
靶向番茄SlACS2基因CRISPR-Cas9 sgRNA的设计和分析
2017-04-10 06:10白云凤张爱萍闫建俊贺飞燕张维锋冯瑞云刘江娜张西英
生物信息学 2017年1期
白云凤,张爱萍,闫建俊,贺飞燕,张维锋*,冯瑞云,刘江娜,张西英
(1.农业部黄土高原作物基因资源与种质创制重点实验室,作物遗传与分子改良山西省重点实验室(山西省农业科学院作物科学研究所),太原 030031;2. 新疆生产建设兵团第6师农业科学研究所,新疆 五家渠831300;3.山西大学生物工程学院,太原030006)
靶向番茄SlACS2基因CRISPR-Cas9 sgRNA的设计和分析
白云凤1,张爱萍2,闫建俊1,贺飞燕3,张维锋1*,冯瑞云1,刘江娜2,张西英2
(1.农业部黄土高原作物基因资源与种质创制重点实验室,作物遗传与分子改良山西省重点实验室(山西省农业科学院作物科学研究所),太原 030031;2. 新疆生产建设兵团第6师农业科学研究所,新疆 五家渠831300;3.山西大学生物工程学院,太原030006)
番茄为呼吸跃变型果实,伴随呼吸跃变产生大量乙烯,即系统II乙烯,易使番茄果实过熟,导致腐烂变质。SlACS2是番茄系统II乙烯合成的限速酶,通过CRISPR-Cas9基因组编辑系统修饰该基因,调控系统II乙烯过量表达,将迟滞番茄过熟。本研究基于RNA-seq建立了SlACS2基因的数字表达谱,表明该基因呈果实特异性表达,在植株的根、茎、叶等部位不表达。SlACS2位于番茄1号染色体,含4个外显子和3个内含子。利用在线工具CRISPRdirect 和CRISPR-P发现第1、2、3外显子分别具有18、9和11条sgRNA。其中,sgRNA1-14和sgRNA3-8及二者的近PAM的12 nt 种子序列在番茄基因组是唯一序列,GC含量高于40%,不存在TTTT终止序列。BLAST结果表明,sgRNA1-14和sgRNA3-8与GenBank公布的8条SlACS2同源序列高度一致,位于该基因的保守区,而与SlACS4和SlACS6的同源序列存在多个SNP,预示这2条sgRNA可用于番茄不同品种SlACS2基因的靶向编辑,并可规避对SlACS家族其他同源基因的脱靶效应。
番茄;ACC合成酶2;sgRNA;CRISPR-Cas9;基因组编辑
FENG Ruiyun1, LIU Jiangna2, ZHANG Xiying2
乙烯(Ethylene)是植物内源激素,对植物生长发育具有重要调控作用。具有呼吸跃变的果实在呼吸跃变前产生基础水平的少量乙烯,被称为系统I乙烯,调控植物正常生长、发育和应对胁迫反应等。系统I乙烯具有自抑制机制。随着果实成熟,伴随呼吸跃变产生大量乙烯,被称为系统II乙烯。系统II乙烯具有自催化机制,促进花的凋亡和果实成熟[1]。
乙烯的生物合成遵循甲硫氨酸(Methionine,Met)→S-腺苷甲硫氨酸(S-Adenosyl methionine,SAM)→1-氨基环丙烷1-羧酸ACC(1-aminocyclopropane-1-carboxylic acid,ACC)→乙烯的途径。ACC是合成乙烯的直接前体,由ACC合成酶ACS(ACC synthase)催化SAM形成,这个过程被认为是乙烯合成的限速步骤,ACS是乙烯合成的限速酶。番茄(Solanumlycopersicum)是呼吸跃变型果实,其ACS(SlACS)由多基因家族编码,SlACS1A和SlACS6主导系统I乙烯的合成,负责番茄果实在呼吸跃变前产生基础水平的少量乙烯,SlACS2和SlACS4主导系统Ⅱ乙烯的合成[2],在果实成熟过程中伴随呼吸跃变产生大量乙烯,使得番茄成熟过程难以控制,果实易过熟导致腐烂变质。
通过基因工程手段抑制番茄SlACS2的表达,主要是把SlACS2的反义RNA基因[3-4]或RNAi基因[5-6]导入番茄,可以减缓番茄果实中的乙烯合成,迟滞果实过熟,延长货架期,提高耐贮运性能。上述利用常规转基因技术,导入的外源基因在植物基因组上的整合位置具有随机性,容易出现一些非预期效应,如转基因整合到宿主基因的“阅读框”,使其不能有效表达,影响基因工程育种效率。民众对于转基因的安全性疑虑,也增加了转基因作物评价和应用成本。
CRISPR-Cas9是一种新型的基因组定点编辑系统[7-8],其通过一段20 nt的sgRNA(Single guide RNA)与基因组同源序列互补配对引导Cas9蛋白结合于基因组的特异性位点,在sgRNA 3’端PAM(Protospacer adjacent motif, 前间区序列邻近基序)元件上游的第3~4 bp处剪切,造成DNA双链断裂(Doubled-strand breaks, DSB)。不同CRISPR-Cas9的PAM序列有所不同,经典酿脓链球菌(Streptpcpccuspyogenes)中的PAM元件序列为5’-NGG-3’。因此,CRISPR-Cas9系统中靶位点的选择是由20 nt 的sgRNA及3’端的PAM序列共同决定的。
Cas9蛋白切割造成的DNA双链断裂有两条修复途径[9],即非同源末端连接(Non-homologous end joining, NHEJ)和同源重组(Homologous recombination, HR)。非同源末端连接是主要修复途径,同源重组只发生在特定的细胞周期和类型中。非同源末端连接容易出错,修复过程中会引入短的缺失或插入,抑制或修饰特定基因功能。与诱发突变相比,CRISPR-Cas9系统为定点突变,效率高、可控性好。CRISPR-Cas9系统通过植物基因组原位编辑来抑制或修饰基因功能,无需整合新基因,比转基因技术更为安全可控,易于消除民众安全性疑虑。
CRISPR-Cas9系统中,20 nt的sgRNA及其3’端的PAM元件总长度仅23 nt,易在基因组的非目标区段出现同源序列。另外,Cas9核酸酶可能切割与sgRNA不完全匹配的序列,导致脱靶效应[10-11]。规避脱靶效应和提高编辑效率是应用CRISPR-Cas9系统的关键。本文设计、筛选出了番茄SlACS2基因的sgRNA,以期为利用CRISPR-Cas9技术抑制SlACS2基因表达、提高番茄的耐贮运性能提供依据。
1 材料与方法
1.1SIACS2数字表达谱的建立
以番茄功能基因组数据库网站(http://ted.bti.cornell.edu/cgi-bin/)中RNA-seq data的数据作基础,建立SlACS2基因的基于RNA-seq的数字表达谱。
1.2SlACS2染色体定位和基因组结构的确定
利用NCBI(http://www.ncbi.nlm.nih.gov/)核酸数据库查询获得SlACS2的cDNA,以此作query,搜索番茄基因组数据库得到相应的gDNA序列和所在染色体信息。通过序列比对确定SlACS2的基因组结构以及外显子所在染色体位置。
1.3 sgRNA的设计和选择
根据CRISPR-Cas9靶点设计原则,利用CRISPRdirect((https://crispr.dbcls.jp/)[12], 2015) 筛选SlACS2外显子区域的sgRNA序列,即3’端有PAM元件的20个连续的碱基序列,PAM元件设定为NGG[13],sgRNA的结构为5’-(N)20NGG-3’,N为任意核苷酸。利用CRISPR-P(http://cbi.hzau.edu.cn/crispr/)[14]对sgRNA的靶向特异性进行综合评分。
2 结果与分析
2.1基于RNA-seq的SlACS2数字表达谱分析
以番茄功能基因组RNA-seq数据库为基础,分析了SlACS2基因(TC125329) 在不同组织以及不同生长时期的的表达模式(见图1)。结果显示,SlACS2在番茄的种子、胚根、顶端分生组织、不同时期的幼苗、根系、叶片、花蕾、绿果中均无表达,在子房和愈伤组织中有低水平表达,在破色期的果实和成熟的红果实中有较高水平的表达。上述基于RNA-seq的表达谱分析证实了SlACS2是与果实成熟相关的基因,利用CRISPR-Cas9技术对该基因进行原位编辑,有可能减缓番茄内源乙烯的产生,迟滞番茄过熟,提高贮运品质。
2.2SlACS2染色体定位和基因组结构
从GenBank 搜索到SlACS2序列(GenBank Accession:NM_001247249.2),mRNA长度1 855 nt, cds位于181~1 629 nt之间,长度1 458 nt。
以SlACS2的cds作Querry,在番茄的基因组数据库进行Blast。比对结果表明,SlACS的gDNA位于番茄的1号染色体(GenBank Accession:NC_015438.2),位置在88 456 011~88 453 490 nt之间,长度2 522 nt,由4个外显子、3个内含子组成(见图2),前3个外显子均较短,位于SlACS2的5’端,可在每个外显子区域设计sgRNA,规避sgRNA分布在外显子和内含子连接处。

图1 基于RNA-seq的SlACS2表达谱Fig.1 The digital expression profile of SlACS2 based on RNA-sequence
注:1-子房; 2-半绿果实; 3-成熟绿果实; 4-破色期果实; 5-成熟红果实; 6-花蕾(0-3 mm); 7-花蕾(3-8 mm); 8-开花前花蕾(8 mm); 9-完全展开花; 10-发育阶段花的混合物; 11-野生番茄花粉; 12-感染假单胞菌叶片; 13-假单胞菌叶片; 14-混合诱导叶片; 15-植株的顶端分生组织(4-6周龄); 16-植株顶端分生组织(8周龄); 17-花前根; 18-挂果期根; 19-营养匮乏的根; 20-完全吸涨5 d后的胚根; 21-完全吸涨7 d后的幼苗; 22-休眠种子; 23-愈伤组织。
PRKM: 每百万reads中来自于特定基因每千碱基长度的reads数。

图2 SlACS2的基因组结构和染色体位置Fig.2 SlACS2 enomic costructure and its location on the chromosome
2.3SlACS2第1外显子sgRNA的设计和评价
SlACS2第1外显子长度171 nt,位于染色体88 456 011~88 455 841 nt之间。根据CRISPR-Cas9靶点设计原则,设定sgRNA长度为20 nt,下游的PAM元件为NGG。CRISPRdirect在线工具分析表明,第1外显子含有18条sgRNA(见表1)。CRISPR-P分析表明,这些sgRNA的靶向特异性综合得分在29~49。
在CRISPR-Cas9系统中,sgRNA的转录通常由U6或者U3启动子驱动,二者驱动转录的活性相对较高。U6或者U3启动子属于Pol III型启动子[15],连续的TTTT序列会使转录终止。在第1外显子的18条sgRNA中,有9条(sgRNA1-1、-4、-5、-6、-8、-9、-11、-12、-18)含有连续的TTTT序列,可排除选择。另外, sgRNA1-2的GC含量较低, sgRNA1-7除了在靶位点外,在番茄5号染色体上也有与之完全匹配的序列,易产生脱靶效应。在其余7条sgRNA(见图3)中,位于外显子132~154 nt的sgRNA1-14特异性最好,该sgRNA和它包含的对特异性起重要作用的、邻近PAM 的12 nt种子序列[16]在番茄基因组上均是唯一序列,其他部位没有与之完全匹配的序列。sgRNA1-14的GC含量45%,靶向特异性综合得分也最高,可作为优选sgRNA。
注:字体加粗表示该sgRNA具有高特异性;含有连续TTTT的靶向序列用灰色表示,含有polIII启动子的载体应避免sgRNA中含连续的TTTT序列。

图3 SlACS2第1外显子主要sgRNAs的分布。Fig.3 The position of some important sgRNAS of exon 1 in SlACS2
注:括号内数值为该sgRNA所在外显子位置。
Cas9蛋白包含氨基端的RuvC-like结构域及位于蛋白中间位置的HNH核酸酶结构域。HNH核酸酶结构域切割与sgRNA互补配对的模板链,RuvC-like结构域切割另1条链。突变Cas9核酸酶的一个功能结构域时,则只能切割DNA双链中的一条链,形成Cas9单切口酶(Nickase)。体内DNA单链断裂修复采用碱基切除修复途径,不能产生突变。当1对分别位于染色体正负链上、方向相反且距离较近的两个sgRNA(即“Paired-sgRNA”)同时引导Cas9单切口酶对基因组编辑时,两个近距离的单链断裂会组成双链断裂,通过非同源末端连接修复,造成短的缺失或插入,导致移码突变;而潜在的脱靶位点处只有单链断裂,修复过程不易发生突变。因此,采用“paired-sgRNA”和Cas9单切口酶编辑策略时,酶切识别所涉及的核苷酸数就由20 nt增加到40 nt,从而提高靶位点的专一性,降低脱靶风险[17-18]。据此,采用“paired-sgRNA”和Cas9单切口酶编辑策略,对第1外显子sgRNA1-3及其互补链的sgRNA1-14进行切割,间隔66 nt,有较大概率产生专一性移码突变。
2.4SlACS2第2、3外显子sgRNA的设计和分析
SlACS2第2外显子位于染色体86 455 742~86 455 614 nt之间,长129 nt,与第1外显子相隔98 nt的内含子。CRISPRdirect在线分析表明,第2外显子有9条sgRNA序列,其GC含量均低于35%,邻近PAM 12 nt 的种子序列的脱靶位点在2个以上(见表2),没有合适的sgRNA可供选择。
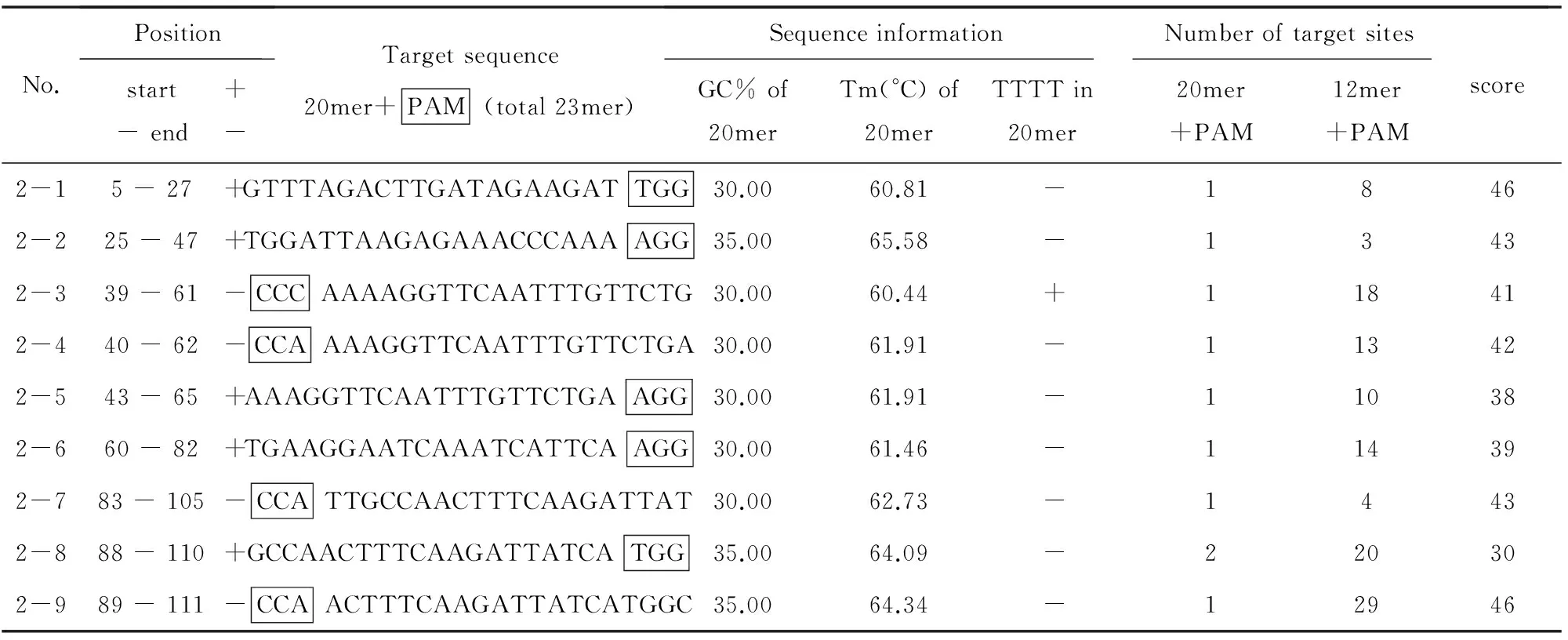
表2 SlCAS2基因第2外显子含有sgRNAsTable 2 sgRNAs of exon 2 in SlCAS2 gene
SlACS2第3外显子位于染色体86 455 528~86 455 368 nt之间,长161 nt,与第2外显子相隔85 nt的内含子,与第4外显子相隔881 nt的内含子。CRISPRdirect在线分析表明,第3外显子有11条sgRNAs。CRISPR-P分析表明,sgRNAs的靶向特异性综合得分在22~48,差异较大(见表3)。在这些sgRNAs中,sgRNA3-9和sgRNA3-10含有连续的TTTT序列,易使转录提前终止;sgRNA3-1、-2、-3的GC含量较低;sgRNA3-6的靶向特异性综合得分最低。在其余5条sgRNAs(见图4)中,sgRNA3-8特异性最好,其邻近PAM 12 nt 的种子序列在番茄全基因组上是唯一序列,其他部位没有与之完全匹配的序列,靶向特异性综合得分也较高。该sgRNA 3’端第20位的碱基为鸟嘌呤G,意味着基因组编辑效率较高[19],可作为优选序列。sgRNA3-11特异性也较好,靶向特异性综合得分也较高,其12 nt 的种子序列除了靶位点外,在5号染色体的1个位点上有与其完全匹配的序列,其脱靶效应可根据脱靶位点两侧的序列(见图5)设计引物,对PCR产物测序予以验证。另外该sgRNA的第一碱基为G,也适合U6启动子转录,是另一优选序列。sgRNA3-8与位于互补链的sgRNA3-4间隔5 nt,sgRNA3-11与位于互补链的sgRNA3-7之间相隔34 nt,这两对sgRNA均可设计为“paired-sgRNA”,用Cas9单切口酶进行编辑。
2.5 优选sgRNA的SNP分析
以SlACS2的第1和第3外显子作Query在GenBank中进行BLAST,搜索同源序列,进行多序列比对(见图6, 图7)。比对结果表明, sgRNA1-14、sgRNA3-11在搜索到的8条序列上均完全一致,sgRNA3-8也只有1个SNP,表明本研究优选的3条sgRNA均具有较好的保守性,可用于不同品种SlACS2基因的靶向编辑。而与GenBank公布的SlACS4和SlACS6比对,sgRNA1-14均有5个SNP,其中3个位于种子序列区;sgRNA3-8分别有3个和6个SNP,且有1个SNP位于PAM的第2个核苷酸,TGG突变为TAG;sgRNA3-11有3个SNP,其中2个位于种子序列区;预示本研究优选的3条sgRNA均可规避对SlACS家族其他成员的脱靶效应。
注:字体加粗示该sgRNA具有高特异性;含有连续TTTT的靶向序列用灰色表示,含有polIII启动子的载体应避免sgRNA中含连续的TTTT序列。

图4 SlACS2第3外显子主要sgRNAs的分布Fig.4 The position of some important sgRNAS of exon 3 in SlACS 2
注:括号内数值为该sgRNA所在外显子位置。
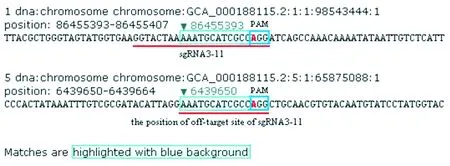
图5 第3外显子sgRNA3-11的脱靶位点和染色体位置Fig.5 The position of off-target site of sgRNA3-11 in exon 3

图6 SlACS2第1外显子sgRNA1-14的SNP位点Fig.6 SNP loci on sgRNA1-14 of exon 1 in SlACS2
3 讨论
科学设计sgRNA,提高基因组编辑效率和最大限度降低脱靶风险是CRISPR/Cas9 系统面临的最大挑战。番茄是模式植物之一,已完成了全基因组测序,sgRNA的特异性及其脱靶位点可通过全基因组层面扫描获得相关信息。sgRNA的设计已有多个工具软件和在线分析平台,如CRISPRdirect综合考虑了sgRNA全长和12 nt及8 nt种子序列的特异性,结果输出中会标出每条sgRNA在外显子上的位置和GC含量,还将特异性高、脱靶位点少的sgRNA以及含有转录终止序列“TTTT”的 sgRNA 分别用亮绿色和灰色作出标示;华中农业大学作物遗传改良国家重点实验室开发的CRISPR-P则根据特定公式对每个sgRNA序列的靶向特异性进行综合评分和排序,并标示出sgRNA靶标位点中是否含限制性内切酶识别序列,以方便采用酶切法检测基因组切割效率。本研究综合运用CRISPRdirect和CRISPR-P两个在线分析平台,对SlACS2基因5’端的第1、2、3个外显子上所分布的sgRNAs进行了综合评价,筛选出3条sgRNAs优选序列,即第1外显子sgRNA1-14、第3外显子sgRNA3-8和sgRNA3-11。其中sgRNA1-14和sgRNA3-8的全序列或12 nt的种子序列在番茄基因组上均为唯一序列,但它们的5’端第1个碱基为A或C。双子叶植物sgRNA 的转录一般选用U6启动子驱动,其转录活性相对较高,有明确的转录起始位点,即以“G”碱基开头。精确起始转录能消除无关 DNA 序列的转录,从而减少脱靶效应的产生[20-21]。因此,如果选用U6启动子,需要在这2条sgRNA的5’端添加鸟嘌呤核苷酸。有研究表明,在sgRNA的5’端额外增加两个鸟嘌呤核苷酸后能够显著提高CRISPR/Cas9 系统的特异性[17],为非G起始的sgRNA应用提供了依据。
乙烯除了促进果实成熟,在植物种子萌发、叶片扩展、根毛伸长、侧根生长、开花、植株衰老等阶段均起重要作用。抑制番茄系统I或过度抑制系统II乙烯的生成,将影响番茄的正常生长或致使果实不能成熟。本研究从SlACS2中筛选出的3条sgRNAs与GenBank中公布的SlACS4和SlACS6的同源序列存在3~8个SNP,其中2~3个SNP分布在12 nt的种子序列区。这些SNP的存在规避了sgRNA对SlACS4和SlACS6的脱靶效应,可望达到预期目的:(1)既抑制系统II乙烯合成限速酶基因SlACS2的表达,又不影响SlACS4的正常表达,使系统II乙烯保持在一定水平而不致过量,以迟滞番茄过熟;(2)不干扰系统I乙烯形成的SlACS6基因的正常表达,保证番茄的正常生长。生物信息学可以提供许多有用信息,但不能代替实验验证。在应用这3条优选的sgRNA改良番茄的耐贮运性状前,可先用原生质体系统验证其靶向能力和编辑效率。
CRISPR-Cas9系统通过sgRNA引导Cas9蛋白在PAM元件上游第3~4 bp处剪切,造成DNA双链断裂,在非同源末端连接修复过程中引入短的缺失或插入,使基因失活,但失活基因仍能复制和转录,消耗植物营养和能量,如能删除失活基因的完整序列或大片断,将降低植物营养和能量的消耗。近年来,已通过双sgRNA引导Cas9系统在植物[22-23]和动物[24]上删除了一段DNA序列,最大缺失片段达105 kb。本研究筛选的sgRNA1-14和sgRNA3-8特异性较好,在染色体上相隔409 nt,利用双sgRNA引导Cas9将其删除,造成DNA大片段缺失,将更彻底的抑制SlACS2基因功能。
References)
[1]MCMURCHIE E, MCGLASSON W, EAKS I. Treatment of fruit with propylene gives information about the biogenesis of ethylene[J]. Nature, 1972, 237(5352): 235-236. DOI:10.1038/237235a0.
[2]BARRY C, LOP-TOUS M, GRIERSON D. The regulation of 1-aminocyclopropane-1-carboxylic acid synthase gene expression during the transition from system-1 to system-2 ethylene synthesis in tomato [J]. Plant Physiology, 2000, 123(3): 979-986. DOI: 10.1104/pp.123.3.979.
[3]OELLER P, LU M, TAYLOR L, et al. Reversible inhibition of tomato fruit senescence by antisense RNA[J]. Science, 1991, 254(5030): 437-439. DOI: 10.1126/science.1925603.
[4]熊爱生,姚泉洪,李贤,等. ACC氧化酶和ACC合成酶反义RNA融合基因导入番茄和乙烯合成的抑制[J]. 实验生物学报,2003,36(6):35-41.
XIONG Aisheng, YAO Quanhong, LI Xian, et al. Double antisense ACC oxidase and ACC synthase fusion gene introduced into tomato by agrobacterium-mediated transformation and analysis the ethylene production of transgenic plants[J]. Acta Biologiae Experimentalis Sinica, 2003,36(6): 35-41.
[5]陈银华,李汉霞 ,叶志彪. 不同结构的外源ACO基因导入番茄对乙烯生成速率的影响[J]. 园艺学报,2007,34(3):644-648.
CHEN Yinhuan, LI Hanxia, YE Zhibiao. Effects of introduction of different T-DNA structures of ACO gene to tomato genome on ethylene production rate[J]. Acta Horticulturae Sinica, 2007, 34(3): 644-648 .
[6]GUPTA A, PAL R, RAJAM M. Delayed ripening and improved fruit processing quality in tomato by RNAi-mediated silencing of three homologs of 1-aminopropane-1-carboxylate synthase gene[J]. Journal of Plant Physiology, 2013, 170(11): 987-995. DOI: 10.1016/j.jplph.2013.02.003.
[7]CONG L, RAN F, COX D, et al. Multiplex genome engineering using CRISPR/Cas systems[J]. Science, 2013, 339(6121): 819-823. DOI: 10.1126/science.1231143.
[8]Doudna J A, Charpentier E. The new frontier of genome engineering with CRISPR-Cas9[J].Science,2014,346(6213):1258096-1258096.DOI:10.1126/science.1258096.
[9]ILIAKIS G, WANG H, PERRAULT A, et al. Mechanisms of DNA double strand break repair and chromosome aberration formation[J]. Cytogenetic & Genome Research, 2004, 104(1-4): 14-20. DOI: 10.1159/000077461.
[10]FU Y, FODEN J, KHAYTER C,et al. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells[J]. Nature Biotechnology, 2013, 31(9): 822-826. DOI:10.1038/nbt.2623.
[11]HSU P, SCOTT D, WEINSTEIN J, et al. DNA targeting specificity of RNA-guided Cas9 nucleases[J]. Nature Biotechnology, 2013, 31(9): 827-832. DOI:10.1038/nbt.2647.
[12]NATIO Y, HINO K, BONO H, et al. CRISPRdirect: software for designing CRISPR/Cas guide RNA with reduced off-target sites[J]. Bioinformatics, 2015, 31(7): 1 120-1 123. DOI:10.1093/bioinformatics/btu743.
[13]ZHANG Y, GE X, YANG F, et al. Comparison of non-canonical PAMs for CRISPR/Cas9-mediated DNA cleavage in human cells[J]. Scientific Reports, 2014(4): 5405. DOI: 10.1038/srep05405.
[14]LEI Y, LU L, LIU H, et al. CRISPR-P: a web tool for synthetic single-guide RNA design of CRISPR-system in plants[J]. Molecular Plant, 2014, 7(9): 1 494-1 496. DOI:10.1093/mp/ssu044.
[15]SHAN Q, WANG Y, LI J, et al. Targeted genome modification of crop plants using a CRISPR-Cas system[J]. Nature Biotechnology, 2013, 31(8): 686-688. DOI:10.1038/nbt.2650 .
[16]PATTANAYAK V, LIN S, GUILINGER J, et al. High-throughput profiling of off-target DNA cleavage reveals RNA-programmed Cas9 nuclease specificity[J]. Nature Biotechnology, 2013, 31(9): 839-843. DOI: 10.1038/nbt.2673.
[17]CHO S W, KIM S, KIM Y, et al. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases [J]. Genome Research, 2014, 24(1): 132-141. DOI: 10.1101/gr.162339.113.
[18]RAN F, HSU P, LIU C, et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity[J]. Cell, 2013, 154(6): 1 380-1 389. DOI: 10.1016/j.cell.2013.08.021.
[19]DOENCH J, HARTENIAN E, GRAHAM D, et al. Rational design of highly active sgRNAs for CRISPRCas9-mediated gene inactivation [J]. Nature Biotechnology, 2014, 32(12): 1262-1267.
[20]BELHAJ K, CHAPARRO-GARCIA A, KAMOUN S, et al. Editing plant genomes with CRISPR/Cas9 [J]. Current Opinion in Biotechnology, 2015(33): 76-84. DOI: 10.1016/j.copbio.2014.11.007.
[21]LI X, JIANG D H, YONG K L, et al. Varied transcriptional efficiencies of multiple Arabidopsis U6 small nuclear RNA genes[J]. Journal of Integrative Plant Biology, 2007, 49(2): 222-229. DOI:10.1111/j.1744-7909.2007.00393.x.
[22]LI J, NORVILLE J, ACH J, et al. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9[J]. Nature Biotechnology, 2013, 31(8): 688-691. DOI:10.1038/nbt.2654.
[23]BROOKS C, NEKRASOV V, LIPPMAN Z, et al. Efficient gene editing in tomato in the first generation using the CRISPR/Cas9 system [J]. Plant Physiology, 2014(166): 1 292-1 297. DOI: 10.1104/pp.114.
[24]SONG Y, YUAN L, WANG Y, et al. Efficient dual sgRNA-directed large gene deletion in rabbit with CRISPR/Cas9 system[J]. Cellular & Molecular Life Sciences Cmls, 2016(73):2959. DOI: 10.1007/s00018-016-2143-z.
Design and evaluation for sgRNAs targetingSlACS2 gene in CRISPR/Cas9 system
BAI Yunfeng1, ZHANG Aiping2, YAN Jianjun1, HE Feiyan3, ZHANG Weifeng1*,
(1.KeyLaboratoryofCropGeneResourcesandGermplasmEhhancementonLoessPlateau,MinistryofAgriculture,ShanxiProvinceKeyLaboratoryofCropGeneticsandMolecularImprovement(InstituteofCropScience,ShanxiAcademyofAgriculturalSciences),Taiyuan030031,China;2.InstituteofAgriculturalScience,SixthDivision,XinjiangProductionandConstructionCorps,Wujiaqu831300,China;3.CollegeofBioengineering,ShanxiUniversity,Taiyuan030006,China)
Tomatoes are typical climacteric fruits and produce large amounts of ethylene, i.e. system II ethylene with the climacteric, which makes the tomato fruits overripe and Perishable. ACC synthase 2 in tomato (SlACS2) is the key enzymy during the process of biosynthesis of the system II ethylene. Suppressing the expression of the system II ethylene will postone the fruits overripen by using CRISPR-Cas9 genome editing system to modify theSlACS2 gene. In this study the digital expression profile ofSlACS2 based on RNA-seq shows thatSlACS2 gene is expressed specifically in fruit.SlACS2 gene is located in chromosome 1 of tomato, containing four exons and three introns. In the first, second and third exon, 18, 9 and 11 sgRNAs are found by using related online tools CRISPRdirect and CRISPR-P respectively. sgRNA1-14 and sgRNA3-8, and their 12nt seed sequences proximal the protspacer adjacent motif (PAM) all are unique and no completely homologous sequences in other location of tomato genome is found. The GC content is higher than 40% and there is no termination sequence TTTT in sgRNA1-14 and sgRNA3-8. The BLAST results show that sgRNA1-14 or sgRNA3-8 is highly consistent with the sequences of the homologous sites of eightSlACS2 homologous genes, suggesting that the two sgRNAs are located in the conserved regions ofSlACS2 gene. Multiple single nucleotide polymorphisms (SNPs) are found between sgRNA1-14 or sgRNA3-8 or their respective seed sequence and the sequences of the homologous sites ofSlACS6 orSlACS4, indicating that the two sgRNAs can be used to edit theSlACS2 gene in different varieties with minimal off-target effects for other members inSlACSfamily.
Tomato; ACC synthase 2; sgRNA; CRISPR-Cas9; Genomic editing
2016-09-03;
2016-10-18.
国家自然科学基金项目(No.30971838);山西省应用基础研究项目(No.201601D011075)。
白云凤,女,博士,研究员,研究方向:植物分子遗传研究;E-mail: byfok@126.com; 张爱萍,女,研究员,研究方向:园艺植物育种;E-mail: 379409419@qq.com.
*通信作者:张维锋,男,研究员,研究方向:植物遗传育种研究;E-mail: zhwfen@sina.com.
10.3969/j.issn.1672-5565.2017.01.201609002.
Q986;S641.2
A
1672-5565(2017)01-007-09
猜你喜欢
华人时刊(2023年1期)2023-03-14
电子科技大学学报(2022年5期)2022-10-29
汉字汉语研究(2021年2期)2021-08-30
中国生殖健康(2020年4期)2021-01-18
汉字汉语研究(2019年2期)2019-08-27
中国生殖健康(2018年4期)2018-11-06
石油化工建设(2018年2期)2018-07-11
河北书画研究(2016年3期)2016-04-28
湖北农业科学(2014年11期)2014-09-10
自动化博览(2014年9期)2014-02-28
