analysis of earliness in upland cotton (Gossypiumhirsutuml.). ii. yield and lint percentage.Euphytica, 105(2):161-166.
[21] 吴吉祥,朱军,季道藩,等.陆地棉产量性状的遗传效应及其与环境互作的分析[J].遗传,1995,17(5):1-4.
WU Ji-xiang, ZHU Jun, JI Dao-fan, et al. (1995). Analysis of Genetic Effect × Environment Interactions for Yield Traits in Upland Cotton [J].Hereditas, 17(5):1-4. (in Chinese)
[22] 周有耀.棉花早熟性与纤维品质性状关系的研究[J].中国棉花,1990,17(5):13-15.
ZHOU You-yao. (1990). Study on the relationship between cotton maturity and fiber quality traits [J].ChinaCotton, 17(5):13-15. (in Chinese)
[23] 宋美珍,喻树迅,范术丽,等.短季棉主要农艺性状的遗传分析[J].棉花学报,2005,17(2):94-98.
SONG Mei-zhen, YU Shu-xun, FAN Shu-li, et al. (2005). Genetic analysis of main agronomic traits in short-season upland cotton (Gossypium hirsutum L.) [J].CottonScience, 17(2): 94-98. (in Chinese)
[24] 艾尼江,朱新霞,管荣展,等.棉花生育期的主位点组遗传分析[J].中国农业科学,2010,43(20):4 140-4 148.
AI Ni-jiang, ZHU Xin-xia, GUAN Rong-zhan, et al. (2010). Genetic Analysis of Major Locus Group Constitutions of Growth Stages in Upland Cotton [J].ScientiaAgriculturaSinica, 43(20):4,140-4,148. (in Chinese)
Supported by:The National Key Technology R&D Program "The project of drought resistant, saline-alkaline tolerant cotton cultivar screening and breeding " (2014BAD03B01)
Genetic Analysis of Growth Traits in Short Season Cotton
TIAN feng1, 2,FAN Shu-li2,WEI Heng-ling2,WANG Han-tao2,ZHAO Shu-qi2,3,PANG Chao-you2,HU Shou-lin1,YU Shu-xun2
(1.CollegeofPlantScience,TarimUniversity,AlarXinjiang843300,China; 2.InstituteofCottonResearchofCAAS/StateKeyLaboratoryofCottonBiology,AnyangHenan455000,China;3.HuanggangAcademyofAgriculturalSciences,HuanggangHubei438000,China)
【Objective】 To study the genetic law of short-season cotton growth period.【Method】Our study utilized four upland cotton cultivars to construct two crosses and analyzed three traits (Flowering time FT, Flowering and boll-setting period FBP, Whole growing period WGP) of growth period using the joint segregation analysis method of mixed major gene plus polygene inheritance model.【Result】The results from phenotypic analysis showed that: the other phenotypic traits were in favor of the early-maturing parent and the partial early-maturing parent, except for the combination of F1in the blooming stage and F2sowing-flowering stage. The genetic analysis results showed that: the same traits of the combination I and II were in accordance with the same genetic model, and were controlled by two pairs of major genes.【Conclusion】It can be seen from the heritability of major genes and polygenes, the three phenotypic traits were mainly controlled by the major genes and the minor genes, the heritability of the three phenotypic traits is high, and the additive effect is large. This study could provide a theoretical basis for QTL mapping and molecular marker assisted breeding of cotton early maturing traits.
short season cultivars; growth stages; major gene plus polygene inheritance; genetic analysis
2016-11-07
国家科技支撑计划“抗旱、耐盐碱棉花品种筛选及新品种培育”(2014BAD03B01)
田锋(1990-),男,新疆阿克苏人,硕士研究生,研究方向为棉花遗传育种,(E-mail)tian_feng12@sina.com
庞朝友(1979-),男,河北邢台人,副研究员,研究方向为棉花遗传育种,(E-mail)chypang@163.com 胡守林(1968-),男,新疆阿克苏人,教授,研究方向为棉花育种,(E-mail)179390139@qq.com
S562.1;S503.2
A
1001-4330(2017)02-0197-09
猜你喜欢
临床肝胆病杂志(2020年1期)2020-12-20
支部建设(2019年24期)2019-09-28
金桥(2019年12期)2019-08-13
文苑(2019年14期)2019-08-09
消费导刊(2017年24期)2018-01-31
现代农业科技(2016年16期)2017-07-19
中国稻米(2017年2期)2017-04-28
安徽农学通报(2017年1期)2017-02-15
中国科技纵横(2016年15期)2016-12-29
中学语文(2015年27期)2015-03-01
