
不同唤醒度负性刺激抑制控制特点
2017-03-11 05:44赵绍晨郑希付
心理研究 2017年1期
赵绍晨郑希付
(1公安边防部队高等专科学校,广州 510663;2华南师范大学心理学院,广州 510631)
不同唤醒度负性刺激抑制控制特点
赵绍晨1郑希付2
(1公安边防部队高等专科学校,广州 510663;2华南师范大学心理学院,广州 510631)
本研究运用事件相关电位(ERP)技术,采用双选择情绪oddball范式,探讨不同唤醒度负性刺激的行为抑制特点。实验选取15名大学生作为被试,要求被试对标准刺激和偏差刺激(75%vs.25%)分别作不同的按键反应,记录按键前后的脑电。偏差刺激由高、低唤醒两种来自国际情绪图片系统的情绪图片组成。本研究主要分析反应抑制过程的N2和P3的波幅和潜伏期。结果显示:对高、低唤醒刺激的抑制反应诱发的N2波幅和潜伏期没有差异,对高唤醒刺激抑制反应诱发的P3波幅大于低唤醒的P3波幅,并且差异显著。结论:对高唤醒刺激进行抑制控制的结果是产生更大的P3波幅,这一过程的结果显示,人类对高唤醒刺激更难进行抑制控制。
唤醒度;行为抑制;oddball;ERP
1 引言
行为抑制作为执行功能的一个重要方面,反映了个体对不恰当刺激行为反应的合理控制,在人类的生存适应中起着重要的作用。Barkley认为,行为抑制包括对优势反应的抑制、停止正在进行的无效的或是与目标无关的行为、对内源性或外源性干扰的控制[1]。而行为抑制能力的缺失或异常常导致冲突或犯罪行为的发生[2]。近年来,关于行为抑制控制的研究发现,执行行为抑制任务的神经基础是额叶-基底神经节回路[3],但是对额叶各区在抑制过程中的作用看法不一[4]。其中一种观点认为,右侧腹外侧前额皮层(ventrolateral prefrontal cortex,VLPFC)负责抑制无关行为,而背侧前额叶(dorsolateral prefrontal cortex,DLPFC)参与工作记忆,前扣带回(anterior cingulate cortex,ACC)探测与目标不一致的冲突[5,6]。其中,右侧VLPFC广泛参与反应抑制、记忆提取抑制、任务定势切换等抑制控制任务,因而被认为是具有一般抑制功能的脑区[7]。最近,Mostofsky和Simmonds[4,7]提出前部辅助运动区(pre-supplementary motor area,pre-SMA)是选择适当的行为的关键区域,其中包括选择投入到适当的运动反应和选择抑制不适当的运动反应,而VLPFC参与维持工作记忆中的信息,DLPFC则参与操作工作记忆中的信息。另外,Hirokazu和Bekkker等研究者发现眶额叶皮层与前扣带回在行为抑制活动的完成中具有重要作用[8]。也有相关的研究发现,在行为抑制过程中,前扣带回的主要作用是冲突监控和错误监控[9,10]。采用go/nogo及双选择oddball范式的研究显示,额-中央区N2、中央-顶区P3成分是与行为抑制控制过程密切相关的两个ERP成分[11]。行为控制相关N2成分被认为主要与冲突觉察与监控有关;而行为控制相关P3的出现则是行为抑制加工过程的直接指标,比如,Go/Nogo任务中P3峰值的出现往往代表着行为抑制加工的完成[8,11]。
人类的情绪活动对行为抑制有着重要的影响[12,13],如负性情绪会导致攻击行为的发生[14]。有研究显示,冲动性人格特质与个体的情绪稳定性存在着显著的负相关[15]。有行为失控症状的精神病患者,如边缘型人格障碍患者、反社会人格障碍患者,在负性情绪下其行为控制缺陷症状更为明显[16],说明个体的情绪状态可能对行为控制过程产生影响。事实上,情绪与行为控制的交互作用近期得到了脑成像研究的支持[17-20]。
情绪是人格系统的动力核心,不同的情绪状态会对行为、记忆、注意等活动有着不同程度的调节作用,负性情绪可引发对环境中负性刺激的注意偏向。Goldstein等提出负性情绪唤起个体的运动行为(为了生存而战斗或逃跑),因此抑制行为反应变得困难。而在相关研究中发现,脑岛前叶在行为抑制过程中受到情绪线索的选择性激活[21,22]。随着抑制难度的增大,负性情绪线索对左侧脑岛的激活也逐渐增大[23]。这些结果表明,脑岛在情绪抑制任务中的激活反映了负性情绪行为抑制线索编码需要更多的认知资源[14]。那么,同样是负性情绪刺激,如果唤醒度不同,行为抑制会表现出什么样的特点呢?
近年来,出现了大量关于情绪与行为抑制的研究[4,18,23,22],但研究一方面主要集中于外显、内隐和情绪干扰任务的效价研究[21,24,25,26-31],另一方面主要集中于生理机制的研究[3,4,32,33-36]。 但是研究很少涉及情绪唤醒度的效应,尽管Goldstein与Shafritz等人的fMRI研究揭示了情绪条件下反应抑制的脑机制,却难以回答神经结构的激活是来源于情绪加工本身,还是来源于情绪对反应抑制的影响[14,21]。相关研究发现,一定程度的消极情绪对任务加工有促进作用。而对于情绪刺激的研究大都集中于效价的维度上,关于消极情绪刺激对任务加工的影响尚未有进一步的研究,根据情绪的维度分类,对于同一种效价,唤醒度维度可能也会产生一定程度的影响。本研究采用高时间分辨率ERP技术,及情绪双选择oddball范式考察不同唤醒度负性情绪的行为抑制特点。
2 实验方法
2.1 被试
15名在校大学生作为被试,其中7名男性,8名女性,年龄范围在18~23岁,平均年龄约20岁,所有被试身心健康,均为右利手,视力正常或矫正后正常,均签署了实验知情同意书,实验结束后给予少许报酬。
2.2 刺激材料
采用双选择oddball范式。在实验中,刺激材料均分为标准刺激和偏差刺激 (其比例为75%vs 25%)两类。标准刺激为一个杯子的自然景物图片,偏差刺激为高唤醒度负性刺激,低唤醒度负性刺激各51张。所有刺激图片均来自国际情绪图片系统。负性图片分为高唤醒度组和低唤醒度组。高低唤醒度图片的效价保持一致,对高低两种唤醒度图片的效价进行t检验,t=1.07,差异不显著;对两组图片的唤醒度进行比较,t=-3.63,p<0.00,差异十分显著。其中高唤醒度组的效价M=3.43,SD=0.32;唤醒度M= 5.00,SD=0.76;低唤醒度组的效价M=3.54,SD=0.70;唤醒度M=4.36;SD=1.00。图片分辨率均为100像素/英寸,大小统一为341像素×256像素。被试双眼与屏幕距离约100cm。
2.3 ERP记录
实验仪器为Brain Product脑电记录系统,参考电极置于双侧乳突连线,前额接地,采用32导电极帽记录脑电,同时记录水平眼电和垂直眼电,滤波带通为DC-100 Hz,采样频率为500Hz/导,头皮电阻降至5千欧以下。进行离线分析(off-line analysis),分析时程(epoch)为700ms,含基线100ms,自动矫正眨眼等伪迹,波幅大于±80μV者在叠加中被自动剔除。
2.4 程序和任务
实验共有3个block,每个block包括136个刺激试次(trial)。各唤醒度条件刺激图片呈现顺序完全随机。在实验开始之前,告知被试本实验考察个体的反应选择及对优势反应进行抑制的能力。首先,在电脑屏幕中央出现十字注视点,随机持续时间为600~800ms。然后呈现1000ms的随机刺激。被试的任务是当标准刺激出现时用左(右)手食指按F键反应;而当偏差刺激出现时,用右(左)手食指按J键反应。反应手进行block和被试间平衡。刺激呈现时间上限1000ms,刺激呈现随被试按键终止。因此,当刺激呈现之后,被试需要在1000ms之内尽可能快速而准确的作按键反应,在练习阶段提供被试对两类刺激图片的反应正确反馈。然后是不定间隔的注视点,以此循环。正式实验之前被试先进行20个试次的练习,所有被试都必须在练习阶段反应正确率达到100%再进入正式实验。记录标准刺激及各唤醒度偏差刺激诱发的EEG活动。实验结束后,对各个被试进行有关实验情况的访谈。
2.5 ERP数据分析和统计
分析时程为700ms,其中刺激呈现前100ms作为基线。对各唤醒度的负性偏差刺激及标准刺激诱发的ERP分别进行叠加和平均。由图1可知,情绪唤醒度效应主要出现在头皮中央及额-中央记录区域。因此,本研究选择以下的14个电极位置记录的ERP波形用于统计分析:FZ,FCZ,CZ,PZ,FP1,FP2,F7,F3,F4,F8,FC5,CP5,CP6,FC6。偏差刺激与标准刺激诱发的ERP从大约100ms开始分离,该差异一直持续到大约500ms结束。这些差异在差异波上集中表现为与晚期抑制控制相关N2(210~300ms)及P3(380~430ms)成分。因此,本研究首先通过平均波幅分析在 210~300ms及 380~430ms两窗口检验行为控制ERP效应的显著性。分析因素为刺激类别(偏差刺激,标准刺激)与电极点(14个记录点)。在行为控制效应显著的基础上,通过测量各唤醒度条件下N2及P3的潜伏期(刺激呈现到波峰之间的时间距)和波幅(基线到波峰),进一步分析负性情绪不同唤醒度对行为控制过程的影响。对上述各成分的波幅与潜伏期进行两因素重复测量方差分析 (ANOVA)。重复测量因素为唤醒度(高,中,低)及电极(14个电极点)。采用Greenhouse-Geisser法矫正自由度以及P值。
3 实验结果
3.1 行为结果
被试对标准刺激及偏差刺激的反应正确率均超过95.84%,对于正确率,刺激唤醒度及刺激类型主效应均不显著。对反应时间,被试对偏差刺激的反应时间(M=426.49±52.35)显著大于对标准刺激的反应时间(M=499.70±46.57),F(1,28)=16.38,p<0.000。此外,对于反应时间,唤醒度主效应显著,F(2,42)=11.74,p<0.001。高唤醒条件下的反应时显著大于中性条件下和低唤醒条件下的反应时间。被试在标准刺激、高唤醒的偏差刺激和低唤醒的偏差刺激条件 下的反应时间分别为:422.98ms,496.48ms,493.67ms。由此可知,被试在各偏差刺激条件下均出现了显著的反应延迟效应,该效应在高唤醒刺激条件下尤为突出。
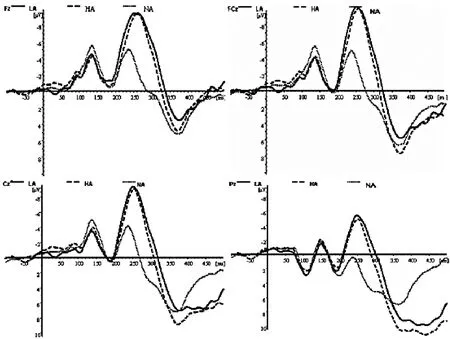
图1 LA(低唤醒度),HA(高唤醒度),NA(中性图片)
3.2 ERP成分与统计分析
对表现行为控制效应的原始波形而言,对N2在210~300ms时间窗的平均波幅重复测量方差分析显示,刺激类别的主效应显著[F(2,28)=11.20;p<0.01],电极点主效应显著[F(13,182)=7.71;p<0.01]。偏差刺激诱发出比标准刺激更大的负偏向,该偏向在差异波中集中体现为显著的额中央区N2活动。高、低唤醒刺激诱发的N2波幅差异不显著。平均波幅在中前部记录点显著大于后部记录点,说明出现了抑制效应。此外,刺激类别与电极点交互效应显著[F(26,364)=4.06;p<0.01]。对不同条件的潜伏期进行重复测量方差分析显示出显著的刺激类别差异 [F(2,28)=16.60;p<0.000]。对P3在380~430ms时间窗的平均波幅统计分析表现出显著的刺激类别 [F(2,28)=10.46;p<0.01]和电极点 [F(13,182)=2.22;p<0.05]主效应。其中高唤醒刺激的P3波幅大于低唤醒刺激的P3波幅。对不同条件的潜伏期进行重复测量方差分析,刺激类别差异不显著。总的来讲,中央-顶区的波幅显著大于前部记录区域的波幅。偏差刺激,尤其在中央区及后顶区,诱发出相比标准刺激更大的显著正偏向,该偏向在差异波中表现为明显的P3活动。由此可知,本实验条件下,偏差刺激诱发出明显的行为抑制效应,该效应集中体现为额-中央区域的N2活动差异及中央顶区P3活动差异(见图1)。
4 讨论
本研究采用双选择的oddball范式,结果发现,个体对偏差刺激的反应时间显著长于对标准刺激的反应时间,说明被试对偏差刺激进行了行为抑制。同时也发现不同唤醒度的视觉情绪刺激对个体的行为控制过程有显著影响。该影响在行为层面上表现为,随着负性情绪唤醒度的增强,个体对偏差刺激反应延迟效应明显,说明个体对偏差刺激的加工除了知觉注意、刺激辨别、反应决策与执行等过程,也包括对优势反应的行为抑制。双选择oddball范式和go/no go范式都是oddball范式的变式,双选择oddball范式中对偏差刺激的反应相当于go/no go范式的no go任务,均是反应对优势行为的抑制控制过程[37,38]。这一范式引发出两个成分 N2 (200-400ms)和P3(300-500ms),一般在前额脑区波幅最明显,并且no go引发的成分均小于go任务引发的成分[39]。No go的N2和P3成分反映了反应抑制的不同过程,N2是对认知过程的一个广泛体现,并不是严格反映抑制控制过程本身。而P3成分则主要是对抑制控制过程的结果进行评估[38]。
以往研究发现,行为抑制任务中N2与反应冲突检测有关,由反应冲突任务诱发的N2显著大于无反应冲突的任务。本研究在210~300ms窗口观测到显著的中央区N2活动,统计结果显示N2波幅唤醒度主效应显著,高唤醒度诱发的波幅较大。由于N2波幅代表着冲突监控中的注意分配[37],这表明高唤醒的负性刺激最大程度的引起了对刺激监控的注意分配。另一方面,偏差刺激中高唤醒度和低唤醒度的负性刺激诱发的脑电比标准的中性刺激显著延迟,由于潜伏期代表着反应冲突觉察的快慢[37],因此,个体对高唤醒度的刺激有更强的抑制。此外,N2还与反应抑制有关[11,38],代表抑制过程的早期阶段,较大的N2即代表在nogo测试条件下中前扣带回的活跃程度较大。时间压力增大意味着反应趋势的增大,N2-nogo也随之增大。该结果提示高唤醒度负性刺激的条件下反应冲突监控需要消耗个体较多的注意资源,觉察反应冲突的速度比中性刺激条件更慢。负性情绪事件具有重要的生存适应价值,人脑对该类事件存在加工偏向。近期国内的一些研究也表明,随着效价强度的增强,负性情绪对任务相关维度加工的影响也增大。本研究结果显示,高、低唤醒度条件诱发的N2的波幅没有差异。这表明,对于不同唤醒的负性刺激,早期的冲突监控过程没有差异,根据本研究的结果,我们可以认为,人类对于高和低唤醒的负性刺激的抑制控制过程早期投入的注意资源是没有差异的。
对行为抑制控制而言,行为抑制条件下中央顶区P3活动的出现是抑制控制加工的直接指标,行为抑制条件诱发的波幅显著大于无控制条件。而且P3被认为是抑制过程的晚期阶段的重要指标。事实上,本研究中很多峰值在刺激呈现后200~300ms之间的ERP成分都有表现。我们的研究结果是,在前额叶的FZ点,情绪刺激的P3波幅比中性刺激的波幅小,偏差刺激相比标准刺激引发较小的P3波幅,也就是说情绪刺激比中性刺激耗费了较少的脑资源。进一步对高、低唤醒刺激行为抑制诱发的P3波幅分析表明,高唤醒条件下的P3波幅相比低唤醒条件下的大,说明在行为控制的晚期,被试在对高唤醒的刺激进行行为抑制时,需要付出更多的资源,高唤醒度的负性情绪刺激更难进行行为抑制。如前所述,人脑对负性事件具有优先加工性。以往的研究显示,情绪刺激影响注意,并易化了接下来的加工。因此高唤醒刺激更多的激活相关脑区,可能影响接下来的行为抑制过程,这些都会导致对高唤醒负性刺激比低唤醒刺激更难抑制控制。
5 结论
本研究采用双选择oddball范式,发现高唤醒度视觉情绪对行为抑制控制的效果具有显著影响。该影响主要表现为对高唤醒度的负性情绪存在更难抑制控制的执行效果。对于行为抑制而言,高唤醒度负性刺激出现后,在抑制控制的早期和低唤醒刺激没有差异,这个时候两种唤醒的N2波幅没有差异。而在行为抑制的晚期,对高唤醒度的负性刺激抑制控制引发的P3波幅明显大于对低唤醒负性刺激引发的P3波幅,这说明,对高唤醒负性刺激的抑制控制,被试要付出更大的努力,因此更难抑制控制。本研究结果表明,对高唤醒负性刺激的行为抑制更难。参考文献
1 Aron A R,Robbins T W,Poldrack R A.Inhibition and the right inferior frontal cortex.Trends in Cognitive Sciences,2004,8(4):170-177.
2 Törnquist K,Malm A,Pasternack M,et al.Tumor necrosis factor-α,sphingomyelinase,and ceramide inhibit store-operated calcium entry in thyroid FRTL-5 cells.JournalofBiologicalChemistry, 1999, 274(14):9370-9377.
3 Dillon D G, PizzagalliD A.Inhibition ofaction,thought,and emotion:A selective neurobiological review.Applied and Preventive Psychology,2007,12(3):99-114.
4 Mostofsky S H,Simmonds D J.Response inhibitionand response selection:Two sides of the same coin.JournalofCognitive Neuroscience, 2008, 20(5):751-761.
5 Buchsbaum B R,Greer S,Chang W L,et al.Metaanalysis ofneuroimaging studies ofthe Wisconsin Card-Sorting task and component processes.Human Brain Mapping,2005,25(1):35-45.
6 Petrides M,Alivisators B,Freys.Differential activation of the human orbital,mid-ventrolateral,and mid-dorsolateral prefrontal cortex during the processing of visual stimuli.Proceedings of the National Academy of Sciences,2002,99(8):5649-5654.
7 Aron A R,Robbins T W,Poldrack R A.Inhibition and the right inferior frontal cortex.Trends in Cognitive Sciences,2004,8(4):170-177.
8 Bekker E M,Kenemans J L,Verbaten M N.Source analysis of the N2 in a cued Go/NoGo task.Cognitive Brain Research,2005,22(2):221-231.
9 Garavan H,Ross T J,Murphy K,et al.Dissociable executive functions in the dynamic control of behavior:Inhibition,error detection,and correction.Neuroimage,2002,17(4):1820-1829.
10 Rubia K,Russell T,Bullmore E T,et al.An fMRI study of reduced left prefrontal activation in schizophrenia during normal inhibitory function.Schizophrenia Research,2001,52(1):47-55.
11 Falkenstein M,Hoormann J,Hohnsbein J.ERP components in Go/Nogo tasks and their relation to inhibition.Acta Psychologica,1999,101(2):267-291.
12 Rowe G,Hirsh J B,Anderson A K.Positive affect increases the breadth of attentional selection.Proceedings of the National Academy of Sciences,2007,104(1):383-388.
13 Tsetsenis T,Ma X,Iacono L L,et al.Suppression of conditioning to ambiguous cues by pharmacogenetic inhibition ofthe dentategyrus.NatureNeuroscience,2007,10(7):896-902.
14 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
15 Scheier M F,Carver C S,Bridges M W.Distinguishing optimism from neuroticism(and trait anxiety,selfmastery,and self-esteem):A reevaluation of the Life Orientation Test.Journal of Personality and Social Psychology,1994,67(6):1063.
16 Posner M I,Rothbart M K,Vizueta N,et al.Attentional mechanisms of borderline personalitydisorder. Proceedingsofthe NationalAcademy ofSciences,2002,99(25):16366-16370.
17 Silbersweig D,Clarkin J F,Goldstein M,et al.Failure of frontolimbic inhibitory function in the context of negative emotion in borderline personality disorder.American JournalofPsychiatry, 2007, 164(12):1832-1841.
18 Shachaf C M,Perez O D,Youssef S,et al.Inhibition of HMGcoA reductase by atorvastatin prevents and reverses MYC-induced lymphomagenesis.Blood, 2007,110(7):2674-2684.
19 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
20 Stadler A,Schima W,Ba-Ssalamah A,et al.Artifacts in body MR imaging:Their appearance and how to eliminate them.European Radiology,2007,17(5):1242-1255.
21 Goldstein M,Brendel G,Tuescher O,et al.Neural substrates of the interaction of emotional stimulus processing and motorinhibitory control: An emotional linguistic go/no-go fMRI study.Neuroimage,2007,36(3):1026-1040.
22 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
23 Schulz E,Maurer U,van der Mark S,et al.Impaired semantic processing during sentence reading in children with dyslexia:Combined fMRI and ERP evidence.Neuroimage,2008,41(1):153-168.
24 Nunez J M,Casey B J,Egner T,et al.Intentional false responding shares neural substrates with response conflict and cognitive control.Neuroimage,2005,25(1):267-277.
25 Chiu P H,Holmes A J,Pizzagalli D A.Dissociable recruitmentofrostralanteriorcingulate and inferior frontal cortex in emotional response inhibition.Neuroimage,2008,42(2):988-997.
26 Hare T A,Tottenham N,Davidson M C,et al.Contributions of amygdala and striatal activity in emotion regulation.Biological Psychiatry,2005,57(6):624-632.
27 Hare T A,Tottenham N,Galvan A,et al.Biological substratesofemotionalreactivity and regulation in adolescence during an emotional go-nogo task.Biological Psychiatry,2008,63(10):927-934.
28 Hare T A,Tottenham N,Galvan A,et al.Biological substratesofemotionalreactivity and regulation in adolescence during an emotional go-nogo task.Biological Psychiatry,2008,63(10):927-934.
29 Lynch J C,Ferreira L A S,Welling L,et al.Multiple intracranial meningiomas:Diagnosis,biological behavior and treatment.Arquivos de Neuro-psiquiatria,2008,66(3B):702-707.
30 Müller J L,Sommer M,Dhnel K,et al.Disturbed prefrontal and temporal brain function during emotion and cognition interaction in criminal psychopathy.Behavioral Sciences&the Law,2008,26(1):131-150.
31 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
32 Aron A R.The neural basis of inhibition in cognitive control.The Neuroscientist,2007,13(3):214-228.
33 Dolan R J.The human amygdala and orbital prefrontal cortex in behavioural regulation.Philosophical Transactions of the Royal Society of London B:Biological Sciences,2007,362(1481):787-799.
34 Elliott R,Rubinsztein J S,Sahakian B J,et al.Selective attention to emotional stimuli in a verbal go/no-go task:An fMRI study.Neuroreport,2000,11(8):1739-1744.
35 Garavan H,Ross T J,Murphy K,et al.Dissociable executive functions in the dynamic control of behavior:Inhibition,error detection,and correction.Neuroimage,2002,17(4):1820-1829.
36 Rubia K,Russell T,Bullmore E T,et al.An fMRI study of reduced left prefrontal activation in schizophrenia during normal inhibitory function.Schizophrenia Research,2001,52(1):47-55.
37 Nagy E,Potts G F,Loveland K A.Sex-related ERP differences in deviance detection.International Journal of Psychophysiology,2003,48(3):285-292.
38 Roche R A,Garavan H,Foxe J J,et al.Individual differences discriminate event-related potentials but not performance during response inhibition.Experimental Brain Research,2005,160(1):60-70.
39 Bokura H,Yamaguchi S,Kobayashi S.Electrophysiological correlates for response inhibition in a Go/NoGo task.Clinical Neurophysiology, 2001, 112(12):2224-2232.
The Behavior Inhibition Characteristics of Different Arousal Negative Stimulation
Zhao Shaochen1,Zheng Xifu2
(1 Public Security Frontier Defense Forces College,Guangzhou 510663;2 College of Psychology,South China Normal University,Guangzhou 510631)
This research discussed the behavior inhibit characteristics of different arousal negative stimulations with the event related potential technology and classical oddball paradigm.This study selected 15 undergraduate and graduate students as the subjects.They were asked to press the key as fast and accurately as possible according to different stimulations(high sensei stimulation,low sensei stimulation and standard stimulus)and corresponding brain wave of these different types of incentives were recorded.The results showed that there was a significant difference among different arousal negative stimulations.In 210~300ms time interval,average amplitude was prominently different as well as latency;In 380~430ms time interval,there was a significant difference in average amplitude,too;It was found from the analysis of behavior inhibition process that the average amplitude of N2 and P3 wave exist significant differences in average volatility.These results reflect that there is a notable difference in behavior inhibition of different arousal negative stimulation.In the condition of high arousal stimulation,it is individual reaction monitoring conflicts and subsequent brain activity of behavior inhibit that were enhanced.The high arousal negative emotion has an obvious facilitated effect to behavior inhibition.
arousal degree;behavior inhibition;oddball;ERP
郑希付,男,教授,博士生导师。Email:zhengxf@scnu.edu.cn
猜你喜欢
哈尔滨工程大学学报(2022年8期)2022-09-11
甘肃教育(2021年10期)2021-11-02
福建江夏学院学报(2021年6期)2021-08-10
心理学报(2021年1期)2021-01-29
初中生学习指导·中考版(2020年2期)2020-09-10
大连民族大学学报(2020年2期)2020-06-16
价值工程(2018年25期)2018-09-26
英美文学研究论丛(2018年1期)2018-08-16
中国医药指南(2017年3期)2017-11-13
中国现代医生(2017年24期)2017-10-17
