
稻米蒸煮品质性状与分子标记关联研究
2017-04-10 02:10高维维陈思平王丽平陈立凯郭涛王慧陈志强
中国农业科学 2017年4期
高维维,陈思平,王丽平,陈立凯,郭涛,王慧,陈志强
(华南农业大学国家植物航天育种工程技术研究中心,广州 510642)
稻米蒸煮品质性状与分子标记关联研究
高维维,陈思平,王丽平,陈立凯,郭涛,王慧,陈志强
(华南农业大学国家植物航天育种工程技术研究中心,广州 510642)
【目的】调查分析代表性水稻种质重要的品质性状和淀粉RVA谱变异,筛选与性状显著关联的分子标记,为稻米品质改良提供依据。【方法】以48份国内外水稻多样性种质为材料,进行稻米品质性状变异调查分析;利用快速淀粉粘滞测定仪(rapid visco analyzer)鉴定材料淀粉RVA谱。利用已报道的稻米淀粉合成相关基因等位标记及稻米籽粒相关QTL连锁分子标记对种质材料进行基因分型。利用GLM模型进行分子标记与品质性状的关联检测,并对显著关联标记进行逐步回归分析;评估等位基因及其组合对目标性状的表型影响效应,同时鉴别对应优异等位基因型及载体品种。【结果】供试材料在直链淀粉含量(AC)、胶稠度(GC)和碱消值(ASV)等表现出广泛的表型变异和多样性,变异系数为26.5%—36.3%。RVA谱检测表明崩解值、消减值和回复值等在材料间具有明显差异,能较好地反映不同种质的淀粉糊化特性。相关分析表明,AC与冷胶黏度、消减值和回复值呈显著正相关,与最高黏度和崩解值呈显著负相关;GC同时与消减值和回复值呈显著负相关。利用154个多态性标记共检测到491个等位变异,基因多样性平均为0.447,多态信息含量(PIC)平均为0.390。性状-标记关联检测共获得22个与稻米品质性状显著关联的位点,单个关联标记位点解释的表型变异(R2)范围为14.11%—75.62%。GBSSI是影响AC和GC的主效基因,分子标记Wx-G/T对AC和GC的表型变异解释率分别为61.44%和41.87%。SSIIa是影响ASV的主效基因,alk-GC/TT和SSIIa-F对ASV的表型变异解释率分别为75.62%和74.46%。利用显著关联标记构建AC、GC和ASV的回归模型方程,决定系数分别为85.30%、40.62%和80.38%。【结论】水稻淀粉RVA谱与AC、GC和ASV密切相关,利用RVA谱可更全面地评价稻米品质性状。利用稻米品质表型-分子标记关联,共鉴定出22个与品质性状显著关联的位点,其中5个位点同时与AC和GC关联。回归模型表明标记的组合可产生不同的表型效应。
水稻;稻米品质;性状-标记关联;等位变异
0 引言
【研究意义】稻米品质的优劣对其商品价值具有重要影响。蒸煮品质是影响稻米口感的关键,成为是否满足优质稻米消费需求和影响国内外稻米市场的重要因素。其中,直链淀粉含量(amylose content,AC)、糊化温度(gelatinization temperature,GT)和胶稠度(gel consistency,GC)是稻米蒸煮品质的主要指标。近年来,利用黏度速测仪RVA进行淀粉糊化特性评价,已在中国育种实践中得到较好应用[1-2]。进一步解析RVA谱特征值与稻米蒸煮品质的相关性,对于全面评价水稻品质特性具有实践价值。此外,寻找与稻米蒸煮品质性状相关的优异等位基因及典型载体材料,探讨不同分子标记的组合对于品质性状的表型效应,对提高分子标记辅助选择(maker assisted selection,MAS)在品质育种中的应用效率具有重要意义。实现对个体基因型鉴定,高效预测表型值,有利提高稻米品质性状选择效率。【前人研究进展】稻米品质性状被认为是典型的质量-数量遗传性状,受多基因控制。显然,多位点的标记关联和效应分析是实现精确分子标记选择的前提。ZHAO等[3]通过标记与性状的关联分析,验证了粒形、垩白、淀粉合成相关QTL/基因的分子标记的可行性;KHARABINA-MASOULEH等[4]通过关联分析获得了31个与稻米蒸煮品质相关的淀粉合成基因SNP位点;邱先进等[5]利用全基因组关联分析解析籼稻垩白的遗传基础和挖掘影响垩白粒率和垩白度的优异等位基因。整体而言,虽然许多稻米品质相关QTL和功能基因,包括影响粒形、垩白性状的QTL[6-8]和淀粉合成相关基因[9-11]得到鉴定,但涉及等位基因组合以及优异等位变异的解析仍是有限。【本研究切入点】稻米品质性状的QTL定位和基因鉴定研究已有较多报道,水稻淀粉合成相关基因与稻米品质的关联分析[12-15]也获得重要进展,但对等位基因组合、优异等位变异及典型载体材料挖掘的研究尚不多。大量已报道的相关分子标记的可用性、变异效应以及贡献力并未明确。【拟解决的关键问题】本研究利用淀粉合成相关基因及稻米籽粒相关位点连锁的分子标记对代表性种质进行分型检测,并对多个稻米品质性状进行基于线性模型的等位基因贡献性检测,挖掘与稻米品质性状显著相关的标记和重要等位基因组合,为水稻品质改良奠定基础。
1 材料与方法
1.1 试验材料
供试材料为华南农业大学国家植物航天育种工程技术研究中心种质资源库所收集的国内外种质材料48份,其中,籼稻38份、粳稻9份和爪哇稻1份(表1)。
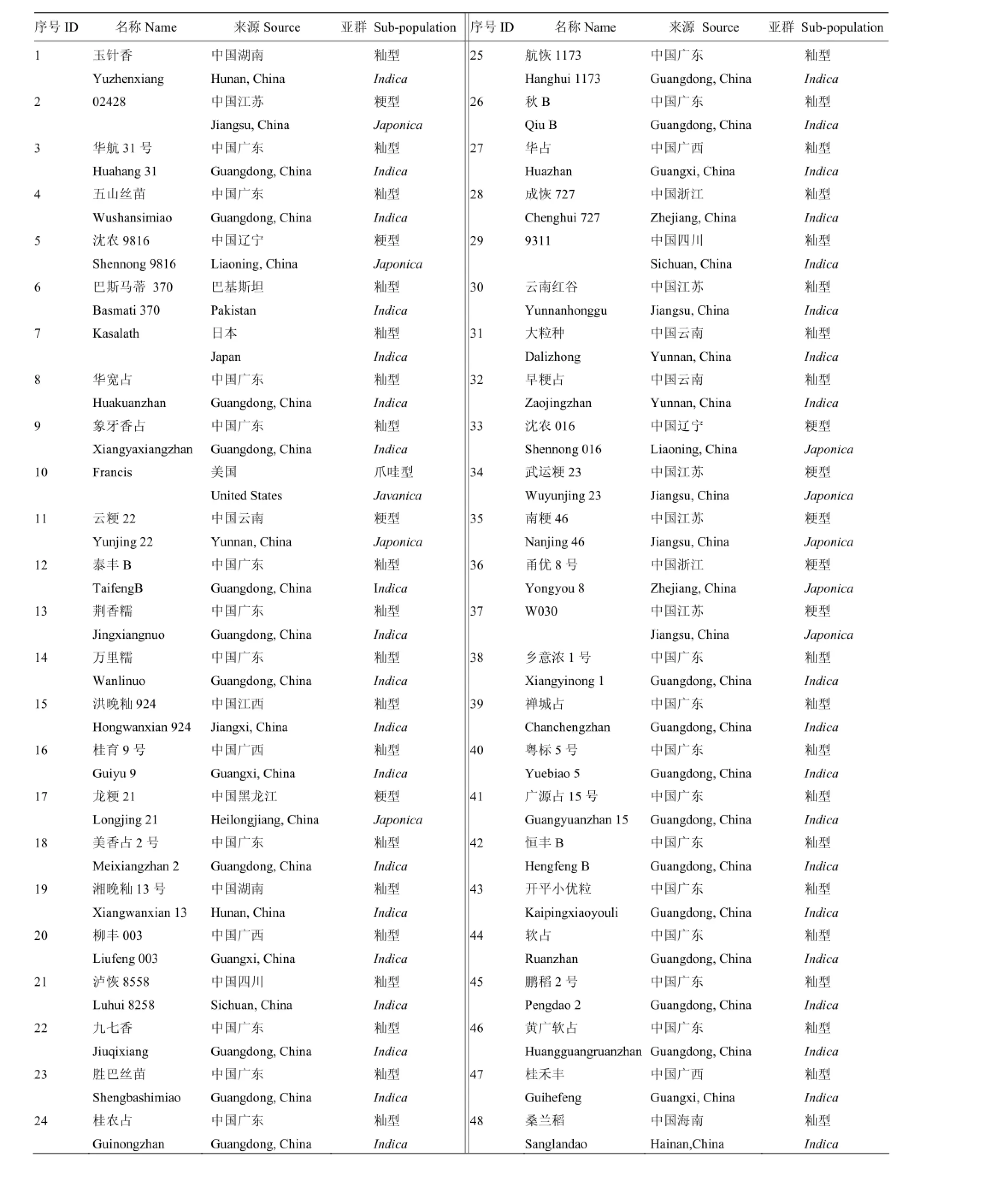
表1 本研究用到的水稻种质材料Table 1 Rice germplasm materials used in this study
1.2 稻米品质性状鉴定
供试材料于2015年晚季种植于国家植物航天育种工程技术研究中心试验基地(广州)。田间管理(水、肥、病虫害防治等)按当地大田常规栽培要求实施。按农业部部颁标准NY147-88[16]测定直链淀粉含量(AC)、胶稠度(GC)和碱消值(ASV)。淀粉RVA谱测定采用波通瑞华科学仪器(北京)有限公司生产的RVA(Rapid Visco Analyzer, Model 3D)快速测定淀粉粘滞特性,并用TCW(Thermal Cycle for Windows)配套软件分析。测定按AACC规程(1995 61-02)要求,即含水量为14.0%时,样品量3.00 g,蒸馏水25.00 mL。测定过程中罐内温度变化为50℃保持1 min,以12℃/min上升到95℃(3.75 min),95℃保持2.5 min,以12℃/min下降到50℃(3.75 min),50℃保持1.4 min。搅拌器在起始10 s内转动速度为960 r/min,以后保持在160 r/min。RVA谱特征用最高黏度(peak viscosity,PKV)、热浆黏度(hot paste viscosity,HPV)和冷胶黏度(cool paste viscosity,CPV)描述。在TCW程序中选用水稻的分析方法( rice method),其中,程序参数设置为:Temp At Visc Rate(2,7,1,36),成糊温度(pasting temperature,PTm)按包劲松[17]的方法手工计算获得。黏度单位用“Rapid Visco Units(RVU)”或厘泊(centipoise,cp)表示。
1.3 分子标记检测
采用CTAB法提取水稻材料叶片基因组DNA。试验筛选所涉及的分子标记包含一部分报道的淀粉合成相关基因的等位分子标记;以及稻米籽粒有关性状连锁的标记,涉及SNP、SSR和STS等标记类型。经初步筛选获得用于关联分析的多态性标记共154个(见电子附表)。所有引物均由金唯智生物技术有限公司合成。不同引物的PCR反应体系及扩增程序参照ZHAO等[3]方法。利用自动荧光毛细管电泳(Fragment Analyzer™)检测PCR扩增产物,并用PROSize 2.0软件对电泳结果进行分析,确定产物DNA片段长度,记录基因型信息。
1.4 数据处理
利用Excel和SAS 9.3进行性状的统计分析。对于多元线性回归分析,将各位点的等位变异转化为虚拟变量,具体参照LEE等[18]的方法。利用Power Marker 3.25软件计算每个位点的等位基因数(number of alleles per locus)、基因多样性(gene diversity)和多态信息含量(polymorphism information content, PIC)。以Structure 2.3.1软件进行群体遗传结构的分析,估计最佳群体组群数K,其取值范围为1—10,将MCMC(markov chain monte carlo)开始时的不作数迭代(length of burn-in period)设为10 000次,再将不作数迭代后的MCMC设为10 000次,迭代次数(number of iterations)设置为5,计算Q参数。采用Tassel 3.0软件一般线性模型(general linear model,GLM),以Q作为协变量,利用表型变异数据对标记逐一进行回归分析,并计算标记对表型变异的解释率。参考BRESEGHELLO[19]和文自翔等[20]方法计算标记位点等位变异的表型效应。
2 结果
2.1 稻米品质相关性状的变异分析和RVA谱分析
通过稻米品质性状的调查,48份种质材料中检测到广泛的遗传变异。其中,AC为5.9%—27.5%、GC为24.5—142.5 mm、ASV为1.0—7.0;AC、GC和ASV的变异系数为26.5%—36.3%(表2)。说明所选材料表现出较高的表型多样性。
基于黏度速测仪测定的RVA谱主要反映水稻淀粉谱粘滞特性。根据与淀粉特性密切相关的蒸煮品质性状,即AC、GC和ASV,选取18份具有代表性的品种(表3)进行RVA谱特征值分析。结果表明,不同品种RVA谱及其特征值存在明显的差异。其中,消减值(setback,SBV)的变异系数最大,其次是回复值(consistence,CSV)和崩解值(breakdown,BDV),其余RVA谱特征值的变异系数均较小。不同水稻品种间RVA谱特征值在PKV、HPV、CPV、PTm和峰值时间(peak time,PaT)上的差异不大,但BDV、SBV和CSV等指标表现明显差异,具有品种特异性。2份糯稻品种的峰值时间最短,为3.6和3.7 min;并且具有最小的SBV和CSV,明显区分于其他非糯种质。
稻米品质性状与RVA谱特征值相关性分析(表4)发现,AC分别与CPV、SBV和CSV呈显著或极显著正相关,与PKV和BDV呈负相关。其中,AC与SBV相关系数达0.794。高直链淀粉的2份材料,Kasalath(AC 27.2%)和泰丰B(AC 27.0%)的SBV均为正值,远高于其他非糯材料。从表3和表4可以看出,不同品种因本身AC和GC的不同,RVA谱显著不同。糯稻品种(万里糯、荆香糯)RVA谱特征值表现为BDV较大,CPV小,SBV为负值且绝对值较大,达到PKV时间短;高直链淀粉
含量和胶稠度较小的品种(Kasalath、泰丰B)RVA谱特征值表现为BDV较小,SBV较大。由此可见,稻米品质中的AC和GC是影响RVA谱特征值的2个重要理化指标。此外,GC分别与SBV和CSV呈显著负相关;ASV与PTm呈极显著负相关。RVA特征值中,SBV与CSV同时与AC和GC显著相关,是反映稻米品质最为重要的2个指标。而利用经验公式换算的PTm与ASV相关系数达0.910,表明该方法测定的PTm可精确地反映稻米淀粉糊化温度。
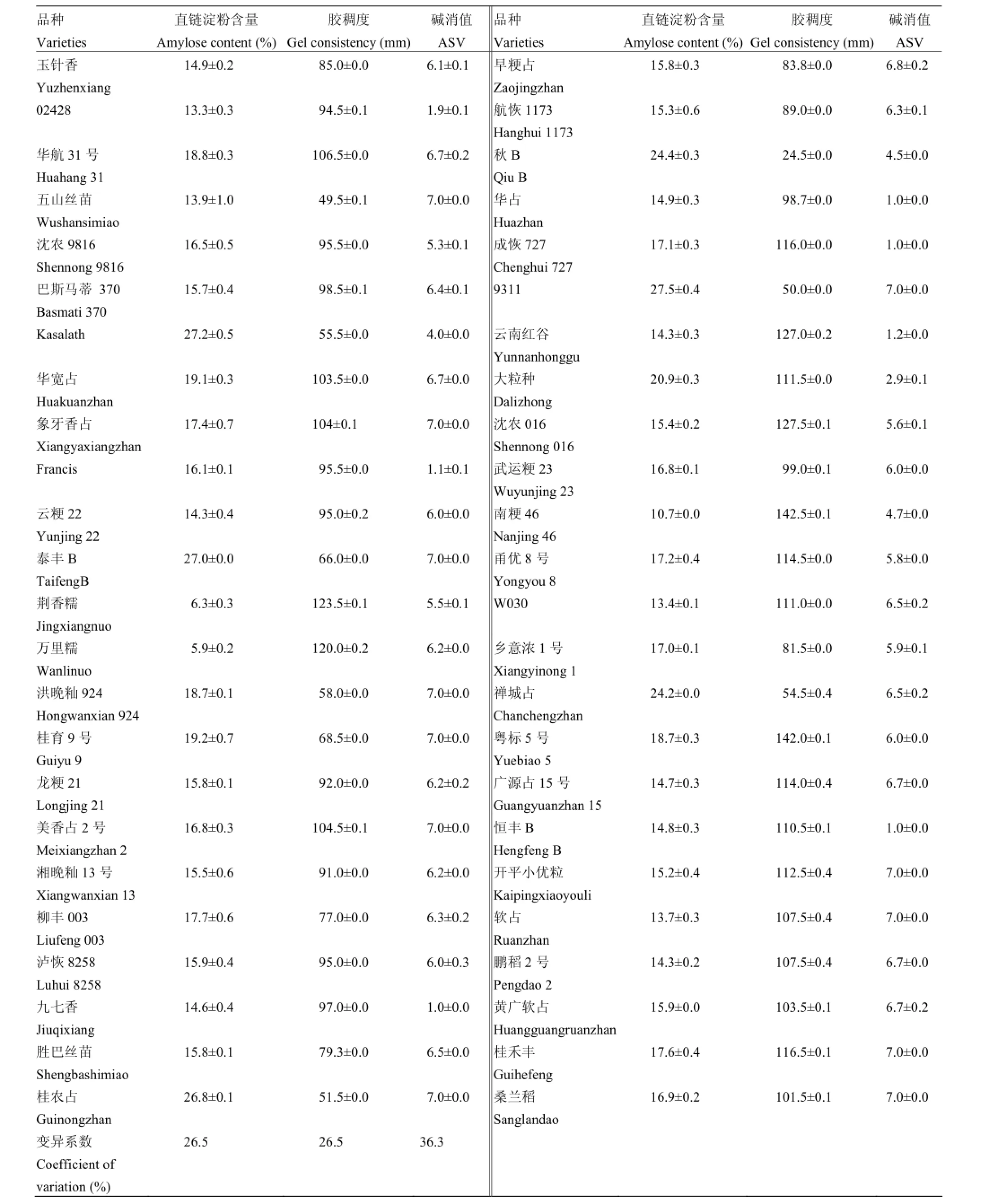
表2 48份水稻材料的稻米品质性状值Table 2 Grain-quality traits of the 48 rice materials
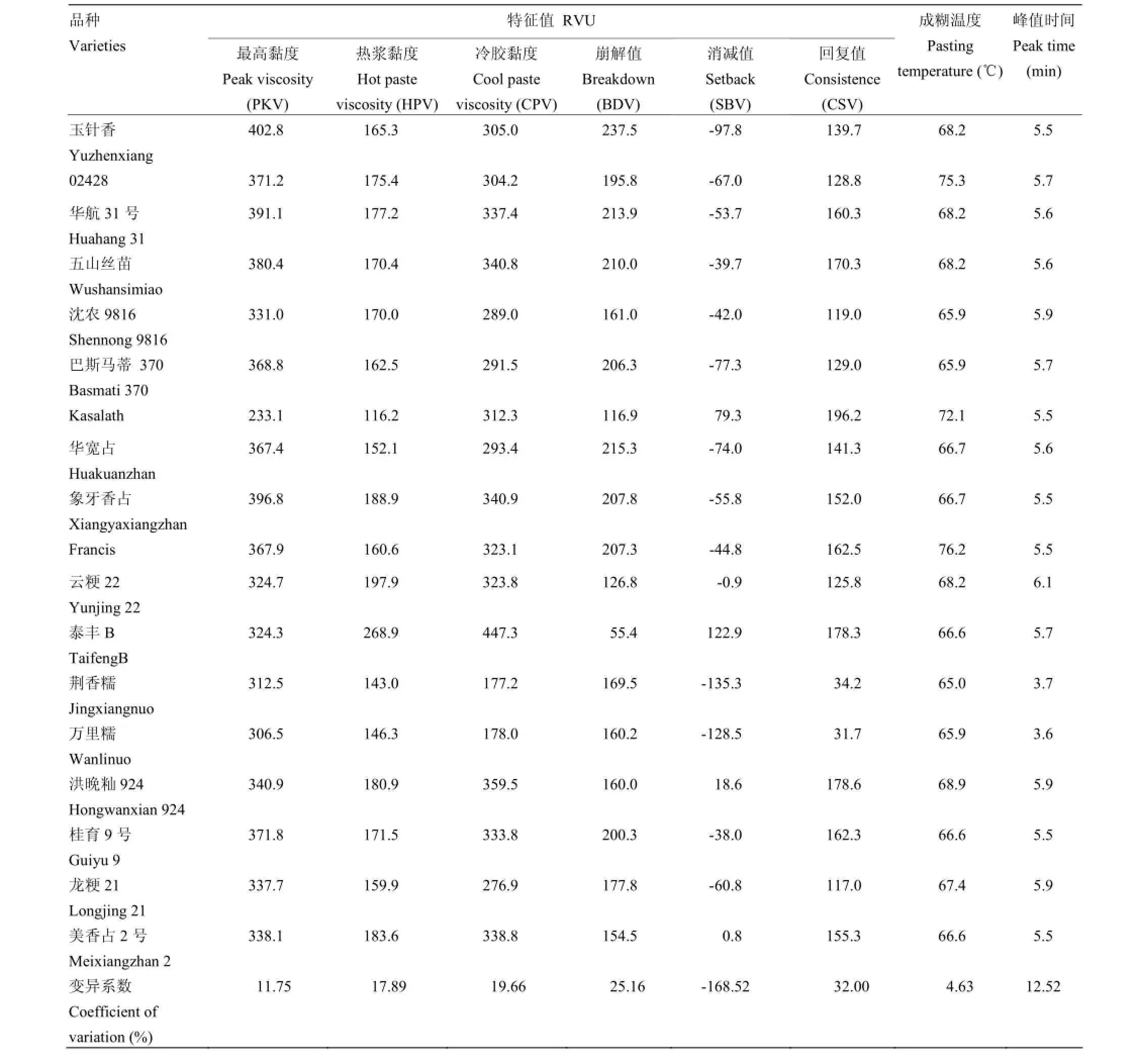
表3 18个品种RVA谱特征值Table 3 RVA profile characteristics of 18 rice varieties
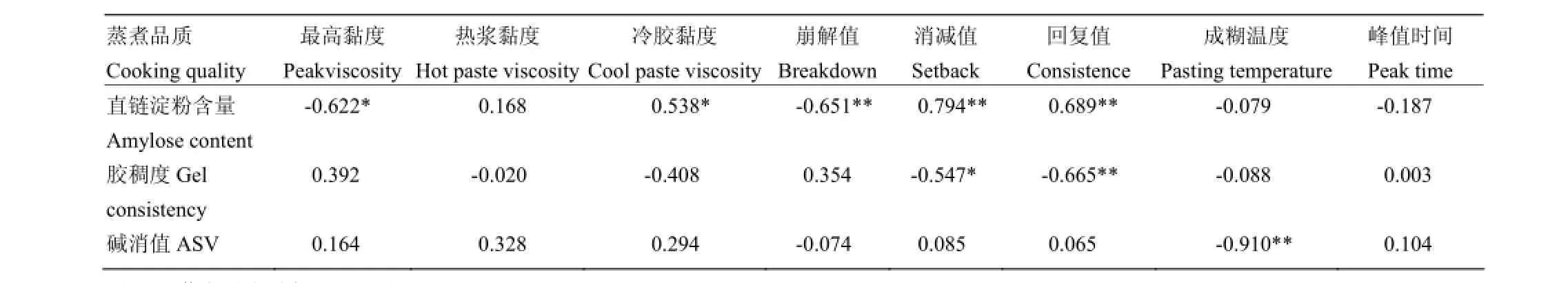
表4 稻米品质性状与RVA谱特征值的相关性Table 4 Correlation between rice quality characteristics and RVA profile characteristics
2.2 标记-等位基因关联性检测
154个多态性标记中共检测到491个等位变异,平均每个位点3.19个,不同位点等位基因数变幅范围为2—8。各位点的基因多样性平均为0.447,变幅为0.041—0.829。对于多态性信息含量(PIC),标记间差异较大,幅度为0.040—0.806,平均值为0.390,其中,27对引物为低度多态位点,82对引物为中度多态位点,45对引物为高度多态位点。
采用EVONNO等[21]介绍的方法进行群体结构分析表明,△K值于亚群数为2时有明显的峰。因而该群体的适宜亚群数确定为2。第一个亚群包括02428、沈农9816、沈农016、云粳22、龙粳21、武运粳23、南粳46、W030、甬优8号和大粒种等粳稻亚群;其他材料分布在第二个亚群,为籼稻亚群。利用Tassel 3.0 软件中的GLM模型,以各材料对应的Q值作为协变量进行关联分析,结果显示,在0.01水平下,对于不同性状共检测到22个关联标记位点,其中同时与AC和GC关联的位点有5个。单个关联标记位点的贡献率(R2)范围是14.11%—75.62%。
标记-性状关联结果表明,与AC关联的位点有15个,表型变异解释率范围为14.34%—61.44%,其中在3个淀粉合酶基因(GBSSI、BeIIb和SSI)中有9个位点,贡献率最高的是Wx-G/T;与GC关联的位点有8个,表型变异解释率范围为14.11%—41.87%,在3个淀粉合酶基因(GBSSI、SSI和SSIIIb)中有5个位点,贡献率最高的是Wx-G/T;与ASV关联的位点有4个,表型变异解释率范围为19.01%—75.62%,其中,SSIIa的2个位点(alk-GC/TT和SSIIa-F)的贡献率是所有关联位点中最高的,分别为75.62%和74.46%(表5)。4个GBSSI的位点、4个BEIIb的位点以及1个SSI的位点都与AC显著关联,GBSSI和SSI上都有2个位点与GC显著关联,2个SSIIa的位点都与ASV显著关联,表明GBSSI、BEIIb和SSI对AC和GC具有一定的决定作用,SSIIa对ASV具有一定的决定作用。其中,标记Wx-G/T、GBSSI-4-IF、SSI-2-F、INV1和RM15206同时与AC和GC关联。
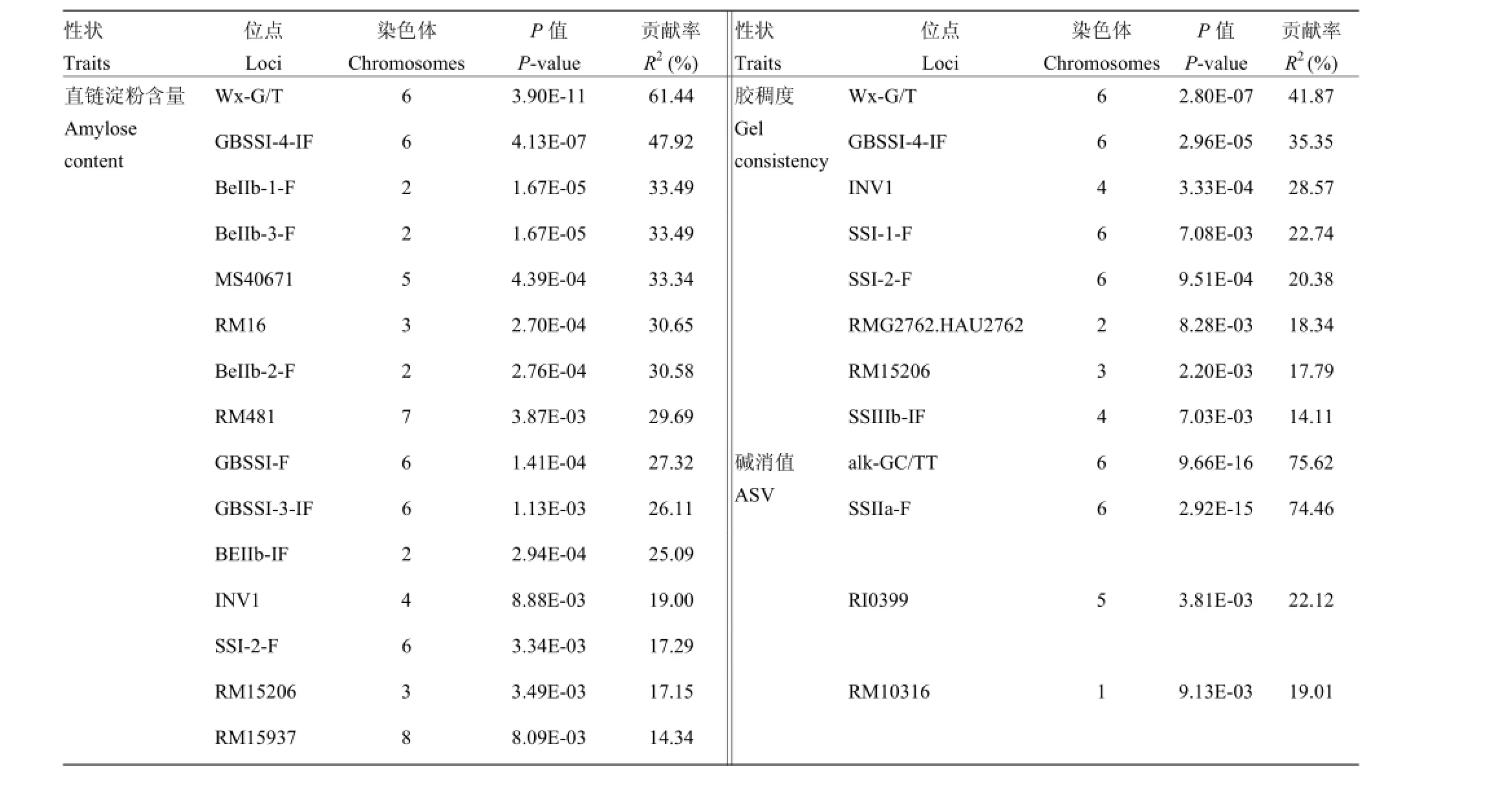
表5 稻米品质性状显著关联的标记位点Table 5 Marker loci significantly associated with grain-quality traits
2.3 关键稻米品质性状等位基因及组合的效应分析
与AC、GC和ASV显著关联且贡献率位于前4位的位点上等位基因效应差值及对应等位基因的载体材料如表6所示。与AC显著关联的位点中,Wx-G/T具有最大的表型效应差异,Wx-G/T-G与Wx-G/T-T的表型效应差值达到了10.6%。AC减效最大的等位变异是Wx-G/T-T,典型载体材料是W030和云粳22。
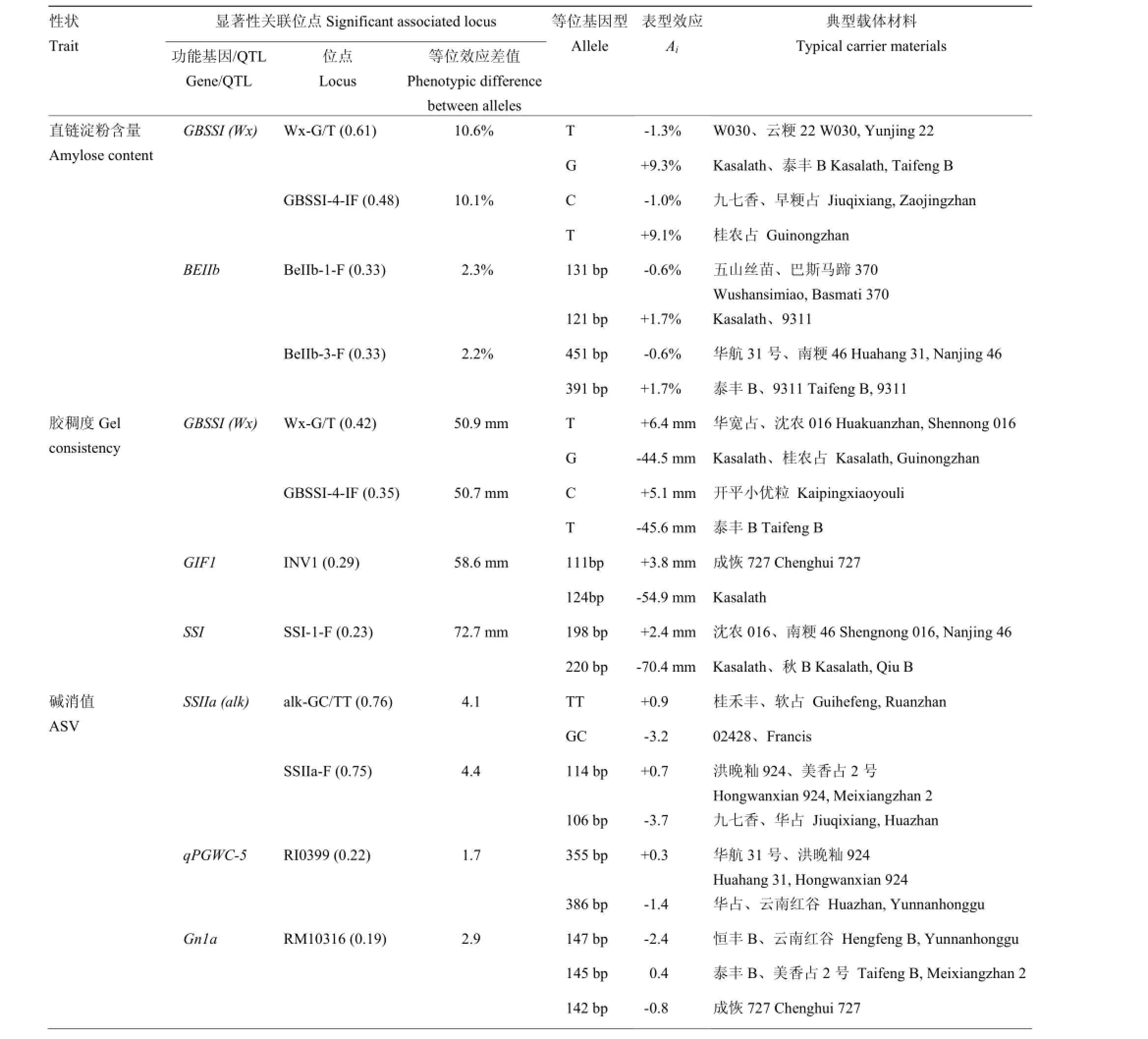
表6 重要品质性状关联位点及表型效应Table 6 Marker loci significantly associated with important grain quality traits and their phenotypic variation
SSI-1-F对GC具有最大的表型效应差异(72.7 mm),其次是贡献率最大的Wx-G/T(+50.9 mm)。增效最大的等位变异是Wx-G/T-T(+6.4 mm),典型载体材料是华宽占和沈农016,其次是GBSSI-4-IF-C(+5.1 mm),典型载体材料是开平小优粒。
与ASV显著关联的位点等位基因间表型效应差异最大的是SSIIa-F(4.4),其次是贡献率最大的alk-GC/TT(4.1),其中,alk-GC/TT-TT具有最大的增效表型效应(+0.9),典型载体材料是桂禾丰和软占。
其中,典型载体材料W030、开平小优粒和软占等的AC、GC和ASV值达国标优质(表2),GBSSI的Wx-G/T、GBSSI-4-IF和SSIIa的alk-GC/TT位点的基因型分别为T/T、C/C、TT/TT,均为优异等位变异。因此,W030、开平小优粒和软占等携带了控制AC、GC及ASV的多个优异等位基因,可作为优质品种选育的骨干亲本。
利用逐步回归的方法对各性状显著性关联位点进行分析的结果表明,AC、GC和ASV逐步回归模型中分别包含5、2和2个关联标记(表7、图1)。不同的等位基因组合形成了广泛的表型差异。Wx-G/T、GBSSI-F、RM16、GBSSI-3-IF和RM15206 5个位点的组合对AC的决定系数为85.30%,其中,Wx-G/T和GBSSI-F为前人验证过的控制AC的主效基因GBSSI的位点,具有最高的决定系数(68.60%和12.08%)其他3个为本研究获得的新位点。不同的等位基因组合具有不同的AC值,其中,G/I/II/A/I、G/I/ II/A/II和G/I/II/C/I等位基因组合具有较高的AC,T/II/I/A/II等位基因组合为糯稻品种,AC值极低;Wx-G/T和RMG2762.HAU2762可以用来预测GC,决定系数为40.62%,表明所选标记对GC进行预测尚有一定的困难;alk-GC/TT和SSIIa-F 2个位点的组合可以用来预测ASV,决定系数为80.38%,alk-GC/TT具有主要的决定作用(76.71%),GC/I和GC/II等位基因组合具有较低的ASV,TT/II等位基因组合具有较高的ASV。
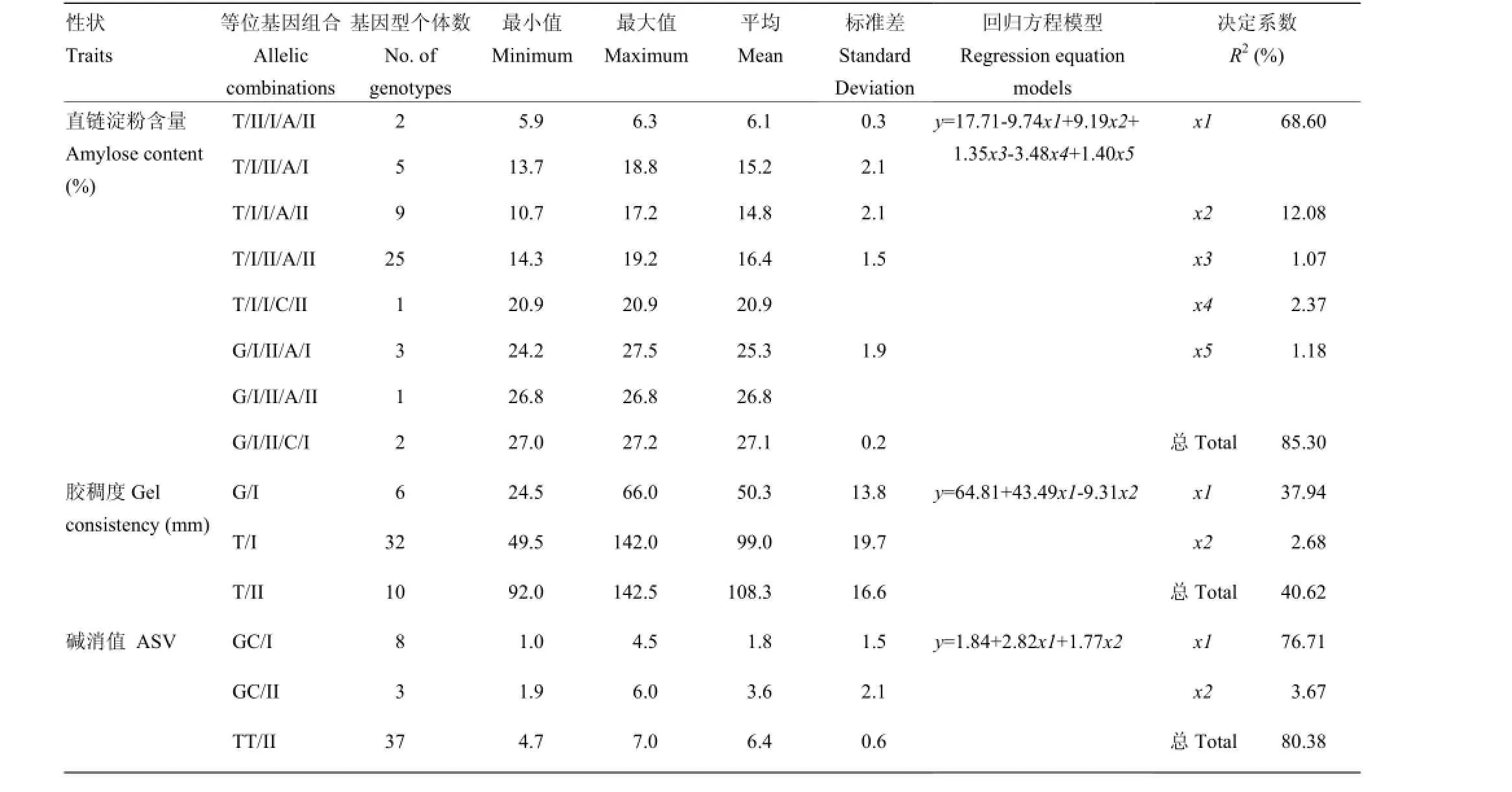
表7 逐步回归模型中标记的等位基因组合Table 7 Allelic combinations based on the stepwise regression models
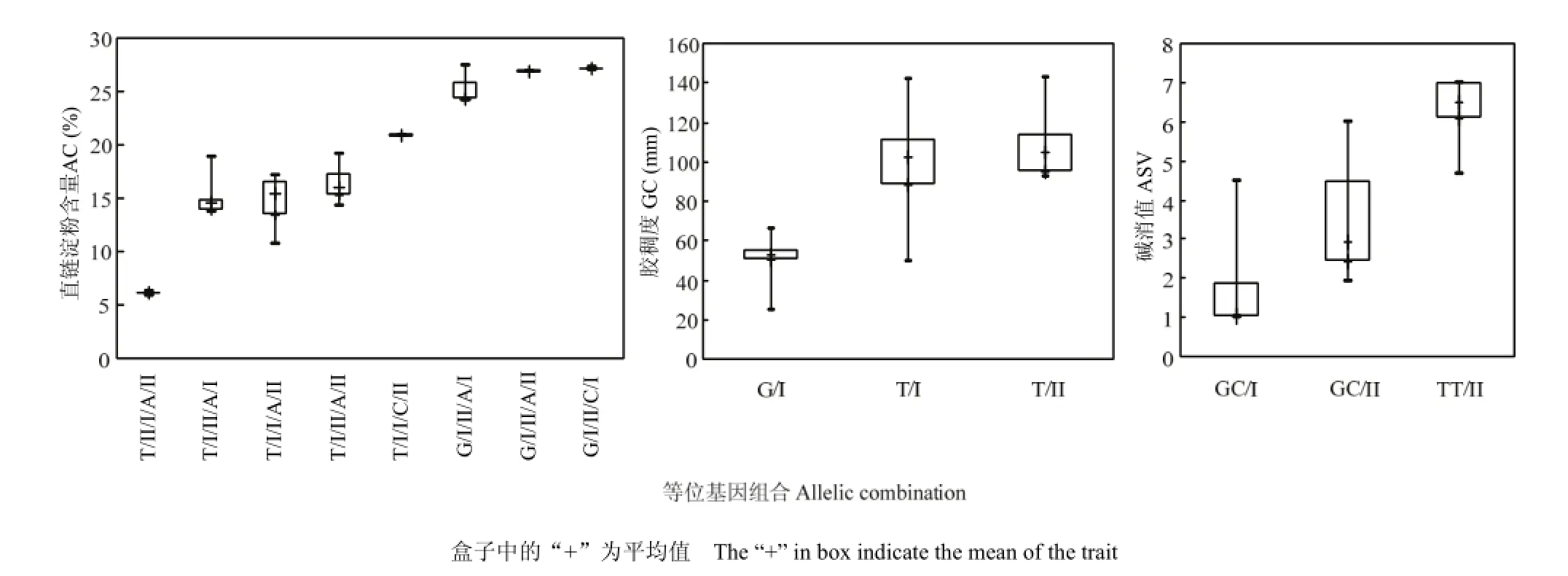
图1 等位基因组合箱形图Fig. 1 Box plots of the allelic combinations
3 讨论
本试验代表性品种的稻米淀粉RVA谱检测结果表明,不同品种的RVA谱及其特征值存在明显的差异,消减值的变异系数最大,其次是回复值和崩解值,这与何秀英等[22]的结果类似,但变异系数均更大,主要是由于所选材料在AC和GC上差异显著。相关性分析发现,RVA特征值与AC和GC存在显著或极显著的相关性,稻米品质中的AC和GC是影响RVA谱特征值的2个重要理化指标,这与前人研究结果相似[23],但其相关系数略小于贾良等[24]的研究结果,这可能是由于所用材料数较少。
本研究所用154个多态性标记,共检测到491个等位变异,平均每个标记位点具有3.19个等位变异,变幅为2—8,平均每对引物的多态信息含量(PIC)为0.390,变幅为0.040—0.806,遗传多样性平均为0.447,略低于前人研究结果[25],这是因为本研究所用的标记部分为SNP标记,为双等位形式,每个位点所携带的多态信息较少。
通过对48份不同水稻品种与稻米品质相关分子标记的关联性分析,发现对于20个与淀粉合成酶相关的基因,稻米AC主要由GBSSI控制,GC也可能主要由GBSSI或与其紧密连锁的基因位点控制[26]。本研究中Wx-G/T对AC和GC的贡献率最大,分别为61.44%和41.87%,同时GBSSI的其他4个位点对AC和GC的贡献率均较大(>25%),说明GBSSI是控制直链淀粉含量和胶稠度的主效基因,这与TIAN等[27]的研究结果相符。GBSSI-4-IF与GC显著关联,这与第10个外显子的C/T SNP对决定高直链淀粉米饭的软硬具有重要作用是一致的[28]。多数研究表明,SSIIa(尤其是第8外显子的GC/TT SNP位点)对糊化温度具有重要影响[29-30]。本研究中SSIIa的2个位点与ASV均显著相关。除了SSIIa外,未检测到其他淀粉合成相关基因的位点与碱消值相关,这可能与多数参试材料的ASV较大有关。
以往研究表明,影响GC的主效基因是Wx基因(GBSSI),但本研究中,在Wx基因型一样的情况下,出现GC表型差异较大的现象,如南粳46和五山丝苗、广源占15号和洪晚籼924等的Wx基因型均为T/T,但GC值差异很大。究其原因,可能有以下几点:(1)基因存在本研究未涉及的其他变异位点;存在某些控制GC的微效基因与Wx基因互作,但微效基因与Wx基因互作关系研究仍未明确;(2)GC受基因型与环境互作影响较大,遗传力较低[31-32]。影响糊化温度的主效基因是alk(SSIIa)[33],本研究中,云粳22的alk基因型为GC/GC,但其碱消值较大(糊化温度低),这与前人研究结论未吻合[33]。NAKAMURA等[34]研究表明,不同SNP位点的相互作用会影响SSIIa酶的活性,在4 198 bp处出现G/A SNP的A位点时,不论SSIIa基因型为GC/GC或TT/TT,均表现出低糊化温度,这可能是导致云粳22表现出低糊化温度的原因。
单个标记用于区分群体的不同种质具有一定的局限性,对显著关联标记构建的回归模型可以用来预测表型,LESTARI等[35]建立了多个用来估计稻米食味品质的回归模型。本试验通过对与AC、GC和ASV显著关联的位点进行逐步回归分析发现,Wx-G/T、GBSSI-F、RM16、GBSSI-3-IF和RM15206 5个位点的组合可以用来预测AC;Wx-G/T和RMG2762.HAU2762可以用来预测GC,但决定系数较低;alk-GC/TT和SSIIa-F 2个位点的组合可以用来预测ASV。其中,Wx-G/T在AC和GC回归方程模型中均具有较大的决定系数,alk-GC/TT在ASV回归方程模型中的决定系数高达76.71%,表明对于AC和GC,Wx-G/T是主效位点,对于ASV,alk-GC/TT是主效位点,这与前人研究结果一致[26,29]。可知,所构建的回归方程模型对分子标记辅助选择育种具有一定的指导意义,尤其是决定系数较大的AC和ASV的回归模型。
挖掘稻米品质性状的优异等位基因及对应的典型载体材料,可为水稻分子设计育种的应用奠定基础。本研究通过对与AC、GC、ASV显著相关且贡献率位于前4位的位点的等位变异分析发现,同一位点等位变异间表型效应有差异。对于AC,Wx-G/T位点对其贡献率最大,且Wx-G/T-T具有最大的减效表型效应,典型载体材料为W030和云粳22;对于GC,贡献率最大的是Wx-G/T,最优等位变异是Wx-G/T-T,典型载体材料是华宽占和沈农016,其次是GBSSI-4-IF-C,典型载体材料是开平小优粒;对于ASV,贡献率最大的是alk-GC/TT,最优等位变异是alk-GC/TT-TT,典型载体材料是桂禾丰和软占。在水稻品质改良中,利用这些淀粉合成相关基因的优异等位基因及载体材料信息进行育种设计、杂交组合配置,从而将不同位点的优异等位基因进行聚合,以达到改良品质性状的目的。例如可利用携带控制AC、GC、ASV的关联位点优异等位变异的材料W030、开平小优粒、软占等作为骨干亲本或者供体亲本。这些材料具有对AC减效效应的等位变异位点Wx-G/T-T、对GC增效效应的等位变异位点GBSSI-4-IF-C和对ASV增效效应的等位变异位点alk-GC/TT-TT。针对现有育种材料开展分子改良或者多基因聚合,实现不同遗传背景的有利等位基因利用。
4 结论
共检测到22个与稻米品质性状显著关联的位点,其中同时与AC和GC关联的位点有5个。单个关联标记位点解释的表型变异比例(R2)范围是14.11%—75.62%。对表型变异解释率最大的是SSIIa的alk-GC/TT和SSIIa-F位点以及GBSSI的Wx-G/T和GBSSI-4-IF位点,表明GBSSI和SSIIa是影响AC、GC和ASV的2个主效基因。利用逐步回归分析获得了可以预测AC、GC和ASV的回归方程模型以及等位基因组合,同时发掘了AC、GC和ASV的优异等位变异及载体材料。
[1] CHENG F, ZHONG L, SHU Q. Studies on the cooking and eating quality properties in chalky milled grains of earlyindicarice.Acta Agronomica Sinica, 2002, 28(3): 363-368.
[2] 胡培松, 翟虎渠, 唐绍清, 万建民. 利用RVA快速鉴定稻米蒸煮及食味品质的研究. 作物学报, 2004, 30(6): 519-524.
HU P S, ZHAI H Q, TANG S Q, WAN J M. Rapid evaluation of rice cooking and palatability quality by RVA profile.Acta Agronomica Sinica, 2004, 30(6): 519-524. (in Chinese)
[3] ZHAO X, ZHOU L, PONCE K, YE G. The usefulness of known genes/QTLs for grain quality traits in anIndicapopulation of diverse breeding lines tested using association analysis.Rice, 2015, 8(1): 1-13.
[4] KHARABIAN-MASOULEH A, WATERS D L E, REINKE R F, WARD R, HENRY R J. SNP in starch biosynthesis genes associated with nutritional and functional properties of rice.Scientific Reports, 2012, 2(8): 1-9.
[5] 邱先进, 袁志华, 陈凯, 杜斌, 何文静, 杨隆维, 徐建龙, 邢丹英,吕文恺. 用全基因组关联分析解析籼稻垩白的遗传基础. 作物学报, 2015, 41(7): 1007-1016.
QIU X J, YUAN Z H, CHEN K, DU B, HE W J, YANG L W, XU J L, XING D Y, LÜ W K. Genetic dissection of grain chalkiness inIndicamini-core germplasm using genome-wide association method.Acta Agronomica Sinica, 2015, 41(7): 1007-1016. (in Chinese)
[6] XU C, LIU Y, LI Y, XU X, XU C, LI X, XIAO J, ZHANG Q. Differential expression of GS5 regulates grain size in rice.Journal of Experimental Botany, 2015, 66(9): 2611-2623.
[7] WANG S, LI S, LIU Q, WU K, ZHANG J, WANG S, WANG Y, CHEN X, ZHANG Y, GAO C. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality.Nature Genetics, 2015, 47(8): 949-954.
[8] LI Y, FAN C, XING Y, YUN P, LUO L, YAN B, PENG B, XIE W, WANG G, LI X. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice.Nature Genetics, 2014, 46(4): 398-404.
[9] CHEN M, BERGMAN C, PINSON S, FJELLSTROM R. Waxy genehaplotypes: Associations with apparent amylose content and the effect by the environment in an international rice germplasm collection.Journal of Cereal Science, 2008, 47(3): 536-545.
[10] SU Y, RAO Y, HU S, YANG Y, GAO Z, ZHANG G, LIU J, HU J, YAN M, DONG G. Map-based cloning proves qGC-6, a major QTL for gel consistency ofjaponica/indicacross, responds by Waxy in rice (Oryza sativaL.).Theoretical and Applied Genetics, 2011, 123(5): 859-867.
[11] GAO Z, ZENG D, CUI X, ZHOU Y, YAN M, HUANG D, LI J, QIAN Q. Map-based cloning of the ALK gene, which controls the gelatinization temperature of rice.Science in China Series C: Life Sciences, 2003, 46(6): 661-668.
[12] 池彦衡. 稻种资源稻米品质评价及可溶性淀粉合酶基因序列多态性和关联分析[D]. 南昌: 江西农业大学, 2011.
CHI Y H. The quality evaluation of rice resources and sequence polymorphism and association analysis of rice soluble starch synthase gene[D]. Nanchang: Jiangxi Agricultural University, 2011. (in Chinese)
[13] 林华. 水稻可溶性淀粉合酶基因SSIIa、SSIIIa等位变异及其与稻米品质的关联分析[D]. 南昌: 江西农业大学, 2011.
LIN H. The research of allelic diversification of soluble starch synthase geneSSIIaandSSIIIaand its association with rice quality[D]. Nanchang: Jiangxi Agricultural University, 2011. (in Chinese)
[14] 徐卫. 水稻可溶性淀粉合酶基因与产量和稻米品质性状的关联分析[D]. 南昌: 江西农业大学, 2013.
XU W. The association analysis of rice soluble starch synthase gene with yield and quality traits[D]. Nanchang: Jiangxi Agricultural University, 2013. (in Chinese)
[15] YANG F, CHEN Y, TONG C, HUANG Y, XU F, LI K, CORKE H, SUN M, BAO J. Association mapping of starch physicochemical properties with starch synthesis-related gene markers in nonwaxy rice (Oryza sativaL.).Molecular Breeding, 2014, 34(4): 1747-1763.
[16] 中华人民共和国农业部部颁标准. NY147-88稻米品质评价方法.北京: 中国标准出版社, 1988: 4-6.
Chinese Ministry of Agriculture.NY147-88, The Method of Rice Quality Evaluation. Beijing: Chinese Criteria Press, 1988: 4-6. (in Chinese)
[17] 包劲松. 应用RVA测定米粉淀粉成糊温度. 中国水稻科学, 2007, 21(5): 543-546.
BAO J S. Accurate measurement of pasting temperature of rice flour by a rapid Visco-Analyser.Chinese Journal of Rice Science, 2007, 21(5): 543-546. (in Chinese)
[18] LEE C, PARK J, KIM B, SEO J, LEE G, JANG S, KOH H. Influence of multi-gene allele combinations on grain size of rice and development of a regression equation model to predict grain parameters.Rice, 2015, 8(1). 1-10.
[19] BRESEGHELLO F. Association mapping of kernel size and milling quality in wheat (Triticum aestivumL.) cultivars.Genetics, 2005, 172(2): 1165-1177.
[20] 文自翔, 赵团结, 郑永战, 刘顺湖. 中国栽培和野生大豆农艺及品质性状与SSR标记的关联分析.Ⅱ优异等位变异的发掘. 作物学报, 2008, 34(8): 1339-1349.
WEN Z X, ZHAO T J, ZHENG Y Z, LIU S H. Association analysis of agronomic and quality traits with SSR markers inGlycine maxandGlycine sojain China: II. Exploration of elite alleles.Acta Agronomica Sinica, 2008, 34(8): 1339-1349. (in Chinese)
[21] EVANNO G, REGNAUT S, GOUDET J. Detecting the number of clusters of individuals using the software structure: A simulation study.Molecular Ecology, 2005, 14(8): 2611-2620.
[22] 何秀英, 程永盛, 刘志霞. 国标优质籼稻的稻米品质与淀粉RVA谱特征研究. 华南农业大学学报, 2015, 36(3): 37-44.
HE X Y, CHENG Y S, LIU Z X. Studies on the rice quality and starch RVA profile characteristics ofindicarice varieties with national high-quality.Journal of South China Agricultural University, 2015, 36(3): 37-44. (in Chinese)
[23] 隋炯明, 李欣, 严松, 严长杰, 张蓉, 汤述翥, 陆驹飞, 陈宗祥, 顾铭洪. 稻米淀粉RVA谱特征与品质性状相关性研究. 中国农业科学, 2005, 38(4): 657-663.
SUI J M, LI X, YAN S, YAN C J, ZHANG R, TANG S Z, LU J F, CHEN Z X, GU M H. Studies on the rice RVA profile characteristics and its correlation with the quality.Scientia Agricultura Sinica, 2005, 38(4): 657-663. (in Chinese)
[24] 贾良, 丁雪云, 王平荣, 邓晓建. 稻米淀粉RVA谱特征及其与理化品质性状相关性的研究. 作物学报, 2008, 34(5): 790-794.
JIA L, DING X Y, WANG P R, DENG X J. Rice RVA profile characteristics and correlation with the physical/chemical quality.Acta Agronomica Sinica, 2008, 34(5): 790-794. (in Chinese)
[25] JIN L, LU Y, XIAO P, SUN M, CORKE H, BAO J. Genetic diversity and population structure of a diverse set of rice germplasm for association mapping.Theoretical and Applied Genetics, 2010, 121(3): 475-487.
[26] 孙业盈, 彦吕, 董春林, 王平荣. 水稻Wx基因与稻米AC、GC和GT的遗传关系. 作物学报, 2005, 31(5): 535-539.
SUN Y Y, YAN L, DONG C L, WANG P R. Genetic relationship among Wx gene, AC, GC and GT of rice.Acta Agronomica Sinica, 2005, 31(5): 535-539. (in Chinese)
[27] TIAN Z, QIAN Q, LIU Q, YAN M, LIU X, YAN C, LIU G, GAO Z, TANG S, ZENG D, WANG Y, YU J, GU M, LI J. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities.Proceedings of the National Academy of Science of the USA, 2009, 106(51): 21760-21765.
[28] TRAN N A, DAYGON V D, RESURRECCION A P, CUEVAS R P, CORPUZ H M, FITZGERALD M A. A single nucleotide polymorphism in the Waxy gene explains a significant component of gel consistency.Theoretical and Applied Genetics, 2011, 123(4): 519-525.
[29] BAO J S, CORKE H, SUN M. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (Oryza sativaL.).Theoretical and Applied Genetics, 2006, 113(7): 1171-1183.
[30] CUEVAS R P, DAYGON V D, CORPUZ H M, NORA L, REINKE R F, WATERS D L, FITZGERALD M A. Melting the secrets of gelatinisation temperature in rice.Functional Plant Biology, 2010, 37(5): 439-447.
[31] LIN J, SHI C, WU M, WU J. Analysis of genetic effects for cooking quality traits of japonica rice across environments.Plant Science, 2005, 168(6): 1501-1506.
[32] SU Y, RAO Y, HU S, YANG Y, GAO Z, ZHANG G, LIU J, HU J, YAN M, DONG G, ZHU L, GUO L, QIAN Q, ZENG D. Map-based cloning proves qGC-6, a major QTL for gel consistency ofjaponica/indicacross, responds by Waxy in rice (Oryza sativaL.).Theoretical and Applied Genetics, 2011, 123(5): 859-867.
[33] GAO Z, ZENG D, CHENG F, TIAN Z. ALK, the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice.Journal of Integrative Plant Biology, 2011, 53(9): 756-765.
[34] NAKAMURA Y, FRANCISCO P B, HOSAKA Y, SATO A, SAWADA T, KUBO A, FUJITA N. Essential amino acids of starch synthase IIa differentiate amylopectin structure and starch quality betweenjaponicaandindicarice varieties.Plant Molecular Biology, 2005, 58(2): 213-227.
[35] LESTARI P, JIANG W, CHU S, REFLINUR, SUTRISNO, KUSBIANTORO B, KIM B, PIAO R, CHO Y, LUO Z, CHIN J, KOH H. DNA markers for eating quality ofindicarice in Indonesia.Plant Breeding, 2015, 134(1): 40-48.
(责任编辑 李莉)
Association Analysis of Rice Cooking Quality Traits with Molecular Markers
GAO WeiWei, CHEN SiPing, WANG LiPing, CHEN LiKai, GUO Tao, WANG Hui, CHEN ZhiQiang
(National Engineering Research Centre of Plant Space Breeding, South China Agricultural University, Guangzhou 510642)
【Objective】 In this study, the variation of major grain-quality traits and RVA profiles of representative rice germplasm were investigated, molecular markers significantly associated with the traits were identified, aiming to provide an important foundation for improvement of grain quality of rice. 【Method】 Global grain-quality properties and their differences were surveyed with a rice panel consisted of 48 diverse germplasms collected from both China and abroad. Starch RVA profile was examined with a Rapid Visco Analyzer. The markers reported to be related to starch biosynthesizing genes and QTLs for development of rice grain were used for genotyping. Trait-marker association for grain-quality properties was detected using the general linear model with Tassel 3.0 software. Moreover, stepwise regression analysis was performed with these detected markerssignificantly associated with grain-quality traits. The phenotypic effects of the alleles and allelic combinations were estimated and both elite alleles and typical carrier genotypes were identified. 【Result】There were wide phenotypic variance and diversity in amylose content (AC), gel consistency (GC) and alkali spreading value (ASV), and the coefficient of variation (CV.) ranged from 26.5% to 36.3%. Based on inspection of RVA profile, significant differences were found in breakdown (BDV), setback (SBV) and consistence (CSV), and these parameters have preferably reflected the diverse starch paste properties among different rice varieties. Correlation analysis showed the AC was positively correlated with cool paste viscosity (CPV), SBV and CSV, while negatively correlated with peak viscosity (PKV) and BDV. The GC was negatively correlated with SBV and CSV. A total of 491 alleles were identified with 154 polymorphic markers, with an average gene diversity of 0.447 and an average polymorphic information content of 0.390. Based on the analysis of trait-marker association, a total of 22 markers were detected to associate with grain-quality traits, which explained the phenotypic variance ranged from 14.11% to 75.62%. The GBSSI gene majorly affected the properties of AC and GC, and the SNP marker of Wx-G/T explained up to 61.44% and 41.87% of the phenotypic variation for AC and GC, respectively. While the SSIIa was the major gene affecting ASV, and the phenotypic variation of ASV explained by the markers of alk-GC/TT and SSIIa-F was up to 75.62% and 74.46%. The model equations based on stepwise regression analysis of AC, GC and ASV were developed using the significant markers, whose determination coefficients were 85.30%, 40.62%, and 80.38%, respectively.【Conclusion】The starch RVA profile wass closely related to AC, GC and ASV. The RVA profile can be used to evaluate rice quality traits more comprehensively. With trait-marker association analysis, 22 markers were detected to be associated with grain-quality traits, and five of these sites were associated with both AC and GC. Regression models showed allelic combination can produce different phenotypic effects.
rice; grain-quality traits; trait-marker association; allelic variation
10.3864/j.issn.0578-1752.2017.04.001
2016-08-22;接受日期:2016-10-17
国家高技术研究发展计划(2016YFD0102102)、广东省科技计划(2015B020231011)、国家现代农业产业技术体系建设专项(CARS-01-12)
联系方式:高维维,E-mail:18819266044@163.com。陈思平,E-mail:867716697@qq.com。高维维和陈思平为同等贡献作者。通信作者陈志强,E-mail:chenlin@scau.edu.cn。通信作者郭涛,E-mail:guoguot@scau.edu.cn
猜你喜欢
川北医学院学报(2022年6期)2022-06-24
美食(2022年5期)2022-05-07
早期教育(家庭教育)(2021年11期)2021-12-17
少儿科学周刊·儿童版(2021年21期)2021-12-11
智慧健康(2021年17期)2021-07-30
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
中国粮食经济(2018年5期)2018-12-27
现代园艺(2017年21期)2018-01-03
中国男科学杂志(2016年5期)2016-12-01
