
PP2A对转化生长因子β1诱导的淋巴瘤细胞凋亡的影响*
2017-03-02 03:00:17汪茂荣
华中科技大学学报(医学版) 2017年1期
赵 哲,刘 莉,刘 琨,姚 平,汪茂荣
湖北民族学院附属民大医院血液内科,恩施 445000
PP2A对转化生长因子β1诱导的淋巴瘤细胞凋亡的影响*
赵 哲,刘 莉,刘 琨,姚 平△,汪茂荣
湖北民族学院附属民大医院血液内科,恩施 445000
目的 探讨转化生长因子β1(TGF-β1)诱导B细胞淋巴瘤细胞株Raji细胞凋亡的效应及机制。方法 采用TGF-β1(5 ng/mL)刺激Raji细胞,流式细胞术检测0、24、48、72 h细胞凋亡情况;定量PCR法检测Bcl-2、Bcl-xl、Caspase-3 mRNA表达;Western blot法检测Bcl-2,Bcl-xl以及Caspase-3蛋白表达及细胞外信号调节激酶ERK1/2磷酸化情况;蛋白磷酸酶活性测定体系检测TGF-β1刺激下蛋白磷酸酶2A(PP2A)活性变化;观察不同浓度PP2A抑制剂冈田酸(okadaic acid,OA)对TGF-β1诱导的细胞凋亡的干预效应。结果 5 ng/mL TGF-β1刺激24、48、72 h后细胞凋亡率分别为(29±3)%、(47±4)%、(60±5)%,差异有统计学意义(均P<0.05);同时,随着刺激时间延长,Bcl-2、Bcl-xl mRNA和蛋白表达逐渐降低,Caspase-3 mRNA和蛋白表达逐渐增加,差异有统计学意义(均P<0.05);ERK1/2总蛋白表达无显著差异,其磷酸化状态p-ERK1/2随TGF-β1刺激时间的延长逐渐减少;PP2A活性在TGF-β1干预15 min后开始升高,1 h达峰值;100、200 nmol/L的OA分别预处理后细胞凋亡率明显下降,TGF-β1组为(60±5)%,干预组分别为(32±4)%、(24±3)%,差异有统计学意义(均P<0.05)。结论 TGF-β1可能通过活化PP2A、抑制ERK1/2通路相关基因的表达而诱导B细胞淋巴瘤Raji细胞凋亡。
转化生长因子β1; 伯基特淋巴瘤; 细胞凋亡; 蛋白磷酸酶2A
转化生长因子β1(transforming growth factor β1,TGF-β1)是一种广泛参与并调节机体细胞增殖、分化和凋亡等功能的重要细胞因子[1-2]。在肿瘤中,TGF-β1具有双向调控作用,既能够抑制肿瘤细胞的增殖,又能够促进肿瘤的发生发展[3]。关于TGF-β1诱导的肿瘤细胞凋亡和生长抑制作用已经在多种细胞中得到验证,包括人和小鼠淋巴细胞以及多种淋巴瘤细胞系[4]。由于淋巴瘤细胞本身可以产生TGF-β1,因此,多种淋巴恶性肿瘤细胞对内源性TGF-β1敏感性降低。但是,通过外源性增加TGF-β1剂量仍然可以诱导淋巴瘤细胞凋亡[5]。然而,目前对于TGF-β1诱导淋巴瘤细胞凋亡的机制仍不十分清楚。本研究以Raji细胞为研究对象,观察TGF-β1诱导淋巴瘤细胞凋亡并探索其可能的诱导机制。
1 材料和方法
1.1 主要材料和仪器
B细胞淋巴瘤细胞株Raji细胞(中国科学院上海细胞生物学研究所细胞库),RPMI 1640培养液、胎牛血清(FBS,美国Gibco),RT-PCR试剂盒(美国Fermentas),PP2A磷酸酶活性检测试剂盒(美国Promega),人重组TGF-β1(美国Peprotech),Annexin Ⅴ-FITC/PI双染法细胞凋亡检测试剂盒(美国BD公司),兔抗人Bcl-2(美国Abcam)、兔抗人Bcl-xl(美国BD)、兔抗人Caspase-3(美国R&D)、小鼠抗人GAPDH(美国Epitomic)、兔抗人磷酸化p-ERK1/2(美国CST)、兔抗人ERK1/2(美国CST)、辣根过氧化物酶(HRP)标记的山羊抗兔/鼠IgG(美国Abcam),冈田酸(okadaic acid,OA,美国Sigma)。流式细胞仪(美国BD公司),凝胶成像和化学发光分析系统(美国Alpha FlurochemTM公司)。
1.2 细胞培养及分组
Raji细胞用含10%FBS的RPMI 1640培养液培养于37℃、5%CO2、95%空气湿度培养箱中培养。隔日换液,每3天传代1次。取对数生长期的细胞进行相关实验。在给予TGF-β1刺激前换用无血清培养液培养细胞12 h使同步化。实验分组:对照组(Control)、TGF-β1刺激组(24、48、72 h)、100 nmol/L OA+ TGF-β1刺激组、200 nmol/L OA+ TGF-β1刺激组。无血清培养同步化后,OA预处理1 h后再给予TGF-β1刺激。
1.3 流式细胞术检测细胞凋亡
根据BD公司提供的Annexin Ⅴ-FITC/PI试剂盒说明书,收集各组细胞,PBS洗涤2次,离心;然后用250 μL结合缓冲液重悬细胞并使其密度为(4~5)×105/mL,加入5 μL Annexin Ⅴ-FITC及2 μL PI,混匀后室温避光孵育15 min,离心收集细胞,用500 μL结合缓冲液冲洗细胞1次,再次离心后用500 μL缓冲液重悬,以流式细胞仪检测分析。每次实验设2个复孔,实验重复3次。
1.4 实时荧光定量PCR法检测Bcl-2、Bcl-xl、Caspase-3mRNA表达
按照Trizol试剂盒说明书提取各组细胞总RNA并逆转录成cDNA,然后按以下条件进行PCR。Bcl-2引物:上游5′-TCGCCCTGTGGATGACTGA-3′,下游5′-GCATCCCAGCCTCCGTTATC-3′,片段长度189 bp;Bcl-xl引物:上游5′-GAGCTGGTGGTTGACTTTCTC-3′,下游5′-TCCATCTCCGATTCAGTCCCT-3′,片段长度119 bp;Caspase-3引物:上游5′-GATTATCCTGA-GATGGGTTTATGTAT-3′,下游5′-ATCACGC-ATCAATTCCACAAT-3′,片段长度124 bp;β-actin引物:上游5′-TCCTCCTGAGCGCAAGTA-CTCT-3′,下游5′-GCTCAGTAACAGTCCGCCT-AGAA-3′,片段长度153 bp。PCR反应条件:94℃预变性5 min,94℃变性30 s、55℃30 s、72℃30 s,共32个循环,72℃延伸10 min。以β-actin为内参,应用相对定量法2-ΔΔCt进行定量分析。每次实验设2个复孔,实验重复4次。
1.5 Western blot检测蛋白表达
RIPA裂解并收集各组细胞,BCA法测定蛋白浓度。取蛋白20 g/孔,在含SDS-PAGE中电泳分离后将蛋白转移至PVDF膜上;PVDF膜经5%脱脂牛奶(1×TBST缓冲液配制)室温封闭1 h,与一抗(Bcl-2、Bcl-xl、Caspase-3、GAPDH、p-ERK1/2、ERK1/2)4℃孵育过夜。1×TBST洗膜(10 min×3次),与辣根过氧化物酶偶联的二抗37℃摇床孵育1 h。经1×TBST洗涤3次后用ECL显色曝光。
1.6 PP2A磷酸酶活性测定
采用Promega磷酸酶活性测定标准程序,Raji细胞经TGF-β10、15、30、60 min,2、24、48、72 h刺激后采用磷酸酶存储缓冲区收集蛋白,离心(100 000 g,4℃,1 h);向核酸纯化柱中加入10 mL G-25,自身重力排干,加入10 mL 冷磷酸酶存储缓冲区洗涤核酸纯化柱,自身重力排干,600 g、4℃、离心5 min;加250 μL蛋白样品入核酸纯化柱,用50 mL离心管接住,600 g、4℃、离心5 min;制作标准曲线:取原液20 μL(浓度1 mmol/L),用无磷水稀释至200 μL,使磷酸盐浓度50 μmol/L,共6个点,每孔加0、2、4、10、20、40 μL,用1×反应缓冲液补齐至50 μL,每样品孔加入5 μL磷酸肽+10 μL 5×反应缓冲液+35 μL上清,37℃孵育30 min,每孔加50 μL钼酸染料,加色法混合终止反应,室温孵育15 min后于630 nm波长处测吸光度值。
1.7 统计学分析
计量资料数据用均数±标准差表示,组间均数比较采用单因素方差分析和t检验。所有实验均重复至少3次。以P<0.05为差异有统计学意义。采用SPSS 19.0软件进行统计学分析。
2 结果
2.1 TGF-β1对Raji细胞凋亡的影响
采用流式细胞术检测TGF-β1不同浓度及不同刺激时间下Raji细胞凋亡情况。1、5、10 ng/mL TGF-β1均能诱导Raji细胞凋亡(均P<0.05)。和5 ng/mL浓度相比,10 ng/mL浓度的TGF-β1作用后凋亡细胞百分率差异无统计学意义(P>0.05)。因此,选择5 ng/mL TGF-β1为凋亡诱导剂量(图1A)。图1B显示,Raji细胞的凋亡率随着TGF-β1作用时间的延长而增加,与对照组比较差异均有统计学意义(均P<0.05)。
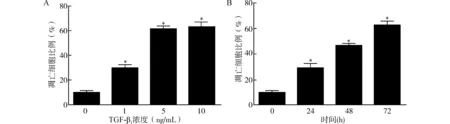
无TGF-β1刺激为对照组;与对照组比较,*P<0.05图1 TGF-β1不同浓度和不同时间刺激下细胞凋亡情况Fig.1 Cell apoptosis under stimulation with TGF-β1 of different concentrations and different time
2.2 TGF-β1对Raji细胞Bcl-2、Bcl-xl、Caspase-3表达的影响
为了明确TGF-β1诱导Raji细胞凋亡效应,我们检测了TGF-β1处理前后细胞中Bcl-2、Bcl-xl以及Caspase-3基因表达的变化趋势。RT-PCR检测结果显示,与对照组比较,5 ng/mL TGF-β1作用于Raji细胞,Bcl-2、Bcl-xl基因表达水平随着作用时间的延长而降低,具有时间依赖性(图2A、2B);而Caspase-3基因表达水平随着作用时间的延长逐渐增加(图2C),差异具有统计学意义(均P<0.05)。Western blot检测结果也表明,TGF-β1刺激后Raji细胞Bcl-2、Bcl-xl蛋白随着刺激时间的延长表达逐渐下降,Caspase-3蛋白表达逐渐增多(图3)。
2.3 TGF-β1对Raji细胞ERK1/2总蛋白及其磷酸化水平的影响
Western blot检测各组ERK1/2总蛋白水平,结果显示,和对照组比较,TGF-β1刺激对ERK1/2总蛋白表达水平无明显影响,而其磷酸化蛋白p-ERK1/2随着刺激时间的延长表达逐渐下降(图3),差异有统计学意义(均P<0.05)。
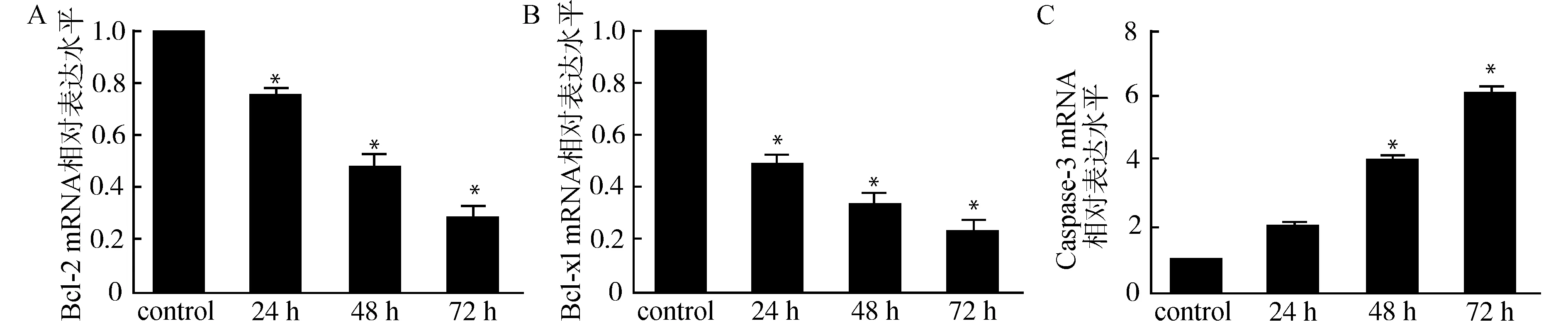
与control组比较,*P<0.05图2 TGF-β1对Raji细胞Bcl-2(A)、Bcl-xl(B)以及Caspase-3(C) mRNA表达的影响Fig.2 Effect of TGF-β1 on mRNA expression of Bcl-2(A),Bcl-xl(B) and Caspase-3(C)
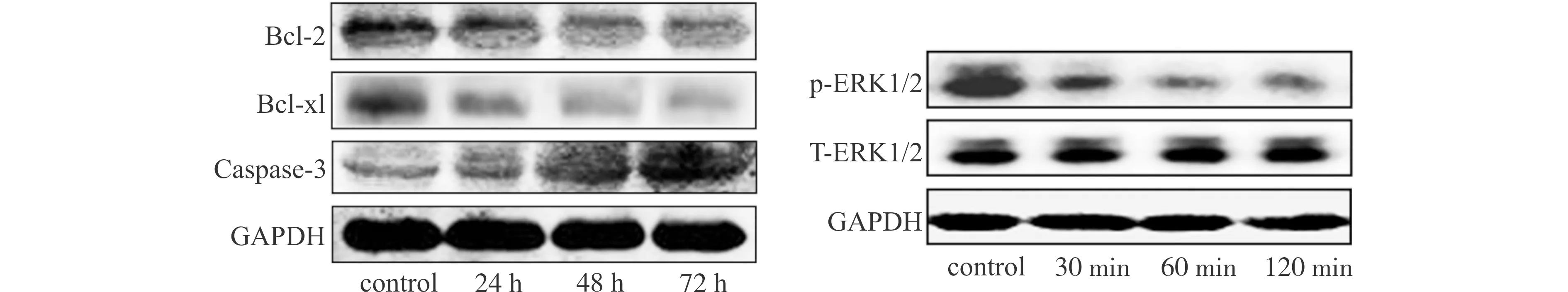
图3 Western blot法检测TGF-β1对Raji细胞Bcl-2、Bcl-xl、Caspase-3以及ERK1/2蛋白表达的影响Fig.3 Effect of TGF-β1 on protein expression of Bcl-2,Bcl-xl,Caspase-3 and ERK1/2 detected by Western blotting
2.4 TGF-β1诱导Raji细胞凋亡过程中PP2A活性变化
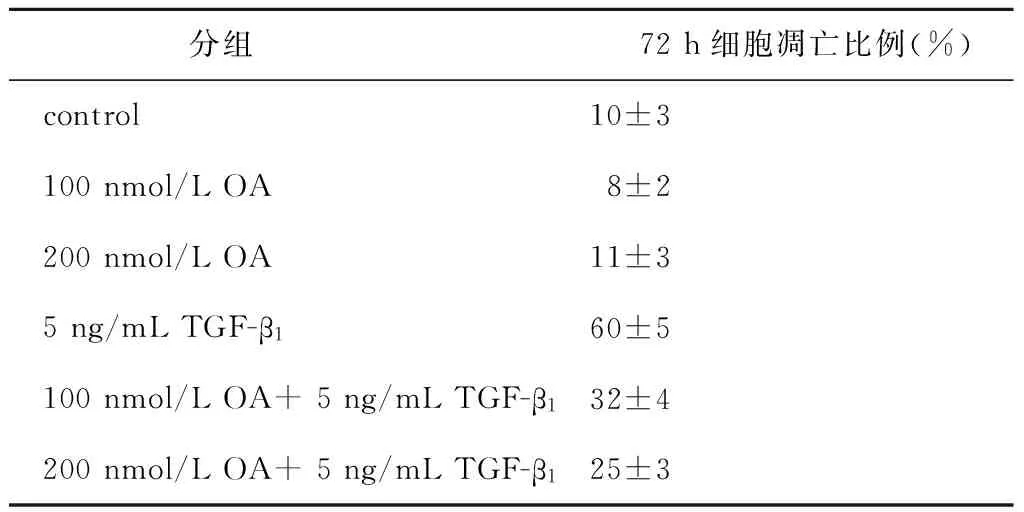
TGF-β1刺激Raji细胞后,于不同刺激时间点收集细胞蛋白,观察细胞总蛋白磷酸酶活性的改变。结果显示,丝/苏氨酸磷酸酶活性于刺激后15 min开始上升,1 h时达到峰值,随后逐渐下调,差异具有统计学意义(均P<0.05)(图4A)。采用PP2A特异性活性抑制剂OA预处理Raji细胞1 h后,给予TGF-β1刺激72 h,采用Western blot法检测不同浓度OA干预下细胞内Bcl-2、Bcl-xl以及Caspase-3蛋白表达变化。结果显示,OA干预组与TGF-β1组相比,Bcl-2、Bcl-xl表达显著上调,而Caspase-3表达显著下调(图4B),并且随着OA浓度的增加,干预效应有增强趋势。为进一步明确OA的干预效应,采用流式细胞术检测凋亡细胞百分率。如表1所示,Raji细胞自然凋亡率为(10±3)%,单纯使用OA对细胞凋亡率无显著影响,差异无统计学意义(均P>0.05);TGF-β1刺激可显著诱导细胞凋亡,其凋亡率为(60±5)%,和对照组相比,差异具有统计学意义(P<0.05);采用100、200 nmol/L浓度OA预处理1 h后,可显著抑制TGF-β1对Raji细胞诱导凋亡的作用,其凋亡率分别降至(32±4)%、(25±3)%,和TGF-β1组相比,差异具有统计学意义(均P<0.05)。
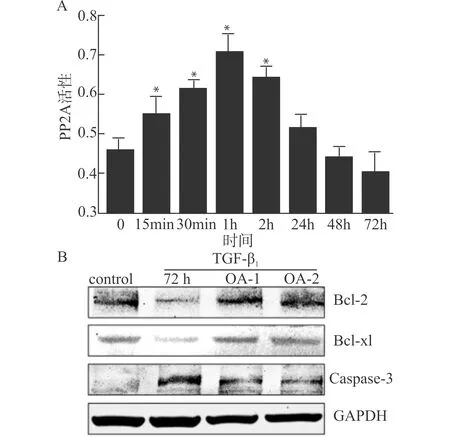
A:PP2A活性随TGF-β1刺激时间变化;B:OA干预对TGF-β1刺激蛋白表达的影响;OA-1:100 nmol/L预处理1 h;OA-2:200 nmol/L预处理1 h;与刺激0min比较,*P<0.05图4 PP2A在TGF-β1诱导Raji细胞凋亡过程中的活性变化及作用Fig.4 Activity change and the role of PP2A in Raji cell apoptosis induced by TGF-β1

分组 72h细胞凋亡比例(%)control10±3100nmol/LOA8±2200nmol/LOA11±35ng/mLTGF-β160±5100nmol/LOA+5ng/mLTGF-β132±4200nmol/LOA+5ng/mLTGF-β125±3
3 讨论
TGF-β作为一种重要的细胞因子,广泛参与胚胎发育、创伤愈合及炎症反应等一系列生理或病理生理过程。TGF-β信号转导包含多种配体与受体的结合。在哺乳动物体内,存在TGF-β1、TGF-β2与TGF-β3等3种配体。而TGF-β受体为受体丝氨酸/苏氨酸激酶两者形成的异源二聚体,包括TβRⅠ、TβRⅡ、TβRⅢ等3种[6]。研究表明,TGF-β信号通路与肿瘤发生密切相关,它既可以作为肿瘤促进因子,通过增强侵袭性质、影响肿瘤微环境及抑制免疫功能等促进肿瘤转移[7],也可以作为肿瘤抑制因子,抑制肿瘤细胞增殖,促进肿瘤细胞凋亡[8]。因此,TGF-β信号通路在不同肿瘤、同种肿瘤不同亚型及肿瘤的不同阶段均可产生不同反应,并呈现不同调控机制[9]。研究发现,恶性淋巴肿瘤细胞对内源性TGF-β敏感性降低,但是,通过外源性增加TGF-β剂量仍然可以诱导肿瘤细胞凋亡[5,10]。TGF-β诱导调节肿瘤细胞凋亡的机制根据细胞类型的不同而变化。除TGF-β-Smad这一经典的诱导凋亡途径之外[11-12],非Smad途径,如Daxx-JNK的活化也可能参与其中[13]。通过上调促凋亡因子的表达(如Bax)和下调抑制凋亡因子的表达(如Bcl-2、Bcl-xl),诱导产生凋亡信号并激活Caspase家族[14]。Caspase-3被认为是凋亡的关键蛋白酶,一旦被激活,即发生下游的级联反应,使凋亡不可避免。
蛋白磷酸酶2A(PP2A)是真核生物体内最为重要的蛋白磷酸酶之一,占细胞总蛋白的1%,在多个信号转导途径中起着重要的调节作用,包括细胞周期调控、细胞增殖、分化以及细胞凋亡[15]。同时,PP2A作为一种重要的肿瘤抑制因子,广泛参与肿瘤细胞的发生和发展。大量研究表明,在多种实体瘤和血液系统肿瘤性疾病中,存在磷酸酶和磷酸激酶的不平衡状态,表现为PP2A磷酸酶活性受到抑制,肿瘤性激酶异常活化,最终引起肿瘤的发生和发展[16]。越来越多的研究证实PP2A在肿瘤中的作用,并且研发出以PP2A为作用靶点的抗肿瘤药物。例如,FTY720是近年来研制出的一种PP2A激活剂,通过激活PP2A,保留其磷酸酶活性,对肿瘤的发生发展具有有效的拮抗作用[17]。相反,采用PP2A特异性抑制剂OA抑制PP2A活性后,能够促进肿瘤的发生[18]。
PP2A通过调节细胞分裂和凋亡信号来发挥肿瘤抑制功能。PP2A的凋亡功能可通过直接去磷酸化一些特异的蛋白来发挥作用,可以同时使抗凋亡蛋白Bcl-2失活而激活凋亡诱导因子Bad;另外,PP2A还可以直接去磷酸化AKT来影响FOX的转录和凋亡活性[19]。在本研究中,我们以B细胞淋巴瘤Raji细胞为研究对象,发现PP2A还能通过去磷酸化ERK1/2发挥促凋亡作用。ERK是丝裂原活化蛋白激酶(MAPK)家族中的一员,受活化的MEK蛋白激酶磷酸化和蛋白磷酸酶去磷酸化作用,前者通过磷酸化激活ER,后者则去磷酸化使其失活。研究发现,磷酸化的ERK(p-ERK)具有抑制肿瘤细胞凋亡的作用。当p-ERK蛋白从细胞质转入到细胞核后,可促进促凋亡基因的表达(如Bad、Bim)和抑制抗凋亡基因的表达(如Bcl-2、Bcl-x1),最终可激活与凋亡相关的信号途径,如Caspase-3表达上调[20]。因此,PP2A通过去磷酸化ERK,降低p-ERK在肿瘤细胞中的表达,最终达到促进肿瘤细胞凋亡的作用。
[1] Bierie B,Moses H L.Tumour microenvironment:TGF-β:the molecular Jekyll and Hyde of cancer[J].Nat Rev Cancer,2006,6(7):506-520.
[2] Elliott R L,Blobe G C.Role of transforming growth factor β in human cancer[J].J Clin Oncol,2005,23(9):2078-2093.
[3] Miyazono K,Suzuki H,Imamura T,et al.Regulation of TGF-β signaling and its roles in progression of tumors[J].Cancer Sci,2003,94(3):230-234.
[4] Saltzman A,Munro R,Searfoss G,et al.Transforming growth factor-β-mediated apoptosis in the ramos B-lymphoma cell line is accompanied by caspase activation and Bcl-xl downregulation[J].Exp Cell Res,1998,242(1):244-254.
[5] Inman G J,Allday M J.Apoptosis induced by TGF-β1in Burkitt’s lymphoma cells is caspase 8 dependent but is death receptor independent[J].J Immunol,2000,165(5):2500-2510.
[6] Massagué J.TGF-β signalling in context[J].Nat Rev Mol Cell Biol,2012,13(10):616-630.
[8] Levy L,Hill C S.Smad4 dependency defines two classes of transforming growth factor β(TGF-β)target genes and distinguishes TGF-β-induced epithelial-mesenchymal transition from its antiproliferative and migratory responses[J].Mol Cell Biol,2005,25(18):8108-8125.
[9] Akhurst R J,Hata A.Targeting the TGF-β signalling pathway in disease[J].Nat Rev Drug Discov,2012,11(10):790-811.
[10] Tótth A,Sebestyén A,Barna G,et al.TGF-β1induces caspase-dependent but death-receptor independent apoptosis in lymphoid cells[J].Anticancer Res,2001,21(2A):1207-1212.
[11] Yamamura Y,Hua X,Bergelson S,et al.Critical role of smads and AP-1 complex in transforming growth factor-β-dependent apoptosis[J].J Biol Chem,2000,275(46):36295-36302.
[12] Lallemand F,Mazars A,Prunier C,et al.Smad7 inhibits the survival nuclear factor-κB and potentiates apoptosis in epithelial cells[J].Oncogene,2001,20(7):879-884.
[13] Perlman R,Schiemann W P,Brooks M W,et al.TGF-β-induced apoptosis is mediated by the adapter protein Daxx that facilitates JNK activation[J].Nat Cell Biol,2001,3(8):708-714.
[14] Motyl T,Grzelkowska K,Zimowska W,et al.Expression of bcl-2 and box in TGF-β1-induced apoptosis of L1210 leukemic cells[J].Eur J Cell Biol,1998,75(4):367-374.
[15] Mumby M.PP2A:Unveiling a reluctant tumor suppressor[J].Cell,2007,130(1):21-24.
[16] Eichhorn P J,Creyghton M P,Bernards R.Protein phosphatase 2A regulatory subunits and cancer[J].Biochim Biophys Acta,2009,1795(1):1-15.
[17] Perrotti D,Neviani P.Targeting a tumor suppressor to suppress tumor growth:news and views on protein phosphatase 2A(PP2A)as a target for anti-cancer therapy[J].Lancet Oncol,2013,14(3):e229-e238.
[18] Fujiki H,Suganuma M.Tumor promotion by inhibitors of protein phosphatases 1 and 2A:the okadaic acid class of compounds[J].Adv Cancer Res,1993,61(9):143-194.
[19] Janssens V,Rebollo A.The role and therapeutic potential of Ser/Thr phosphatase PP2A in apoptotic signalling networks in human cancer cells[J].Curr Mol Med,2012,12(3):268-287.
[20] 冯可,王超,周虎,等.ERK1/2抑制剂AZD8330对Burkitt淋巴瘤细胞株Raji细胞的作用及其机制研究[J].中华血液学杂志,2015,36(2):148-152.
(2016-09-05 收稿)
PP2A-dependent Apoptotic Effect of Exogenous Transforming Growth Factor-β1on Human Burkitt’s Lymphoma Cell Line Raji Cells and Its Mechanism
Zhao Zhe,Liu Li,Liu Kunetal
DepartmentofHematology,MindaHospitalofHubeiUniversityforNationalities,Enshi445000,China
Objective To study the apoptotic effect of exogenous TGF-β1on human Burkitt’s lymphoma cell line Raji cells and its mechanism.Methods Raji cells were treated with recombinant TGF-β1at different concentrations(0,1,5,10 ng/mL)or stimulated by TGF-β1for different time(0,24,48,72 h).The apoptosis rate of Raji cells was detected by flow cytometry.Gene expression levels of Bcl-2,Bcl-xl and Caspase-3 were assayed by real-time PCR.Western blot analysis was used to assay the protein expression level of Bcl-2,Bcl-xl,Caspase-3,ERK1/2 and p-ERK1/2.Protein Ser/Thr phosphatase activity was assayed photometrically using the Serine/Threonine Phosphatase Assay System according to the manufacturer’s directions.Results Cell apoptosis rate was (29±3)%,(47±4)% and (60±5)%,24,48 and 72 h after stimulation of 5 ng/mL TGF-β1(allP<0.05).Simultaneously,mRNA and protein expression levels of Bcl-2 and Bcl-xl were decreased,and those of Casepase-3 were increased with time.Total protein expression of ERK1/2 was not significantly different,but p-ERK1/2 was decreased with TGF-β1stimulation.After pretreatment of 100 and 200 nmol/L okadaic acid(OA),cell apoptosis rate was significantly decreased [(60±5)%vs.(32±4)% and (24±3)%;bothP<0.05].Conclusion TGF-β1induces Raji cell apoptosis by the activation of PP2A and down-regulation of ERK1/2 signal pathway related genes.
transforming growth factor β1; Burkitt’s lymphoma; apoptosis; PP2A
*湖北省教育厅科研项目(No.B2015101)
赵 哲,男,1979年生,主治医师,硕士研究生,E-mail:1047687209@qq.com
△通讯作者,Corresponding author,E-mail::yaoping0564@126.com
R733.4
10.3870/j.issn.1672-0741.2017.01.012
猜你喜欢
传染病信息(2022年3期)2022-07-15 08:24:12
家庭医学(下半月)(2019年11期)2020-01-16 08:39:08
天津医科大学学报(2019年6期)2019-08-13 07:04:42
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
广西林业科学(2016年1期)2016-03-20 05:33:00
磁共振成像(2015年5期)2015-12-23 08:52:50
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
医学研究杂志(2015年8期)2015-06-22 14:00:57
天津医科大学学报(2015年3期)2015-06-05 12:21:49
遗传(2014年3期)2014-02-28 20:59:01
