
Edaravone protects against oxygen-glucose-serum deprivation/restoration-induced apoptosis in spinal cord astrocytes by inhibiting integrated stress response
2017-03-09 06:05:50BinDaiTingYanYixingShenYoujiaXuHaibinShenDongChenJinrongWangShuanghuaHeQirongDongAiliangZhang
中国神经再生研究(英文版) 2017年2期
Bin Dai, Ting Yan, Yi-xing Shen You-jia Xu Hai-bin Shen, Dong Chen, Jin-rong Wang, Shuang-hua He, Qi-rong Dong, Ai-liang Zhang,
1 Department of Orthopedics, The Second Af filiated Hospital of Soochow University, Suzhou, Jiangsu Province, China
2 Department of Orthopedics, Binhai County People’s Hospital, Binhai, Jiangsu Province, China
3 Safety Assessment and Research Center for Drug, Pesticide and Veterinary Drug of Jiangsu Province, School of Public Health, Nanjing Medical University, Nanjing, Jiangsu Province, China
4 Department of Orthopedics, Third Af filiated Hospital of Soochow University, Changzhou, Jiangsu Province, China
Edaravone protects against oxygen-glucose-serum deprivation/restoration-induced apoptosis in spinal cord astrocytes by inhibiting integrated stress response
Bin Dai1,2, Ting Yan3, Yi-xing Shen1, You-jia Xu1, Hai-bin Shen2, Dong Chen2, Jin-rong Wang2, Shuang-hua He4, Qi-rong Dong1,*, Ai-liang Zhang4,*
1 Department of Orthopedics, The Second Af filiated Hospital of Soochow University, Suzhou, Jiangsu Province, China
2 Department of Orthopedics, Binhai County People’s Hospital, Binhai, Jiangsu Province, China
3 Safety Assessment and Research Center for Drug, Pesticide and Veterinary Drug of Jiangsu Province, School of Public Health, Nanjing Medical University, Nanjing, Jiangsu Province, China
4 Department of Orthopedics, Third Af filiated Hospital of Soochow University, Changzhou, Jiangsu Province, China
We previously found that oxygen-glucose-serum deprivation/restoration (OGSD/R) induces apoptosis of spinal cord astrocytes, possibly via caspase-12 and the integrated stress response, which involves protein kinase R-like endoplasmic reticulum kinase (PERK), eukaryotic initiation factor 2-alpha (eIF2α) and activating transcription factor 4 (ATF4). We hypothesized that edaravone, a low molecular weight, lipophilic free radical scavenger, would reduce OGSD/R-induced apoptosis of spinal cord astrocytes. To test this, we established primary cultures of rat astrocytes, and exposed them to 8 hours/6 hours of OGSD/R with or without edaravone (0.1, 1, 10, 100 μM) treatment. We found that 100 μM of edaravone signif i cantly suppressed astrocyte apoptosis and inhibited the release of reactive oxygen species. It also inhibited the activation of caspase-12 and caspase-3, and reduced the expression of homologous CCAAT/enhancer binding protein, phosphorylated (p)-PERK, p-eIF2α, and ATF4. These results point to a new use of an established drug in the prevention of OGSD/R-mediated spinal cord astrocyte apoptosis via the integrated stress response.
nerve regeneration; edaravone; apoptosis; astrocytes; integrated stress response; reactive oxygen species; PERK; eIF2α; activating transcription factor 4; CCAAT/enhancer binding protein homologous protein; caspase-3, caspase-12; neural regeneration
Graphical Abstract
Introduction
Astrocytes are ubiquitously distributed in the central nervous system and play an important role in synapse formation, plasticity, and the development and maintenance of the blood—brain barrier (Chen et al., 2016). They also provide metabolic and trophic support to neurons (Pabst et al., 2016). Properly functioning astrocytes are particularly important in maintaining neuronal viability under ischemic conditions, where energy depletion and metabolic disruption are severe (Ouyang et al., 2013; Shindo et al., 2016). Impairment or dysfunction of astrocytes can result in neuronal death. Edaravone (3-methyl-1-phenyl-2-pyrazolin-5-one) is a free radical scavenger used in the treatment of acute ischemic stroke (Turan et al., 2017). It suppresses the productionof reactive oxygen species (ROS), early accumulation of oxidative products, and subsequent inf l ammatory responses. A growing body of evidence indicates that edaravone protects all three major cell types of the neurovascular unit (neuron, astrocyte, and cerebral endothelium). For example, it protects against MPP+-induced cytotoxicity in rat primary cultured astrocytes by inhibiting the mitochondrial apoptotic pathway (Chen et al., 2008). Wang et al. (2011) reported that edaravone treatment reduced brain edema, blood-brain barrier permeability, and neuronal death, and improved neurological function, in a rat model of traumatic brain injury. Furthermore, our previous study suggested that caspase-12 activation and the integrated stress response, which involves protein kinase R-like endoplasmic reticulum kinase (PERK), eukaryotic initiation factor 2-alpha (eIF2α) and activating transcription factor 4 (ATF4), are involved in the apoptosis of spinal cord astrocytes induced by oxygen-glucose-serum deprivation/restoration (OGSD/R) (Zhang et al., 2010).
Despite such reports of the favorable ef f ects of edaravone on the central nervous system, its role in OGSD/R-mediated astrocytic apoptosis remains elusive. In the present study, we investigated whether edaravone protects against the integrated stress response and caspase-12 in OGSD/R-mediated astrocytic apoptosis.
Materials and Methods
Animals
Female and male Sprague-Dawley rats, aged 3 months and weighing 200—250 g (Shanghai Super B&K Laboratory Animal Corp., Ltd., Shanghai, China, SCXK (Hu) 2008-0016) were housed under a 12-hour light/dark cycle with free access to food and water. Spinal cords were obtained from the newborn rat pups (1—2 days old) for astrocyte culture. All animal experiments were approved by the Animal Experimental Committee of Soochow University, China.
Primary culture of spinal cord astrocytes
The spinal cords were dissected under sterile conditions, the meninges carefully removed, and the cord tissue dissociated in 0.25% trypsin for 6 minutes at 37°C. The cell suspension was centrifuged at 219 × g for 5 minutes. Spinal cord astrocytes were cultured in accordance with the improved method of Black et al. (1993). The cultures were maintained at 37°C in a humidif i ed atmosphere of 5% CO2and 95 % air. More than 95 % of the cells were immunopositive for the astrocytic marker glial fi brillary acidic protein (GFAP). The cells were used for subsequent experiments at approximately 12—15 days in culture. They were divided into three groups, incubated under the following conditions: (1) control group: oxygen-glucose-serum (OGS)-supplied medium (Hyclone, Logan, UT, USA); (2) OGSD/R group: OGS-deprived medium for 8 hours, followed by OGS-supplied medium for 6 hours; (3) four OGSD/R + edaravone groups: incubation in edaravone (Sigma, St. Louis, MO, USA) at concentrations of 0.1, 1, 10, or 100 μM, and subsequent incubation in OGS-deprived medium for 8 hours, then OGS-supplied medium for 6 hours.
Cell viability assay
Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) was used to measure cell viability, according to the manufacturer’s protocol. Cells were plated on 96-well plates (5,000 cells/well) and allowed to adhere for 24 hours at 37°C in a humidif i ed atmosphere of 5% CO2and 95% air. Subsequently, CCK-8 solution was added to each well and the cells were incubated for an additional 2 hours at 37°C for optical absorbance measurement at 450 nm (ELx800 Absorbance Microplate Reader; Bio-Tek Instruments, Inc., Winooski, VT, USA).
Apoptotic assay
Apoptotic astrocytes were counted by plating the cells onto glass slides and allowing them to adhere for 24 hours at 37°C in a humidif i ed atmosphere of 5% CO2and 95% air. The cells were then fi xed for 1 hour in 4% paraformaldehyde at room temperature, washed three times with phosphate buf f ered saline (PBS) and stained with 5 μg/mL Hoechst 33342 (Sigma) for 30 minutes at 37°C in a moist chamber. The morphological features of apoptosis were observed by fl uorescence microscopy (Olympus BX51, Tokyo, Japan). At least 400 cells were counted from 12 randomly selected fi elds per dish. Apoptotic cells were def i ned as those stained with Hoechst 33342, showing bright nuclei, and calculated as a percentage of the total number of cells.
ROS
ROS levels were determined using 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA) (Beyotime Institute of Biotechnology, Haimen, China). Astrocytes in sub-conf l uent culture (5 × 104cells/cm2) adhered on Petri dishes were incubated with or without edaravone for 30 minutes, followed by 8 hours/6 hours of OGSD/R, and then incubated with 10 μM H2DCF-DA dissolved in PBS (1 mL) at 37°C for 30 minutes. Images were captured using an Olympus BX51 microscope coupled with an Olympus DP70 digital camera. Fluorescence intensity was measured at excitation and emission wavelengths of 488 nm and 530 nm, respectively. All ROS levels were normalized to the mean ROS level of the control group.
Immunostaining
All incubations were at room temperature unless stated otherwise. Cells were fi xed in 4% paraformaldehyde in PBS for 30 minutes and permeabilized in 2% bovine serum albumin/0.2% Triton X-100/PBS for 1 hour. They were then incubated with goat anti-GFAP polyclonal antibody (1:50; Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at 4°C, followed by FITC-conjugated rabbit anti-goat IgG (1:100; Santa Cruz Biotechnology) for 2 hours in the dark. After washing with PBS, the cells were incubated with mouse monoclonal anti-caspase-12 antibody (1:100) for 10 hours at 4°C, then TRITC-conjugated goat anti-rabbit IgG (1:100; Santa Cruz Biotechnology) for 2 hours in the dark, and fi nally with 4′,6-diamidino-2-phenylindole (DAPI) (1:10,000) for 15 minutes in the dark. Images were captured using a fl uorescence microscopy imaging system (Olympus, Hatagaya, Japan).
Western blot assay
Cells were plated on 6-well plates and allowed to adhere for 24 hours at 37°C before treatment. Astrocyte cell lysates were prepared in ice-cold radioimmunoprecipitation assay buf f er (Beyotime) supplemented with protease inhibitor cocktail (Roche, Basel, Switzerland) and 1 mM phenylmethyl sulfonylf l uoride (Calbiochem). after centrifugation at 12,000 × g for 15 minutes, the lysate supernatants were denatured and loaded onto a 10 % gel for sodium dodecyl sulfate polyacrylamide gel electrophoresis. The separated proteins (1 μg) were transferred to nitrocellulose membranes. The membranes were blocked by overnight incubation in 5% nonfat milk at 4°C, and then incubated with the primary antibodies or 3 hours at room temperature. Individual blots were probed with mouse polyclonal anti-cleaved-caspase-3, mouse monoclonal anti-B-actin (Santa Cruz Biotechnology), rabbit polyclonal anti-PERK, rabbit polyclonal anti-phosphorylated (p)-PERK (CST Inc., Beverly, MA, USA), polyclonal anti-eIF2α, polyclonal anti-p-eIF2α (S51), polyclonal anti-caspase-12, polyclonal anti-ATF4, and polyclonal anti-CCAAT/enhancer binding protein homologous protein (CHOP) (Abcam, USA).The membranes were washed and incubated with horseradish peroxidase-conjugated goat anti-mouse or anti-rabbit IgG (1:5,000) for 1 hour at room temperature. The immune complexes were visualized using an enhanced chemiluminescence kit (Pierce, Rockford, IL, USA) according to the manufacturer’s protocol, and quantified by gray value measurement using a Tanon imaging system (Tanon, Shanghai, China). The relative expression levels were calculated from the gray value ratios of the target protein to B-actin (housekeeping protein).
Statistical analysis
Paired t-tests or one-way analysis of variance followed by the Student-Newman-Keuls post hoc test were carried out using SPSS 13.0 soThware (SPSS, Chicago, IL, USA). Statistical signif i cance was def i ned as P < 0.05.
Results
Edaravone inhibited OGSD/R-induced apoptosis of astrocytes in rat spinal cord
The effect of edaravone on the apoptotic status of cells in culture under OGSD/R was fi rst assessed by morphological examination. Apoptotic cells with condensed or fragmented nuclei were detected after OGSD/R (Figure 1A). Cultures exposed to 8 hours/6 hours of OGSD/R had significantly more apoptotic cells than non-treated cultures (P < 0.05; Figure 1B). Moreover, high concentrations of edaravone (10 and 100 μM) signif i cantly suppressed apoptosis after 8 hours/6 hours of OGSD/R (P < 0.05; Figure 1B). The CCK-8 cell viability assay conf i rmed the Hoechst staining results (Figure 1C).
Edaravone inhibitd the release of ROS after OGSD/R
ROS production in spinal cord astrocytes was measured by fl uorescent staining with fl uorescent dye H2DCF-DA (Figure 2). OGSD/R led to an increase in ROS release in spinal cord astrocytes. In addition, 100 μM edaravone significantly inhibited the release of ROS after 8 hours/6 hours of OGSD/R (P < 0.05; Figure 2).
Edaravone reduced caspase-12 protein expression after OGSD/R
Western blot assay revealed that cleaved caspase-3 expression was elevated after 8 hours/6 hours of OGSD/R (Figure 3). Edaravone-treated cells had significantly lower cleaved caspase-3 protein expression after OGSD/R than those exposed to OGSD/R without edaravone (Figure 3; P < 0.05). Cytoplasmic staining of pro-caspase-12 was also elevated after 8 hours/6 hours of OGSD/R, and this effect was also smaller in edaravone-treated cells (P < 0.05; Figure 4A). A similar increase was observed in cleaved-caspase-12 expression in OGSD/R-exposed astrocytes, and this was also reduced by edaravone (P < 0.05; Figure 4B, C).
Edaravone reduced the expression of CHOP after OGSD/RTo investigate the effect of edaravone on the induction of CHOP in astrocytes exposed to OGSD/R, we examined CHOP expression by western blot assay. CHOP expression was elevated after OGSD/R, and this ef f ect was reduced by edaravone (P < 0.05; Figure 5).
Edaravone reduced expression of p-PERK, p-eIF2α, and ATF4 after OGSD/R
Finally, we investigated the effect of edaravone on PERK/ eIF2α/ATF4 signaling after OGSD/R. Protein expression of p-PERK, p-eIF2α, and ATF4 was significantly lower after edaravone treatment in astrocytes exposed to 8 hours/6 hours of OGSD/R than in those without edaravone (P < 0.05; Figure 6). This indicates that edaravone protects spinal cord astrocytes against OGSD/R-induced apoptosis by inhibiting the integrated stress response and caspase-12.
Discussion
We previously reported that OGSD/R induces apoptosis of spinal cord astrocytes, and that this effect is regulated by the PERK/eIF2α/ATF4 integrated stress response and activation of caspase-12 (Zhang et al., 2010). Therefore, suppressing either PERK/eIF2α/ATF4 or caspase-12 activation may be novel targets for preventing OGSD/R-mediated astrocyte apoptosis. Here, we show that edaravone suppresses OGSD/R-induced elevation of PERK/eIF2α/ATF4 and CHOP expression, and activation of caspase-12 and caspase-3, subsequently decreasing apoptosis in spinal cord astrocytes.
Edaravone is an established drug in Japan, where it has been used to treat patients with acute ischemic stroke for more than 10 years, but it does not have marketing authorization in Europe or the USA (Edaravone Acute Infarction Study, 2003). Other free radical scavengers, such as ebselen and tirilazad, progressed to clinical trials in patients with cerebral infarction, but were terminated because of inadequate therapeutic ef f ects (Handa et al., 2000; Hukkelhoven et al., 2002). In contrast, edaravone has been administered within 24 hours of cerebral infarction in patients with lacunae,large-artery afterosclerosis, and cardioembolic cerebral infarction (Okamura et al., 2014). Here, we have shown for the first time that edaravone significantly suppresses apoptosis in spinal cord astrocytes after OGSD/R.
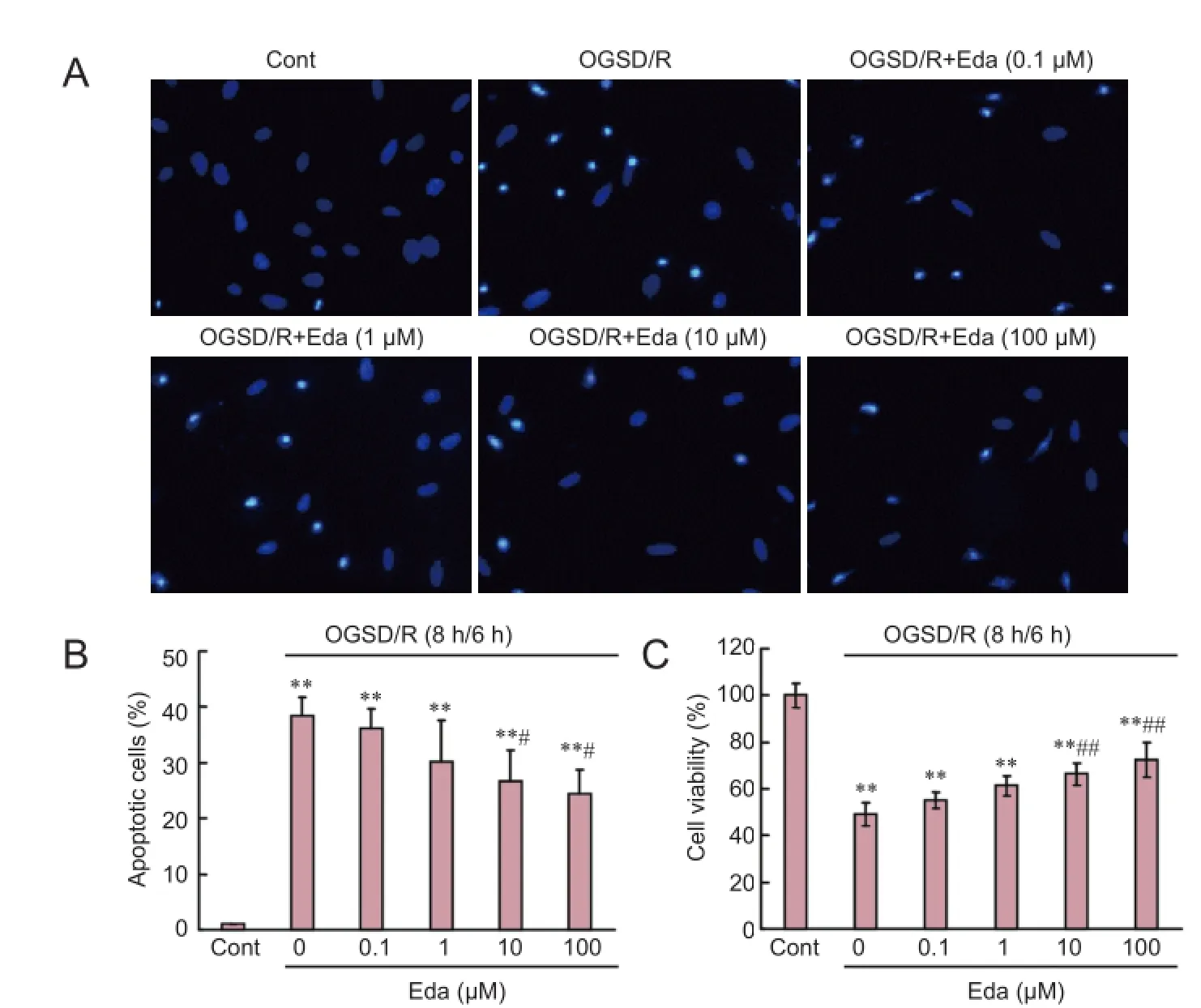
Figure 1 Edaravone (Eda) reduces oxygen-glucose-serum deprivation/ restoration (OGSD/R)-induced apoptosis of spinal cord astrocytes.
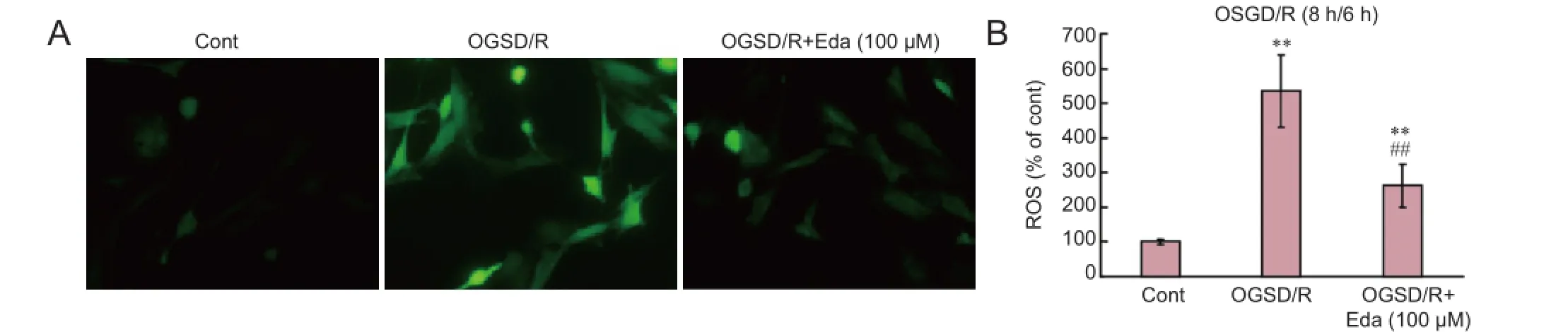
Figure 2 Edaravone (Eda) reduces oxygen-glucose-serum deprivation/restoration (OGSD/R)-induced release of reactive oxygen species (ROS) in spinal cord astrocytes.
ROS generation occurs during endoplasmic reticulum stress by protein oxidation (Bhandary et al., 2012). ROS levels increase upon exposure to toxic agents, such as irradiation and environmental pollutants, or during enzymatic reactions, including amino acid oxidase and NADP/NADPH oxidase (Kim et al., 2014). Edaravone acts as a strong scavenger of free radicals in multiple cell lines and experimental systems, including cancer cells, neurons and peripheral blood lymphocytes (Mizuno et al., 2007; Chen et al., 2014; Cheng and Zhang, 2014). Consistent with these reports, we found that edaravone inhibited the generation of ROS in spinal cord astrocytes after OGSD/R.
The endoplasmic reticulum is an active organelle that acts by folding and modifying secretory and membrane proteins. The unfolded protein response is a cellular stress response related to the endoplasmic reticulum and is activated upon the detection of an accumulation of unfolded or misfolded proteins in the lumen of the endoplasmic reticulum. A malfunction of the endoplasmic reticulum stress response caused by aging, genetic mutations, or environmental factors can result in various diseases, such as diabetes, inf l ammation, and nervous system disorders including Alzheimer’s disease, Parkinson’s disease, and cerebral ischemia (Yoshida, 2007; Lin, 2015; Liu and Connor, 2015; Yang and Hu, 2015). PERK undergoes dimerization and autophosphorylation to initiate a transient cellular translational arrest and inactivating eukaryotic translation initiation factor (eIF2α) (Harding et al., 1999; Ma et al., 2002). p-eIF2α can translate genes, one of which encodes ATF4, a member of the cAMP response element-binding family of transcription factors (Lu et al., 2004). In the present study, edaravone blocked OGSD/R-induced phosphorylation of PERK andeIF2α and expression of ATF4 in spinal cord astrocytes.
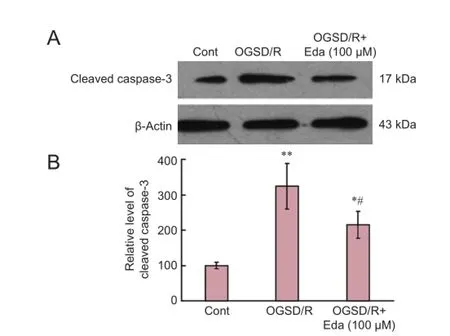
Figure 3 Edaravone (Eda) inhibits the caspase-3 signaling pathway after oxygen-glucose-serum deprivation/restoration (OGSD/R).
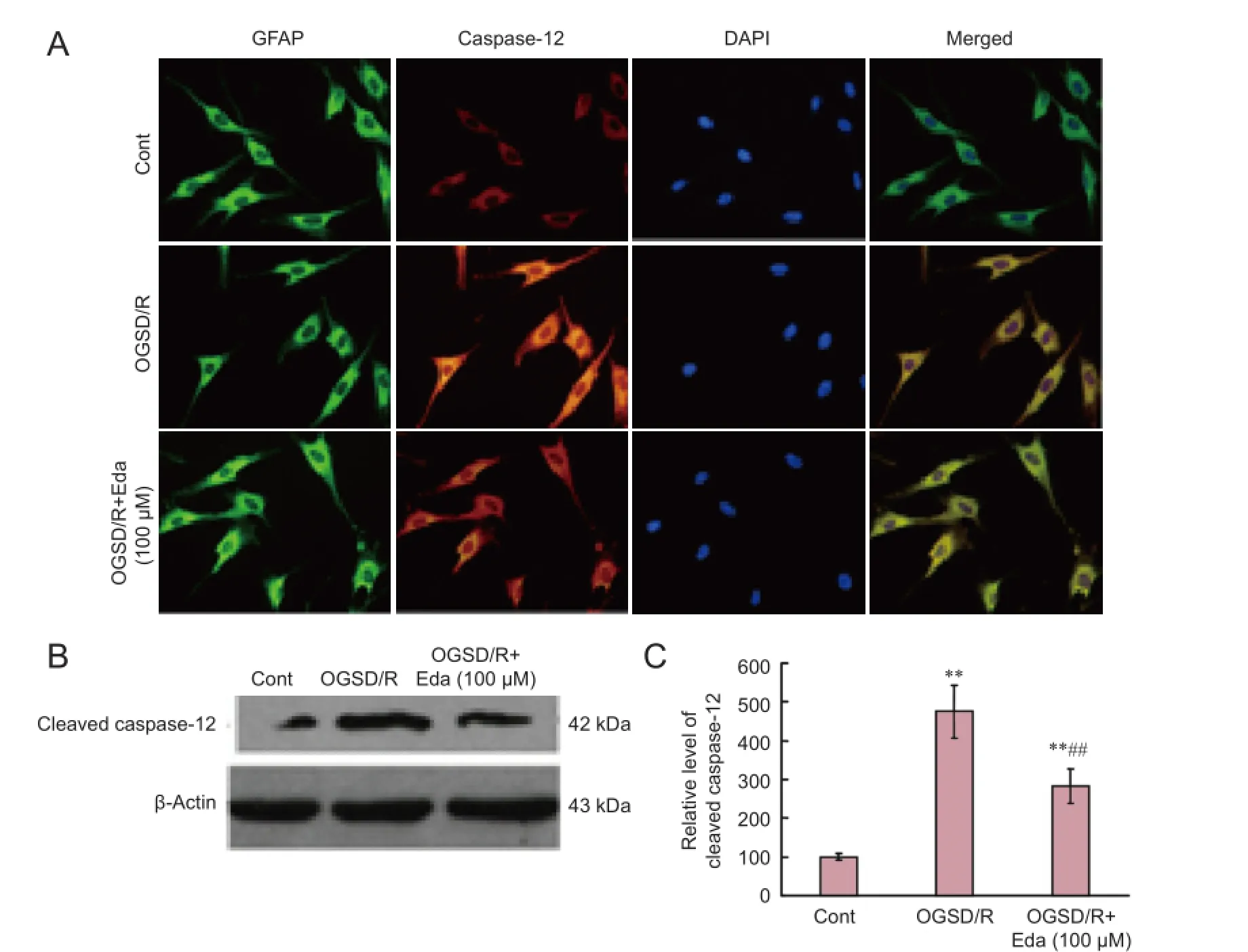
Figure 4 Edaravone (Eda) reduces oxygen-glucose-serum deprivation/restoration (OGSD/R)-induced increase in caspase-12 protein expression.
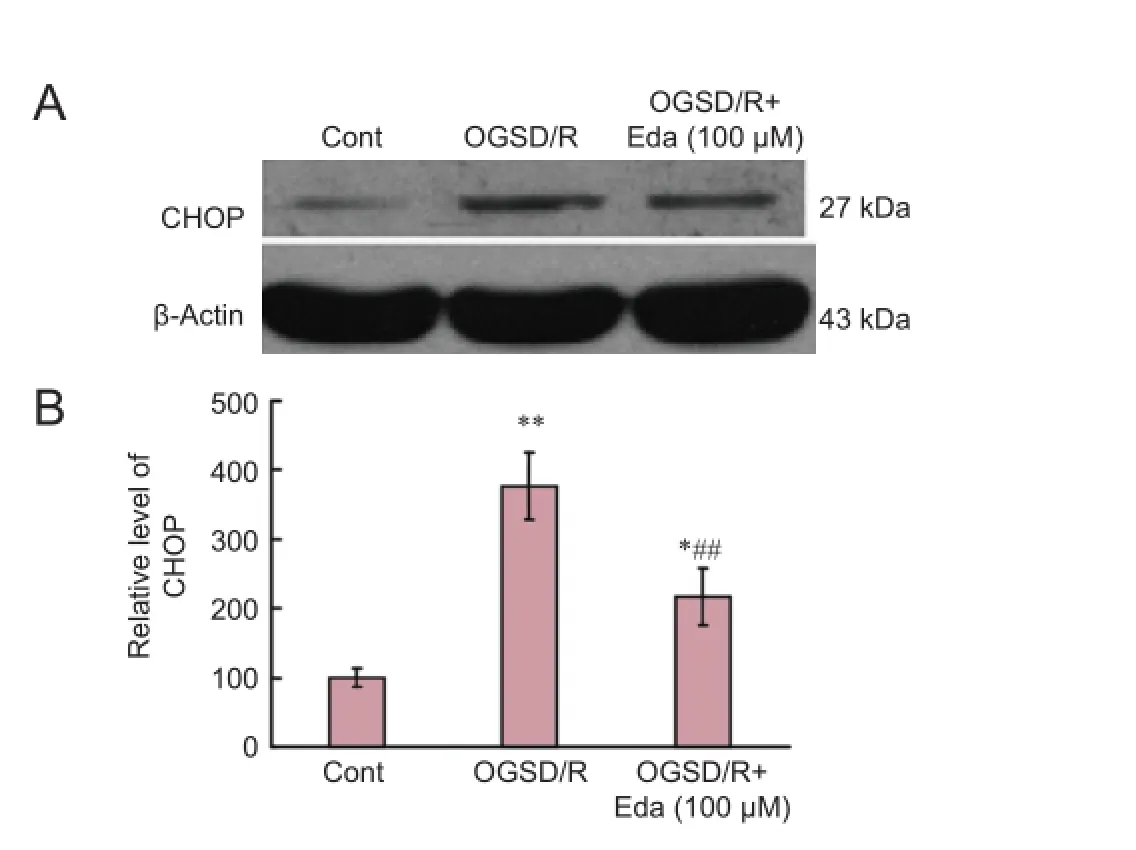
Figure 5 Ef f ects of edaravone (Eda) on the CCAAT/enhancer binding protein homologous protein (CHOP) signaling pathway after oxygen-glucose-serum deprivation/restoration (OGSD/R).
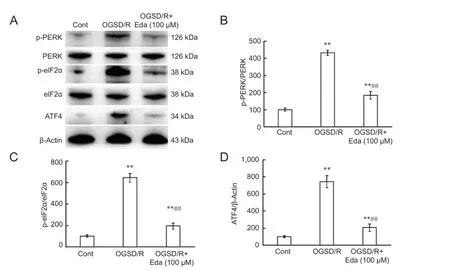
Figure 6 Ef f ects of edaravone (Eda) on the PERK/eIF2α/ATF4 signaling pathway after oxygen-glucose-serum deprivation/restoration (OGSD/R).
CHOP is a proapoptotic unfolded protein response gene and may play a role in the contribution of endoplasmic reticulum stress to myocardial ischemia, cerebral infarction, and acute kidney injury (Sano and Reed, 2013). During endoplasmic reticulum stress, caspase-12, an endoplasmic reticulum stress-associated apoptosis marker, dissociates from the endoplasmic reticulum membrane and is activated (Shibata and Kobayashi, 2008). Caspase-3 is the key protease in cell demolition during apoptosis (Boland et al., 2013). H2O2could induce apoptosis in neuronal cells during endoplasmic reticulum stress by regulating the expression of CHOP, JNK, Bcl-2, Bax and Bim and the activation of caspase-12 (Ye et al., 2014). Medicarpin, a pterocarpan class of naturally occurring benzopyran furanobenzene compound, sensitizes myeloid leukemia cells to TNF-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through activation of the ROS-JNK-CHOP and caspase-3 pathways (Trivedi et al., 2014). Ilimaquinone, a cytoplasmic microtubule inhibitor, enhances the sensitivity of human colon cancer cells to TRAIL-induced apoptosis through ROS-ERK/p38 MAPK-CHOP-mediated upregulation of TRAIL receptor 1 (DR4) and TRAIL receptor 2 (DR5) expression (Do et al., 2014). In this study, we found that edaravone decreased the expression of CHOP, and the activation of caspase-12 and -3, in spinal cord astrocytes after OGSD/R.
In conclusion, edaravone suppresses OGSD/R-induced apoptosis of spinal cord astrocytes. It blocks ROS generation, CHOP expression, caspase-12 and -3 activation, and inhibits the PERK/eIF2/ATF4 signaling pathway. Our study highlights a potential new use for an established drug: the prevention of OGSD/R-mediated spinal cord astrocyte apoptosis.
Acknowledgments:This paper was greatly improved by critical comments from Dr. Y Zhu.
Author contributions:QRD and ALZ designed this study. BD, YXS, YJX, and SHH performed experiments. TY, HBS, DC, and JRW analyzed data. ALZ wrote the paper. All authors approved the fi nal version of the paper.
Conf l icts of interest:None declared.
Plagiarism check:This paper was screened twice using CrossCheck to verify originality before publication.
Peer review:This paper was double-blinded and stringently reviewed by international expert reviewers.
Bhandary B, Marahatta A, Kim HR, Chae HJ (2012) An involvement of oxidative stress in endoplasmic reticulum stress and its associated diseases. Int J Mol Sci 14:434-456.
Black JA, Sontheimer H, Waxman SG (1993) Spinal cord astrocytes in vitro: phenotypic diversity and sodium channel immunoreactivity. Glia 7:272-285.
Boland K, Flanagan L, Prehn JH (2013) Paracrine control of tissue regeneration and cell proliferation by Caspase-3. Cell Death Dis 4:e725.
Chen H, Wang S, Ding JH, Hu G (2008) Edaravone protects against MPP+ -induced cytotoxicity in rat primary cultured astrocytes via inhibition of mitochondrial apoptotic pathway. J Neurochem 106:2345-2352.
Chen L, Liu Y, Dong L, Chu X (2014) Edaravone protects human peripheral blood lymphocytes from gamma-irradiation-induced apoptosis and DNA damage. Cell Stress Chaperones 20:289-295.
Chen Q, Boire A, Jin X, Valiente M, Er EE, Lopez-Soto A, Jacob LS, Patwa R, Shah H, Xu K, Cross JR, Massague J (2016) Carcinoma-astrocyte gap junctions promote brain metastasis by cGAMP transfer. Nature 533:493-498.
Cheng XL, Zhang JJ (2014) Ef f ect of edaravone on apoptosis of hippocampus neuron in seizures rats kindled by pentylenetetrazole. Eur Rev Med Pharmacol Sci 18:769-774.
Do MT, Na M, Kim HG, Khanal T, Choi JH, Jin SW, Oh SH, Hwang IH, Chung YC, Kim HS, Jeong TC, Jeong HG (2014) Ilimaquinone induces death receptor expression and sensitizes human colon cancer cells to TRAIL-induced apoptosis through activation of ROS-ERK/p38 MAPK-CHOP signaling pathways. Food Chem Toxicol 71:51-59.
Edaravone Acute Infarction Study G (2003) Ef f ect of a novel free radical scavenger, edaravone (MCI-186), on acute brain infarction. Randomized, placebo-controlled, double-blind study at multicenters. Cerebrovasc Dis 15:222-229.
Handa Y, Kaneko M, Takeuchi H, Tsuchida A, Kobayashi H, Kubota T (2000) Ef f ect of an antioxidant, Ebselen, on development of chronic cerebral vasospasm after subarachnoid hemorrhage in primates. Surg Neurol 53:323-329.
Harding HP, Zhang Y, Ron D (1999) Protein translation and folding are coupled by an endoplasmic-reticulum-resident kinase. Nature 397:271-274.
Hukkelhoven CW, Steyerberg EW, Farace E, Habbema JD, Marshall LF, Maas AI (2002) Regional dif f erences in patient characteristics, case management, and outcomes in traumatic brain injury: experience from the tirilazad trials. J Neurosurg 97:549-557.
Kim HG, Kim YR, Park JH, Khanal T, Choi JH, Do MT, Jin SW, Han EH, Chung YH, Jeong HG (2014) Endosulfan induces COX-2 expression via NADPH oxidase and the ROS, MAPK, and Akt pathways. Arch Toxicol 89:2039-2050.
Lin CL (2015) Attenuation of endoplasmic reticulum stress as a treatment strategy against ischemia/reperfusion injury. Neural Regen Res 10:1930-1931.
Liu J, Zhang Y, Xu R, Du J, Hu Z, Yang L, Chen Y, Zhu Y, Gu L (2013) PI3K/Akt-dependent phosphorylation of GSK3beta and activation of RhoA regulate Wnt5a-induced gastric cancer cell migration. Cell Signal 25:447-456.
Liu Y, Connor JR (2015) From adaption to death: endoplasmic reticulum stress as a novel target of selective neurodegeneration? Neural Regen Res 10:1397-1398.
Lu PD, Harding HP, Ron D (2004) Translation reinitiation at alternative open reading frames regulates gene expression in an integrated stress response. J Cell Biol 167:27-33.
Ma K, Vattem KM, Wek RC (2002) Dimerization and release of molecular chaperone inhibition facilitate activation of eukaryotic initiation factor-2 kinase in response to endoplasmic reticulum stress. J Biol Chem 277:18728-18735.
Mizuno N, Takahashi T, Kusuhara H, Schuetz JD, Niwa T, Sugiyama Y (2007) Evaluation of the role of breast cancer resistance protein (BCRP/ABCG2) and multidrug resistance-associated protein 4 (MRP4/ABCC4) in the urinary excretion of sulfate and glucuronide metabolites of edaravone (MCI-186; 3-methyl-1-phenyl-2-pyrazolin-5-one). Drug Metab Dispos 35:2045-2052.
Okamura K, Tsubokawa T, Johshita H, Miyazaki H, Shiokawa Y (2014) Edaravone, a free radical scavenger, attenuates cerebral infarction and hemorrhagic infarction in rats with hyperglycemia. Neurol Res 36:65-69. Osada N, Kosuge Y, Ishige K, Ito Y (2010) Characterization of neuronal and astroglial responses to ER stress in the hippocampal CA1 area in mice following transient forebrain ischemia. Neurochem Int 57:1-7.
Ouyang YB, Xu L, Lu Y, Sun X, Yue S, Xiong XX, Gif f ard RG (2013) Astrocyte-enriched miR-29a targets PUMA and reduces neuronal vulnerability to forebrain ischemia. Glia 61:1784-1794.
Pabst M, Braganza O, Dannenberg H, Hu W, Pothmann L, Rosen J, Mody I, van Loo K, Deisseroth K, Becker AJ, Schoch S, Beck H (2016) Astrocyte intermediaries of septal cholinergic modulation in the hippocampus. Neuron 90:853-865.
Sano R, Reed JC (2013) ER stress-induced cell death mechanisms. Biochimica Biophys Acta 1833:3460-3470.
Shibata N, Kobayashi M (2008) The role for oxidative stress in neurodegenerative diseases. Brain Nerve 60:157-170.
Shindo A, Maki T, Mandeville ET, Liang AC, Egawa N, Itoh K, Itoh N, Borlongan M, Holder JC, Chuang TT, McNeish JD, Tomimoto H, Lok J, Lo EH, Arai K (2016) Astrocyte-derived pentraxin 3 supports blood-brain barrier integrity under acute phase of stroke. Stroke 47:1094-1100.
Trivedi R, Maurya R, Mishra DP (2014) Medicarpin, a legume phytoalexin sensitizes myeloid leukemia cells to TRAIL-induced apoptosis through the induction of DR5 and activation of the ROS-JNK-CHOP pathway. Cell Death Dis 5:e1465.
Turan M, Ciger E, Arslanoglu S, Borekci H, Onal K (2017) Could edaravone prevent gentamicin ototoxicity? An experimental study. Hum Exp Toxicol doi:10.1177/0960327116639360.
Wang GH, Jiang ZL, Li YC, Li X, Shi H, Gao YQ, Vosler PS, Chen J (2011) Free-radical scavenger edaravone treatment confers neuroprotection against traumatic brain injury in rats. J Neurotrauma 28:2123-2134.
Yang JW, Hu ZP (2015) Neuroprotective ef f ects of atorvastatin against cerebral ischemia/reperfusion injury through the inhibition of endoplasmic reticulum stress. Neural Regen Res 10:1239-1244.
Ye J, Han Y, Chen X, Xie J, Liu X, Qiao S, Wang C (2014) l-Carnitine attenuates HO-induced neuron apoptosis via inhibition of endoplasmic reticulum stress. Neurochem Int 78C:86-95.
Yoshida H (2007) ER stress and diseases. FEBS J 274:630-658.
Zhang A, Zhang J, Sun P, Yao C, Su C, Sui T, Huang H, Cao X, Ge Y (2010) EIF2alpha and caspase-12 activation are involved in oxygen-glucose-serum deprivation/restoration-induced apoptosis of spinal cord astrocytes. Neurosci Lett 478:32-36.
Copyedited by Slone-Murphy J, Frenchman B, Wang J, Li CH, Qiu Y, Song LP, Zhao M
How to cite this article: Dai B, Yan T, Shen YX, Xu YJ, Shen HB, Chen D, Wang JR, He SH, Dong QR, Zhang AL (2017) Edaravone protects against oxygen-glucose-serum deprivation/restoration-induced apoptosis in spinal cord astrocytes by inhibiting integrated stress response. Neural Regen Res 12(2):283-289.
Open access statement: This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Funding: This work was supported by a grant from the Science & Technology Bureau of Changzhou City of China, No. CJ20130029.
*Correspondence to:
Qi-rong Dong or
Ai-liang Zhang,
dqrsdefy@163.com or
ailiangzhang07@163.com.
orcid:
0000-0003-2916-5792
(Qi-rong Dong)
10.4103/1673-5374.199006
Accepted: 2017-01-25
- 中国神经再生研究(英文版)的其它文章
- Hyperbaric oxygen preconditioning improves postoperative cognitive dysfunction by reducing oxidant stress and inf l ammation
- Nonhuman primate models of focal cerebral ischemia
- The cortical activation pattern during bilateral arm raising movements
- Neuroprotective ef f ect of the Chinese medicine Tiantai No. 1 and its molecular mechanism in the senescence-accelerated mouse prone 8
- Adenyl cyclase activator forskolin protects against Huntington’s disease-like neurodegenerative disorders
- Ef f ect of electroacupuncture on the mRNA and protein expression of Rho-A and Rho-associated kinase II in spinal cord injury rats