
白眉姬鹟身体条件对繁殖投入的影响
2017-03-24 06:45:47刘珍妮姚纪元张立世赵殿郑姜云垒王海涛
东北师大学报(自然科学版) 2017年1期
刘珍妮,姚纪元,张立世,赵殿郑,姜云垒,王海涛
(1.吉林农业大学动物科学与技术学院,吉林 长春 130118;2.东北师范大学生命科学学院,吉林 长春 130024)
白眉姬鹟身体条件对繁殖投入的影响
刘珍妮1,姚纪元1,张立世1,赵殿郑1,姜云垒1,王海涛2
(1.吉林农业大学动物科学与技术学院,吉林 长春 130118;2.东北师范大学生命科学学院,吉林 长春 130024)
在吉林省左家自然保护区山地次生林中,利用人工巢箱招引白眉姬鹟,进行了其身体条件对繁殖投入的影响研究.结果表明:白眉姬鹟雌鸟身体条件与首枚卵产期、窝卵数及巢材重量投入无显著相关性;雄鸟身体条件与首枚卵产期呈显著负相关(r=-0.525,P<0.001),与窝卵数呈显著正相关(r=0.312,P=0.029);雌鸟身体条件与孵化期坐巢时间呈显著正相关(r=0.367,P=0.013);雌雄鸟身体条件与递食次数无显著相关性,但亲鸟育雏次数随窝雏数增加而显著增加(r=0.552,P<0.001).研究表明白眉姬鹟可根据自身和配偶身体条件调整繁殖投入.
白眉姬鹟;身体条件;繁殖投入
广义的鸟类繁殖投入是对配对、筑巢、产卵、孵化、育幼直至幼鸟独立前的所有繁殖行为的投入[1-2].繁殖投入主要体现在生产子代以及抚育和管护时所消耗的能量、时间和资源量[3].身体条件代表了个体的营养水平、健康状况及生存经验,身体条件可以真实地反映表型质量[4];很多实验指出鸟类身体条件与窝卵数、亲鸟抚育、成活率及繁殖力等因素呈正相关[5-6],所以个体身体条件的差异可能导致繁殖投入的不同,亲鸟可能根据自身条件调整繁殖对策并做出权衡,达到繁殖成效的最大化[7-8].
白眉姬鹟(Ficedulazanthopygia)隶属于雀形目(Passeriformes)、鹟科(Muscicapidae)、姬鹟属(Ficedula),无亚种分化,繁殖期为5—7月,在我国东北、华北、华东、华中以及俄罗斯远东地区繁殖,在东南亚越冬.[9]国外对该鸟未见研究报道;国内研究主要集中于食性、人工巢箱条件下的繁殖参数、巢址选择、分子系统发育等方面[10-12].本文对白眉姬鹟身体条件对繁殖期各方面投入的影响进行了初步分析,比较了各时期投入的差异,探讨了白眉姬鹟的身体条件是否影响各时期的投入,以为雀形目鸟类繁殖投入与形态进化的关系研究提供基础资料.
1 研究区域概况
研究区位于吉林左家自然保护区,该保护区为吉林省东部森林和西部平原的过渡地带,地理位置为北纬44°01~44°06、东经126°00~126°01′,海拔200~530 m.地区内森林以天然次生阔叶林为主,乔木主要有蒙古栎(Quercusmongolicus)、糠椴(Tiliamandshurica)、黑桦(Betuladahurica)、水曲柳(Fraxinusmandschurica)、春榆(Ulmusjaponica)、黄檗(Phellodendromamurense)等;灌木主要有榛子(Corylusheterophylla)、刺玫(Rosedavurica)等;草本植物以菊科(Asteraceae)和蓼科(Polygonaceae)植物为主.保护区内共记录到鸟类190种,优势种包括大山雀(Parusmajor)、普通(Sittaeuropaea)、白眉姬鹟(Ficedulazanthopygia)、喜鹊(Picapica)等[13].
2 研究方法
2014—2015年3—7月,以保护区山地次生林为研究区域,选择生境结构基本相似,且植被生长良好的9块林地为研究样地.3月中旬在样地悬挂人工巢箱,人工巢箱采用内部规格为25 cm×12 cm×12 cm的山雀式木板箱,洞口直径为4 cm,巢箱开口在前壁上方1/3处.悬挂树种随机选择,悬挂高度3~4 m,巢箱间距为50 m左右.研究区域内白眉姬鹟于5月中旬开始繁殖,白眉姬鹟进入繁殖期前,每隔2日检查一遍巢箱是否被白眉姬鹟占用,确定被利用的标准为巢箱中是否出现巢材或卵,如巢箱被利用,则每天进行观察以确定繁殖参数.记录首枚卵产期、窝卵数、出雏数和出飞数.在孵化期第6天及育雏期第7天采用录像和直接观察的方法记录白眉姬鹟的行为活动,每天观察时间8:00—16:00.录像时选择晴朗天气.在室内对录像进行回放观察,记录亲鸟坐巢时间和育雏次数.在育雏期中后期,利用挡板法捕捉雌雄亲鸟,利用游标卡尺和电子天平测量雌雄鸟的喙长、翅长、尾长、跗跖长、体重等身体指标.雏鸟出飞后收集巢材称重.
统计分析时,利用雌雄鸟体重与翅长、跗跖长的主成分得分回归残差作为雌雄鸟身体条件指标[14],利用单样本t检验验证雌雄鸟不同个体之间的身体特征和身体条件是否存在显著差异,通过单样本的Kolmogorov-Smirnov检验判定各组数据是否符合正态分布.本文数据符合正态分布,则选择双尾Pearson相关分析方法分析白眉姬鹟雌雄鸟身体条件与首枚卵产期、窝卵数及递食次数的关系,分析时将首枚卵产期转换为Julian日期格式数值后与成鸟身体条件做相关性分析.由于雌鸟单独筑巢并单独孵化,所以仅采用Pearson相关分析方法分析雌鸟身体条件与巢材重量和坐巢时间的关系.以上数据分析均用SPSS21.0软件完成,显著性水平P<0.05.
3 结果
3.1 白眉姬鹟不同个体身体特征比较
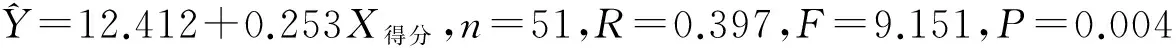
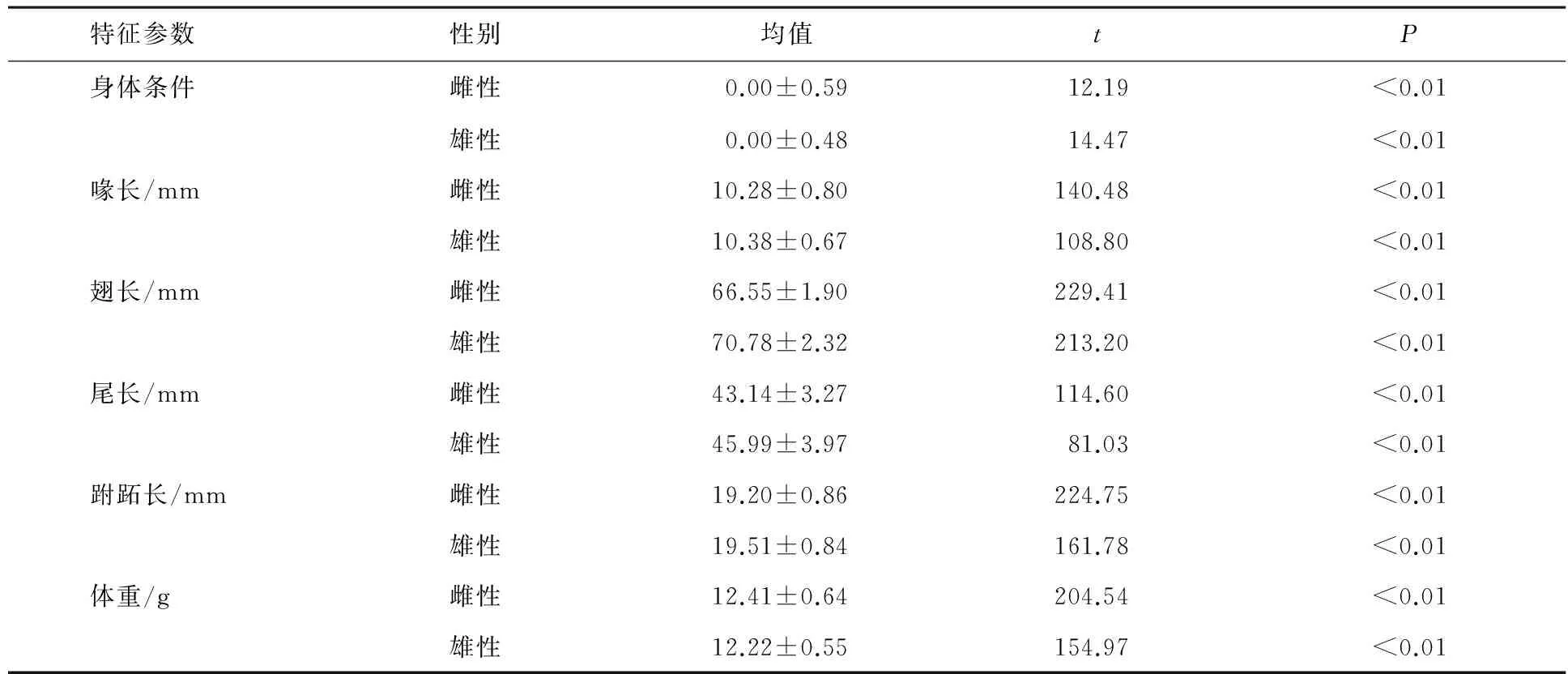
表1 白眉姬鹟雌雄鸟形态特征参数

表2 雌鸟主成分分析结果

表3 雄鸟主成分分析结果
3.2 白眉姬鹟身体条件对繁殖投入的影响
白眉姬鹟雌鸟身体条件与窝卵数和首枚卵产期均无显著相关性;雄鸟身体条件与首枚卵产期显著负相关,与窝卵数显著正相关(见表4).白眉姬鹟雌鸟身体条件与巢材重量无显著相关性,与孵化期坐巢时间显著正相关(见表4).白眉姬鹟雌鸟身体条件与递食次数无显著相关性(Pearson相关:r=-0.019,P=0.899,n=47);白眉姬鹟雄鸟身体条件与递食次数无显著相关性(Pearson相关:r=0.187,P=0.208,n=47).亲鸟的递食数与雏鸟数显著正相关(Pearson相关:r=0.552,P<0.001,n=46).
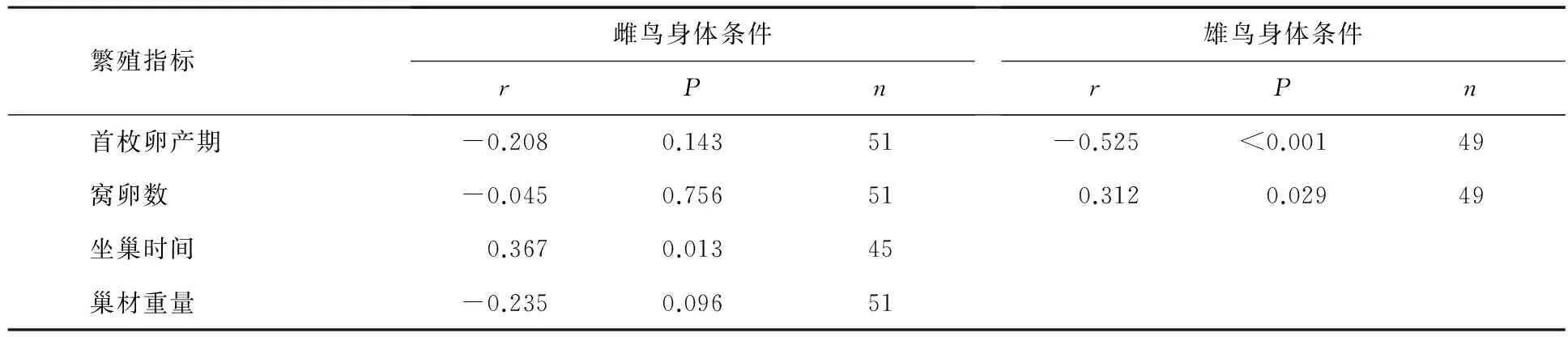
表4 白眉姬鹟雌雄鸟身体条件与首枚卵产期、窝卵数、巢材重量及坐巢时间的关系
4 讨论
4.1 身体条件对筑巢投入的影响
巢材的运输反映了亲鸟对筑巢活动的投资,是繁殖投入的重要组成部分[15-16].筑巢投入主要与筑巢期间的被捕食风险和在获取巢材过程中的能量消耗有关,有研究表明身体条件好的个体有更重的巢材[17-18].Toms等人[19]报道了蓝山雀(Cyanistescaeruleus)巢重和雌鸟身体条件之间有正相关关系.白眉姬鹟由雌性筑巢,本研究中未发现白眉姬鹟雌鸟身体条件和巢材重量的相关性.白眉姬鹟为夏候鸟,繁殖时间有限,筑巢期较短,在观察中发现有筑巢仅一天就产卵的现象,还有利用大山雀旧巢产卵的现象,雌鸟可能将有限的时间和能量用于繁殖的其他阶段,所以白眉姬鹟身体条件的差异并不能反映巢材重量的差异.
4.2 身体条件对产卵投入的影响
鸟类繁殖时间的选择是其在自然选择条件下的一种适应,常与食物丰富度有关[20].由于身体条件能直接反映其储备的能量,间接反映食物的丰富度,所以许多研究者提出繁殖时间可能受到身体条件的影响[21-23].对白眉姬鹟的研究中未发现雌鸟身体条件与首枚卵产期有相关性,但雄鸟身体条件与首枚卵产期显著负相关.白眉姬鹟为夏候鸟,身体条件好的雄鸟可能更早到达繁殖地,并且在争夺配偶、抢占领地时有一定优势[24],能较早获得繁殖的机会.
鸟类对产卵的投入受个体资源、时间和能量的限制,两性育雏鸟类中,环境质量[25],雌鸟身体条件[5]和雄鸟身体条件[26]三个因素可能会影响窝卵数.本研究发现白眉姬鹟窝卵数与雄鸟身体条件呈正相关,雌鸟可能根据雄鸟的身体条件调整窝卵数.雄鸟身体条件好,雌鸟从雄鸟那里不仅可以得到一些直接的益处,如优质食物和隐蔽场所等,而且还会得到一些间接的益处,如后代会获得雄鸟优质的基因,这些直接和间接的益处使雌鸟承担的繁殖风险较小,从而增加了窝卵数.
4.3 身体条件对孵化投入的影响
鸟类的孵化行为对雏鸟在卵中的发育起重要作用,直接影响鸟类的繁殖成功情况[27].亲鸟坐巢时间必然在满足自身基本能量需求与保持卵温使雏鸟胚胎正常发育之间做权衡[28].白眉姬鹟由雌鸟单独孵化,与两性孵卵的鸟类相比,单独孵化的雌鸟需要承担自我能量需要、孵化时间分配和自我保护等多项责任[29].本研究结果表明白眉姬鹟雌鸟身体条件与孵化期坐巢时间呈显著正相关,这说明身体条件好的雌鸟可能有更多的能量储备,从而相应减少外出觅食时间,把更多的时间和能量用于孵卵.鸟类孵化期的能量分配取决于亲鸟的能量储备,身体条件良好的个体比身体条件差的投入更多,类似的关系在斑姬鹟(Ficedulahypoleuca)中也有报道[30].
4.4 身体条件对育雏投入的影响
鸟类身体条件可以反映觅食能力,身体条件好的亲鸟觅食能力强、递食率高,可能会给雏鸟提供更多的营养物质促进其生长[31-32].但本研究未发现亲鸟身体条件与递食率之间的相关性,可能因为递食率还受到食物丰富度、环境因素的影响[33].亲鸟递食率通常随后代的数量增加而增加,本研究结果支持窝雏数假说[34].白眉姬鹟亲鸟育雏次数与窝雏数呈正相关,与魏忠民等[35]的研究结果一致.
[1] TRIVERS R.Parental investment and sexual selection[M].Harvard:Biological Laboratories,Harvard University,1972.
[2] PARISH D M B,COULSON J C.Parental investment,reproductive success and polygyny in the lapwing,Vanellusvanellus[J].Animal Behaviour,1998,56(4):1161-1167.
[3] 张晓爱,赵亮,胥志清.鸟类生态能量学的几个基本问题[J].动物学研究,2001,22(3):231-238.
[4] LEHIKOINEN A,JAATINEN K,ÖST M.Do female ornaments indicate quality in eider ducks [J].Biology Letters,2010,6(2):225-228.
[5] GLADBACH A,GLADBACH D J,QUILLFELDT P.Seasonal clutch size decline and individual variation in the timing of breeding are related to female body condition in a non-migratory species,the Upland GooseChloephagapictaleucoptera[J].Journal of Ornithology,2010,151(4):817-825.
[6] MASELLO J F,QUILLFELDT P.Body size,body condition and ornamental feathers of Burrowing Parrots:variation between years and sexes,assortative mating and influences on breeding success [J].Emu,2003,103(2):149-161.
[7] HANSSEN S A,ERIKSTAD K E,JOHNSEN V,et al.Differential investment and costs during avian incubation determined by individual quality:an experimental study of the common eider(Somateriamollissima)[J].Proceedings of the Royal Society of London B:Biological Sciences,2003,270(1514):531-537.
[8] 陶慧娟,姜云垒,秦博,等.栗斑腹鹀双亲递食率比较研究[J].东北师大学报(自然科学版),2014,46(3):105-108.
[9] 郑光美.中国鸟类分类与分布名录[M].北京:科学出版社,2011.
[10] 张维,王海涛,杨志杰.人工巢箱条件下白眉姬鹟的繁殖参数[J].动物学杂志,2008,43(6):123-126.
[11] 曹长雷,高玮,由玉岩,等.人工巢箱条件下白眉姬鹟的繁殖习性和巢址选择[J].生态学杂志,2010(6):1193-1197.
[12] 雷忻,廉振民,雷富民,等.基于线粒体基因cytb的鹟亚科部分鸟类系统发育[J].动物学报,2007,53(1):95-105.
[13] 孙寒梅,高玮,宫亮,等.吉林省左家自然保护区鸟类组成的多样性研究[J].东北师大学报(自然科学版),2008,40(1):100-110.
[14] POWELL L L,DOBBS R C,MARRA P P.Habitat and body condition influence American Redstart foraging behavior during the non-breeding season [J].Journal of Field Ornithology,2015,86(3):229-237.
[15] LOMBARDO M P.Nest architecture and reproductive performance in Tree Swallows(Tachycinetabicolor) [J].The Auk,1994:814-824.
[16] 张晓爱,刘泽华,赵亮,等.青藏高原常见雀形目鸟类的筑巢特征[J].动物学研究,2006,27(2):113-120.
[17] LENS L,WAUTERS L A,DHONDT A A.Nest-building by crested titParuscristatusmales:an analysis of costs and benefits [J].Behavioral Ecology and Sociobiology,1994,35(6):431-436.
[18] DE NEVE L,SOLER J J.Nest-building activity and laying date influence female reproductive investment in magpies:an experimental study [J].Animal Behaviour,2002,63(5):975-980.
[20] WALSBERG G E.Avian ecological energetic [J].Avian Biology,1983,7:161-220.
[21] LASIEWSKI R C,DAWSON W R.Are-examination of the relation between standard metabolic rate and body weight in birds [J].The Condor,1967,69(1):13-23.
[22] MURPHY M T.Body size and condition,timing of breeding,and aspects of egg production in Eastern Kingbirds [J].The Auk,1986:465-476.
[23] WIGGINS D A.Natural selection on body size and laying date in the tree swallow [J].Evolution,1991:1169-1174.
[24] DUVAL E H,KEMPENAERS B.Sexual selection in a lekking bird:the relative opportunity for selection by female choice and male competition [J].Proceedings of the Royal Society of London B:Biological Sciences,2008,275(1646):1995-2003.
[25] ROBINSON T J,SIEFFERMAN L,RISCH T S.Seasonal tradeoffs in reproductive investment in a multi-brooded passerine [J].The Condor,2010,112(2):390-398.
[26] VELANDO A,ALONSO-ALVAREZ C.Differential body condition regulation by males and females in response to experimental manipulations of brood size and parental effort in the blue-footed booby[J].Journal of Animal Ecology,2003,72(5):846-856.
[27] 汪青雄,肖红,杨超,等.陕西红碱淖棕头鸥孵卵行为[J].生态学杂志,2015,34(3):760-764.
[28] 赵亮,张晓爱,李来兴.角百灵和小云雀的孵化行为[J].动物学报,2002,48(5):695-699.
[29] CRESSWELL W,HOLT S,REID J M,et al.The energetic costs of egg heating constrain incubation attendance but do not determine daily energy expenditure in the pectoral sandpiper [J].Behavioral Ecology,2004,15(3):498-507.
[30] HILLSTROM L.Body mass reduction during reproduction in the Pied FlycatcherFicedulahypoleuca:physiological stress or adaptation for lowered costs of Locomotor[J].Functional Ecology,1995:807-817.
[31] COOCH E,LANK D,DZUBIN A,et al.Body size variation in lesser snow geese:environmental plasticity in gosling growth rates [J].Ecology,1991:503-512.
[32] SAINO N,CALZA S,MOLLER A P.Immunocompetence of nestling barn swallows in relation to brood size and parental effort [J].Journal of Animal Ecology,1997:827-836.
[33] TREMBLAY I,THOMAS D,BLONDEL J,et al.The effect of habitat quality on foraging patterns,provisioning rate and nestling growth in Corsican Blue TitsParuscaeruleus[J].Ibis,2005,147(1):17-24.
[34] SLAGSVOLD T.Clutch size variation of birds in relation to nest predation:on the cost of reproduction [J].The Journal of Animal Ecology,1984:945-953.
[35] 魏忠民,姜云垒,张立世,等.人工巢箱条件下白眉姬鹟育雏期双亲投入[J].东北林业大学学报,2012,40(6):141-143.
(责任编辑:方 林)
Effects of body condition on the reproductive investment of Yellow-rumped Flycatcher
LIU Zhen-ni1,YAO Ji-yuan1,ZHANG Li-shi1,ZHAO Dian-zheng1,JIANG Yun-lei1,Wang Hai-tao2
(1.Animal’s Scientific and Technological Institute,Jilin Agricultural University,Changchun 130118,China; 2.School of Life Sciences,Northeast Normal University,Changchun 130024,China)
This paper conducted a study on the effects of body condition on reproductive investment in the Yellow-rumped Flycatchers(Ficedulazanthopygia) inside artificial nest box in secondary-forest of Zuojia Nature Protection area in Jilin Province during March to July in 2014—2015.The results indicated that female body condition had no significant correlation with the laying time,clutch size and nest materials weight.Male body condition was negative correlation with the laying time(r=-0.525,P<0.001),and positive correlation with clutch size(r=0.312,P=0.029).The remarkable correlations between female body condition and incubating duration were found(r=0.367,P=0.013).Found that adult body condition had no significant correlation with brooding times.In addition,brood size was positive correlation with brooding times(r=0.552,P<0.001).This study shows that Yellow-rumped Flycatchers can vary their reproductive investment according to their own body condition or their mate’s body condition.
Yellow-rumped Flycatcher;body condition;reproductive investment
1000-1832(2017)01-0105-05
10.16163/j.cnki.22-1123/n.2017.01.020
2016-04-01
国家自然科学基金资助项目(31270471,31470458).
刘珍妮(1990-),女,硕士研究生; 通讯作者:姜云垒(1962-),男,博士,教授,主要从事鸟类生态学研究.
Q 959.7+39 [学科代码] 180·5744
A
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:52:08
今日农业(2021年1期)2021-11-26 07:00:56
兽医导刊(2020年22期)2020-12-17 09:53:08
生物进化(2020年3期)2020-12-05 07:14:20
广东第二课堂·小学(2019年5期)2019-06-14 08:26:35
汽车观察(2018年12期)2018-12-26 01:05:26
森林与人类(2018年6期)2018-12-03 10:06:24
科学大观园(2018年1期)2018-05-30 10:48:04
生殖医学杂志(2015年5期)2015-04-07 04:49:20
中国果业信息(2014年1期)2014-01-23 11:15:01
