
烤烟根系分泌物对植烟土壤酶及养分活化作用的影响
2017-03-30 05:38肖明礼韦建玉金亚波
广东农业科学 2017年12期
肖明礼,刘 高,韦建玉,金亚波,王 军,王 行,王 维
(1.广东中烟工业有限责任公司技术中心,广东 广州 510385;2.华南农业大学农学院,广东 广州 510642;3.广西中烟工业有限责任公司,广西 南宁 530001;4.广东省烟草南雄科学研究所,广东 南雄 512400)
根系是植物的主要吸收器官,向周围土壤环境释放多种代谢产物,包括糖类、蛋白质、氨基酸、维生素及有机酸、酚类等化合物,其中很多低分子量的化合物可作为化感物质影响植物根际土壤中微生物群落、土壤酶活性及土壤养分含量,同时影响根系与植物及其他有机体之间的信息传递。近年来,国内外学者对根系分泌物进行了大量研究,王晓英[1]研究了白三叶根系分泌物的化感作用,结果表明,白三叶根系分泌物可抑制高羊茅、草地早熟禾种子的萌发和幼苗生长,并认为这是白三叶能够成功入侵禾本科草坪的原因之一;孙燕等[2]研究了咖啡根系分泌物对嫁接后植物生长的影响,结果表明,根系分泌物可以促进植株生长或使植株生长所受抑制减少;徐国伟等[3]研究了秸秆还田和施氮量对水稻根系分泌物的影响,结果表明,根系分泌物与氮素积累量和氮素利用率呈显著正相关。周冀衡等[4]对不同基因型烤烟根系分泌物进行研究,结果表明香料烟(沙姆逊)根系分泌物的活化能力最强,白肋烟(白肋21)次之,烤烟(NC89)较弱。常二华等[5]研究表明,氮磷营养水平对水稻根系分泌物有重要的调控作用。宋日等[6]研究指出,不同作物根系分泌物对土壤团聚体大小及稳定性的影响具有重要作用,而土壤团聚体与土壤理化性质和生物学特性密切相关。可见,根系分泌物对植物生长发育至关重要。作物根系分泌物作为一个新兴研究热点已有大量研究,但有关烤烟根系分泌物对土壤结构、土壤养分及酶活性影响的研究则较少。本研究以广西河池产区植烟田为供试土壤,采用水培法收集烤烟根系分泌物,分析烤烟根系分泌物对土壤速效养分的影响,并通过对土壤酶活性的测定研究烤烟根系分泌物促进土壤速效养分释放的原因,为烤烟生产过程中土壤微生态深入研究提供参考。
1 材料和方法
1.1 试验材料
供试材料为广西河池市罗城县植烟土壤(沙泥田)0~20 cm耕作层的土壤,其基本理化性质为pH 5.72、有机质含量2.87%、全氮含量0.173%、全磷含量0.044%、全钾含量2.84%、速效氮含量89.32 mg/kg、速效磷含量18.37 mg/kg、速效钾含量124.65 mg/kg,土壤风干过筛后备用。采用托盘育苗法进行常规育苗,育苗营养液采用生产常规育苗肥料配方,待长到6~7叶期,剪叶一次,进行水培移栽,水培营养采用Hoagland's(霍格兰氏)营养液配方。待长到现蕾期,完整取出生长一致的烟株,分别放入1 L去离子水中培养,每隔24 h更换收集一次去离子水,连续收集2 d。将收集液减压浓缩至原收集液体积的10%(45℃),过0.45 μm滤膜,获得根系分泌物浓缩提取液。
1.2 试验方法
将根系分泌物浓缩提取液设置低(A1:5 mL分泌物+10 mL去离子水))、中(A2:10 mL分泌物+5 mL去离子水)、高(A3:15 mL分泌物)3个浓度。取50 g土样于培养皿中,将上述溶液均匀点滴入土样中,各处理重复10次,置于25℃培养箱黑暗培养,每天称重,用去离子水补足损失水分。培养5、15、30 d后,取土样分析。
1.3 测定指标及方法
土壤水稳性团聚体测定采用非真空快速浸润法[7],碱解氮含量测定采用凯氏定氮法、速效磷含量测定采用碳酸氢钠浸提为钼锑抗比色法)、速效钾含量测定采用中性乙酸铵提取为火焰光度计法)、有机质含量测定采用硫酸重铬酸钾容量法[8];蔗糖转化酶、磷酸酶、过氧化氢酶、多酚氧化酶活性测定采用荧光微型板酶检测技术,脲酶活性测定采用NH+4释放量法[9]。
2 结果与分析
2.1 烤烟根系分泌物对不同粒径水稳性团聚体组成含量的影响
水稳性团聚体组成是影响土壤理化性质及生物学性质的重要因素,对土壤肥力及生产力至关重要。由表1可知,不同浓度烤烟根系分泌物处理下,土壤水稳性团聚体均主要集中于大团聚体(粒级>1 mm)和小团聚体(粒级<0.1 mm),且整个培养过程中,不同根系分泌物处理的土壤小团聚体比例先降后升,大团聚体比例则先升后降。整体来看,土壤水稳性小团聚体和大团聚体分布比例均升高,且大团聚体分布比例高于小团聚体,其中各处理小团聚体在培养过程中的升高比例分别为13.8%、19.8%、12.8%,而大团聚体升高比例分别为38.4%、27.3%、30.7%,可见烤烟根系分泌物可显著增加土壤中大团聚体的数量,0.1~1.0 mm粒级的土壤水稳性团聚体分布比例降低。不同培养时间下,随着根系分泌物浓度的增加,小团聚体含量减少,大团聚体含量增加,0.1~1.0 mm粒级土壤水稳性团聚体未表现明显变化规律。说明,烤烟根系分泌物有利于微团聚体团聚成大团聚体,减少烤烟生长过程中栽培技术措施对土壤水稳性团聚体的破坏。
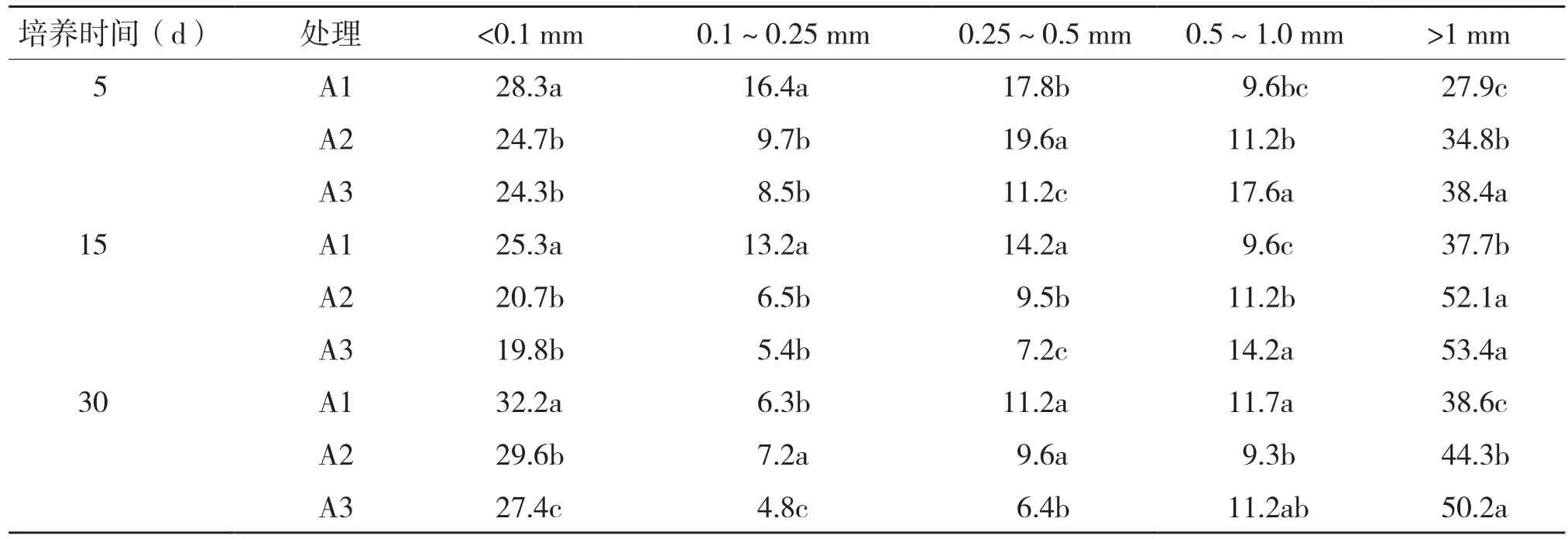
表1 根系分泌物对不同粒径水稳性团聚体含量(%)的影响
2.2 烤烟根系分泌物对土壤养分含量的影响
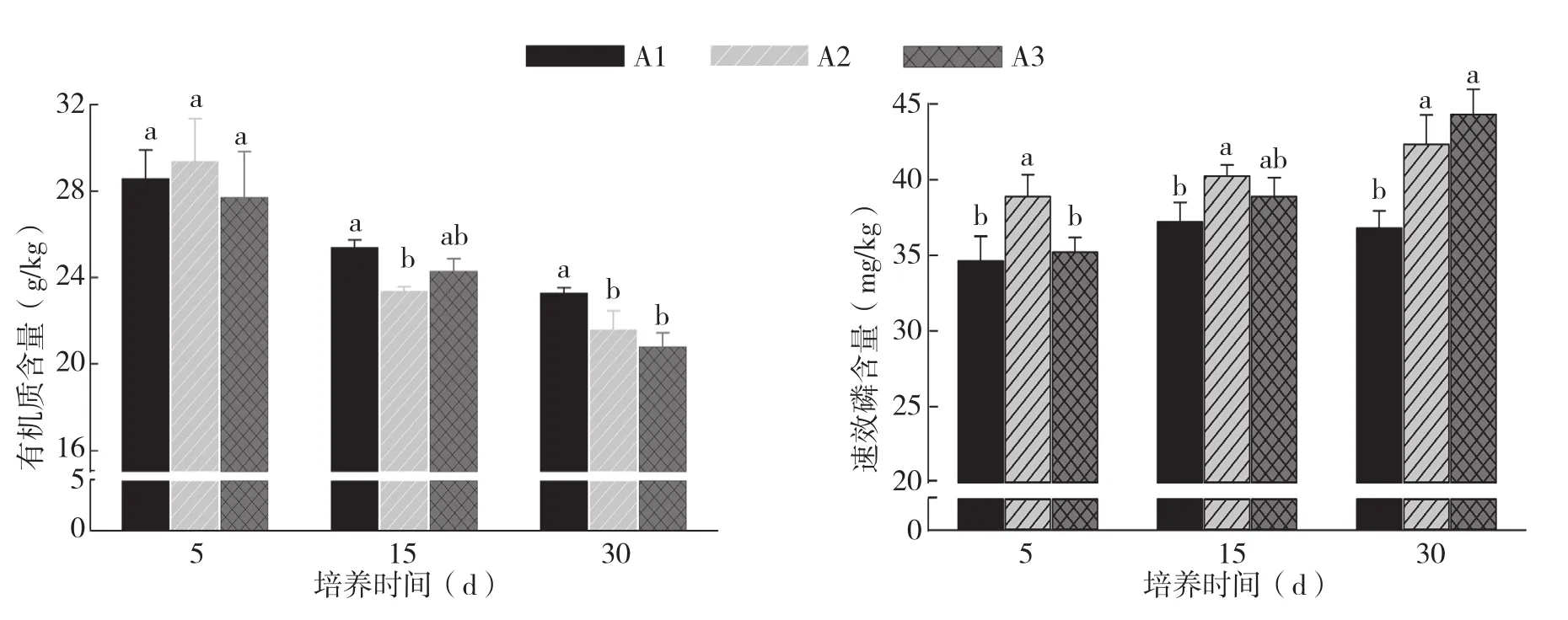
图1 烤烟根系分泌物对土壤有机质、速效磷含量的影响
由图1可知,添加根系分泌物后,随着培养时间增加,土壤中有机质含量下降。培养5 d后不同浓度处理土壤有机质含量差异不显著。但培养30 d后,A1处理土壤有机质含量显著高于A2、A3处理,而A2、A3处理间差异不显著。
由图1、图2可知,添加根系分泌物后,随着培养时间增加,土壤速效磷、速效钾、速效氮含量增加。其中,培养5 d后,A2处理土壤速效磷含量明显高于A1、A3处理,培养15、30 d后,A2、A3处理的速效磷含量均高于A1处理,而A2、A1处理间差异不显著。不同浓度处理土壤速效钾含量在培养5 d及培养15 d的变化趋势和速效磷趋于一致,但在培养30 d后,A3处理土壤速效钾含量与A1、A2处理相比显著提高。培养5、15 d后,不同浓度处理间土壤速效氮含量差异不显著,但培养30 d后,A2、A3处理显著高于A1处理,A2、A3处理间差异不显著。综合分析认为,不同浓度的根系分泌物对土壤养分含量有明显影响,A3处理烤烟根系分泌物更有利于土壤中速效磷、速效钾及速效氮的释放。而A1处理烤烟根系分泌物则更有利于土壤有机质含量增加。
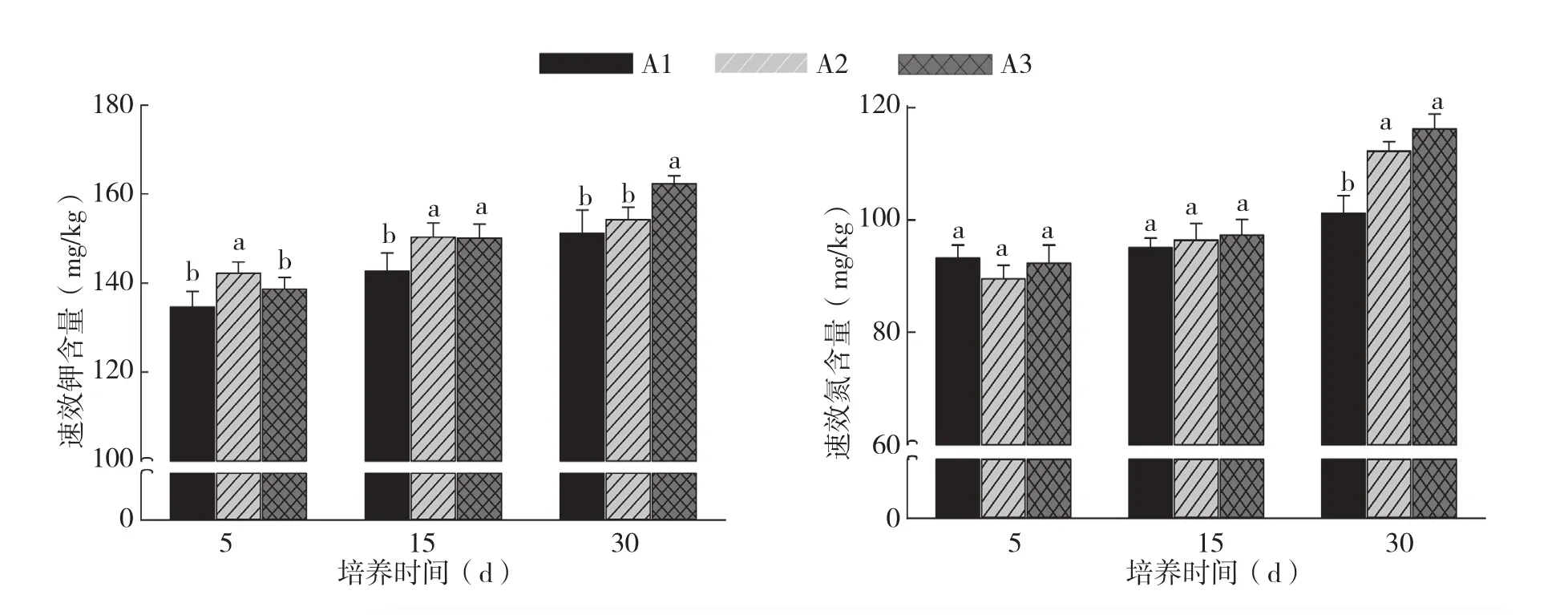
图2 烤烟根系分泌物对土壤速效氮、速效钾含量的影响
2.3 烤烟根系分泌物对土壤酶活性的影响
由图3~图5可知,烤烟根系分泌物对土壤酶活性有一定影响。培养30 d后,A3处理土壤过氧化氢酶活性比A1处理提高9.4%,达到显著水平。而在培养15 d前,各浓度处理间差异不显著。各处理土壤的多酚氧化酶活性在培养15 d后达到最大值,且随着根系分泌物浓度的增加土壤多酚氧化酶的活性增加,处理间差异显著。培养30 d后,A3处理土壤多酚氧化酶活性显著高于A1、A2处理。
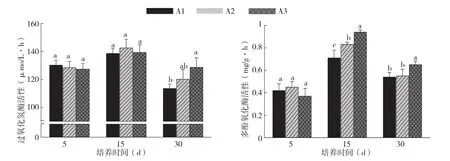
图3 烤烟根系分泌物对土壤过氧化氢酶、多酚氧化酶活性的影响
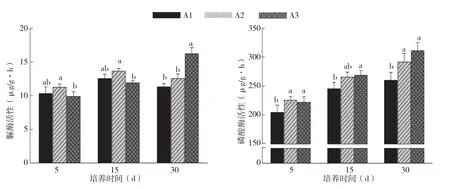
图4 烤烟根系分泌物对土壤过脲酶、磷酸酶活性的影响
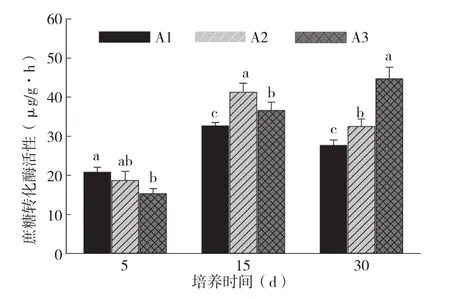
图5 烤烟根系分泌物对土壤蔗糖转化酶活性的影响
A1、A2处理土壤脲酶活性差异不显著,且在培养15 d前,A2处理土壤脲酶活性显著高于A3处理,而在培养30 d后,A3处理反而高出A2处理20.8%。A3处理土壤磷酸酶活性显著高于A1处理,但与A2处理差异不显著。各处理土壤磷酸酶活性随培养时间的增加而增加。不同浓度根系分泌物的土壤蔗糖转化酶活性在不同的培养时间下差异较大。培养5 d后,A1处理显著高于A3处理,培养15 d后,土壤蔗糖转化酶活性最高的为A2处理,而在培养30 d后,土壤蔗糖转化酶活性最高的为A3处理,并分别比A2、A1处理提高35.3%、53.3%。
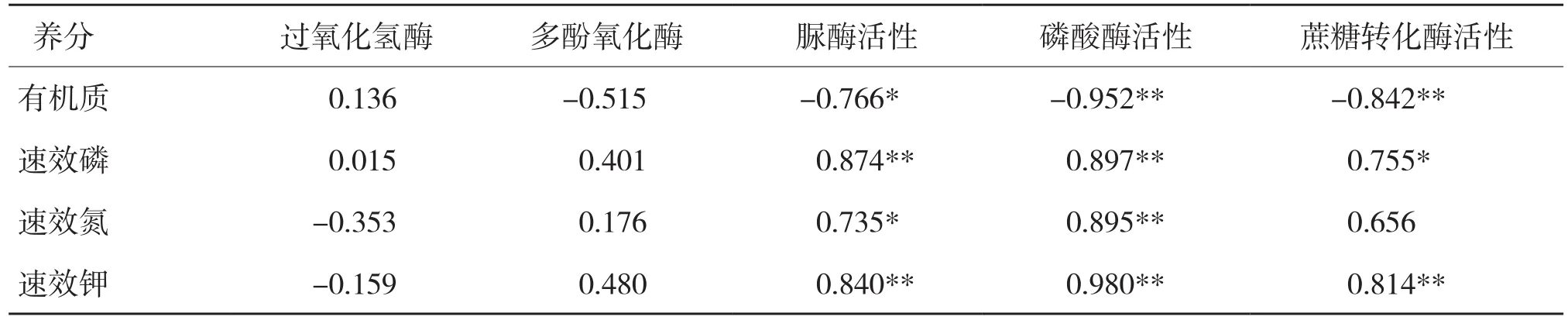
表2 土壤养分含量与土壤酶活性的相关分析
2.4 土壤养分含量与土壤酶活性的相关分析
从表2可以看出,过氧化氢酶和多酚氧化酶活性与土壤养分含量的相关系数较小,而脲酶活性、磷酸酶活性、蔗糖转化酶活性与土壤有机质、速效磷、速效氮、速效钾的含量关系密切。其中,脲酶活性、磷酸酶活性、蔗糖转化酶活性与土壤有机质含量呈显著负相关,说明脲酶活性、磷酸酶活性、蔗糖转化酶活性的增加有助于土壤有机质的转化。脲酶活性与土壤速效磷、速效钾的含量呈极显著正相关,与速效氮含量呈显著正相关。此外,磷酸酶活性与速效磷、速效氮、速效钾的含量呈极显著正相关,相关系数分别为0.897、0.895、0.980。而蔗糖转化酶与土壤速效钾和速效磷的含量也呈显著正相关。可见,土壤酶直接参与土壤中物质的转化及养分的释放和固定,其活性与土壤肥力状况密切相关。
3 结论与讨论
土壤团聚体是指土粒通过各种自然过程的作用而形成的直径<10 mm的结构单位,根系分泌物是影响土壤团聚体大小和数量的主要因素之一[10-12]。宋日等[13]研究表明,大豆根系分泌物可显著增加土壤中水稳性团聚体的含量。本研究指出,根系分泌物可有效增加土壤团聚体数量。土壤小团聚体含量先减后增,土壤大团聚体含量先增后减,但是整体来看,土壤大团聚体的增加幅度大于土壤小团聚体的增加幅度,所以不同浓度根系分泌物均能显著增加土壤大团聚体含量。在烤烟的生产过程中,大田起垄措施及二次培土都会很大程度的破坏土壤结构,根系分泌物可以在一定程度上修复这种破坏。
有机质含量是判别土壤活性的重要指标之一,有机质的分解、积累及平衡是保持和提高土壤肥力的基础[14],植烟土壤有机质含量的高低对烟草的生长发育及产质量有直接的影响[15-21]。耿贵[22]研究表明,玉米、大豆、甜菜3种作物的根系分泌物对土壤有机质含量影响不明显。本研究表明,不同浓度根系分泌物均能有效降低土壤有机质含量,且浓度越大有机质含量越低,可能是因为烤烟根系分泌物可以增强根系分泌酶活性,而根系分泌酶在土壤有机质转化过程中起重要作用,根系分泌酶活性增强,有机质转化、分解加快。土壤中的氮、磷、钾是作物生长发育必不可缺的营养元素,作物根系分泌物中也含有少量的氮,但因为土壤中氮含量较多,并不会明显增加土壤中全氮含量,但会改变土壤中活性氮的含量和比例[23],本研究表明烤烟根系分泌物可以增加土壤中速效磷、速效钾、速效氮含量,且随根系分泌物浓度的增加而增加,所以烤烟根系分泌物可以通过提高土壤中速效磷、速效钾、速效氮的含量和比例,进而影响烤烟的生长发育。
土壤酶对作物的生长发育至关重要,主要来源于动植物残体、土壤微生物及根系分泌物[24],其活性可以反映土壤内生化反应的方向和强度。土壤过氧化氢酶是一种重要的土壤微生态环境指标,能有效防止作物过氧化氢毒害[25];土壤多酚氧化酶是一种复合酶,常用于土壤环境修复;脲酶在土壤氮素转化过程中起着重要作用,其活性在一定程度上能够反映土壤供氮的能力,可以作为土壤生化性质的评价指标之一[26-27];土壤磷酸酶在土壤磷素循环中发挥着重要作用,其活性高低直接影响着土壤中有机磷的生物化学矿化过程和有效磷的供给水平;土壤转化酶的强弱可以直接反映了土壤熟化程度和肥力的水平,其活性高低可作为评价土壤肥力的指标[28-29],在反映土壤肥沃状况的同时比其他酶更为明显[30],土壤肥力高的转化酶活性相应就高。本试验结果表明,根系分泌物可有效增强土壤过氧化氢酶、多酚氧化酶、脲酶、磷酸酶、蔗糖转化酶活性。烤烟根系分泌物可显著影响土壤过氧化氢酶活性,培养前期3种浓度处理差异不显著,培养30 d时,过氧化氢酶活性随根系分泌物浓度的增加而增加,并且需要一定的时间才能更好的发挥作用。培养15 d时,不同浓度根系分泌物可显著影响土壤多酚氧化酶活性,整个培养过程中酶活性先增后降,说明根系分泌物对多酚氧化酶活性的影响有一定时效性。从整个培养过程来看,只有高浓度处理下,脲酶活性是一直增加的,这说明高浓度的根系分泌物对土壤脲酶的影响最大。不同浓度根系分泌物处理对磷酸酶活性的影响均随培养时间的增加而增加,整个培养过程中、高浓度处理差异性均不显著,这两个处理与低浓度处理差异显著。中低浓度根系分泌物随培养时间增加对蔗糖转化酶活性的影响均是先增后减,高浓度根系分泌物对蔗糖转化酶活性的影响则是随培养时间的增加而增加,在培养天数为30 d时,3种处理蔗糖转化酶活性差异显著,说明高浓度的根系分泌物对蔗糖转化酶活性的影响最大且时效较长。
[1]王晓英. 白三叶根系分泌物的化感作用及其GC-MS分析[J]. 河南农业科学,2016,45(5):96-100,134.
[2]孙燕,杨建峰,等. 咖啡根系分泌物对嫁接后植株生长及叶片保护酶活性的影响[J]. 生态学杂志,2016,36(5):1310-1314.
[3]徐国伟,李帅,赵永芳,等. 秸秆还田与施氮对水稻根系分泌物及氮素利用的影响研究[J].草业学报,2014,23(2):140-146.
[4]周冀衡,李永平,杨虹琦,等. 不同基因型烟草根系分泌物对难溶性磷钾的活化效应[J].湖南农业大学学报(自然科学版),2005,31(3):276-280.
[5]常二华,张耗,张慎凤,等. 结实期氮磷营养水平对稻根系分泌物的影响及其与稻米品质的关系[J]. 作物学报,2007,33(12):1949-1959.
[6]宋日,刘利,马丽艳,等. 作物根系分泌物对土壤团聚体大小及其稳定性的影响[J]. 南京农业大学学报,2009,32(3):93-97.
[7]安婉丽,高灯州,潘婷,等. 水稻秸秆还田对福州平原稻田土壤水稳性团聚体分布及稳定性影响[J]. 环境科学学报,2016,36(5):1833-1840.
[8]盛茂银,熊康宁,崔高仰,等. 贵州喀斯特石漠化地区植物多样性与土壤理化性质[J]. 生态学报,2015,35(2):434-448.
[9]荣勤雷,梁国庆,周卫,等. 不同有机肥对黄泥田土壤培肥效果及土壤酶活性的影响[J]. 植物营养与肥料学报,2014,20(5):1168-1177.
[10]Zhang M K,He Z L,Chen G C,et. al. Formation and water stability of aggregates in red soil as affected by organic matter [J]. PEDOSPHERE,1996,6(1):39-45
[11]张知贵,王明贤,杨华,等. 植物根系分泌物研究进展[J]. 广东农业科学,2013,40(2):219-222.
[12]Marschner P,Rengel Z. Contributions of Rhizosphere Interactions to Soil Biological Fertility[M]. L K. Aboutt and Murphy(eds):Soil Biological Fertility-A Key to suatainable Land Use in Agriculture,2003.
[13]宋日,刘利,马丽艳,等. 大豆根系分泌物对土壤团聚体大小和稳定性的影响[J]. 东北林业大学学报,2009,37(7):84-86.
[14]马敬,罗世琼,杨占南,等. 鱼腥草土壤养分与微生物特征分析[J]. 广东农业科学,2015,42(19):55-62.
[15]陈江华,李志宏,刘建利,等. 全国主要烟区土壤养分丰缺状况评价[J]. 中国烟草学报,2004,11(3):14-18.
[16]孙燕,高焕梅,和林涛. 土壤有机质及有机肥对烟草品质的影响[J]. 安徽农业科学,2007,35(20):6160-6161.
[17]王怡,陈建军,李福君,等. 广东南雄烟区主要植烟土壤类型养分状况分析[J]. 广东农业科学,2014,41(1):37-41.
[18]Tso T C. Production,physiology,and biochemistry of tobacco plant[J]. Production Physiology & Biochemistry of Tobacco Plant,1990.
[19]邓小华,谢鹏飞,彭新辉,等. 土壤和气候及其互作对湖南烤烟部分中性挥发性香气物质含量的影响[J]. 应用生态学报,2010,21(8):2063-2071.
[20]彭新辉,邓小华,易建华,等. 气候和土壤及其互作对烟叶物理性状的影响[J]. 烟草科技,2010(2):48-58.
[21]何承刚,张金华,苏德艳,等. 保山烤烟总糖含量与植烟土壤有机质含量的分布特征及关系分析[J]. 云南农业大学学报,2008,23(6):832-835.
[22]耿贵. 作物根系分泌物对土壤碳、氮含量、微生物数量和酶活性的影响[D]. 沈阳:沈阳农业大学,2011.
[23]杨万勤,王开运. 土壤酶研究动态与展望[J].应用与环境生物学报,2002,8(5):564-570.
[24]马云华,王秀峰,魏岷,等. 黄瓜连作土壤酚类物质积累对土壤微生物和酶活性的影响[J].应用生态学报,2005,16(11):2149-2153.
[25]Woese C R. Bacterial evolution background[J].Microbiological Reviews,1987,51:221-271.
[26]黄世伟. 土壤酶活性与土壤肥力[J]. 土壤通报,1981(4):37-39.
[27]和文祥,朱铭莪. 陕西土壤脲酶活性与土壤肥力关系分析[J]. 土壤学报,1997,34(4):392-398.
[28]李萍,徐雅梅. 不同培肥措施对藏东南土壤酶活性的影响[J]. 土壤肥料,2002(5):33-35.
[29]赵哲权,王明九,刑建民. 施用作物秸杆对土壤酶活性的影响[J]. 土壤肥料,1990(3):28-29.
[30]熊明彪,田应兵,雷孝章,等. 小麦生长期内土壤养分与土壤酶活性变化及其相关性研究[J]. 水土保持学报,2003(4):27-30.
猜你喜欢
东南大学学报(自然科学版)(2022年4期)2022-08-12
食品与发酵工业(2021年9期)2021-05-21
腐植酸(2020年1期)2020-11-29
伴侣(2019年5期)2019-05-27
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
现代农村科技(2018年7期)2018-08-06
现代园艺(2017年13期)2018-01-19
现代园艺(2017年21期)2018-01-03
人人健康(2017年23期)2017-12-26
绿色科技(2016年16期)2016-10-11
