
闽北闽粤栲天然林主要树种幼树器官碳氮磷化学计量特征分析
2017-02-23 07:43:38郑德祥蔡杨新杨玉洁钟兆全缪三华吴文斌
林业科学研究 2017年1期
郑德祥,蔡杨新,杨玉洁,钟兆全,缪三华,吴文斌
(1.福建农林大学林学院,福建 福州 350002; 2.顺昌国有林场,福建 顺昌 353200)
闽北闽粤栲天然林主要树种幼树器官碳氮磷化学计量特征分析
郑德祥1*,蔡杨新1,杨玉洁1,钟兆全2,缪三华1,吴文斌1
(1.福建农林大学林学院,福建 福州 350002; 2.顺昌国有林场,福建 顺昌 353200)
[目的]选取4种主要优势树种幼树为研究对象,分析其不同器官中C、N、P的生态化学计量特征,为闽粤栲天然林的更新与保护提供理论参考。[方法]应用碳氮分析仪与HNO3-HClO4消煮法分别测定幼树的干、根、枝、叶中的C、N、P含量的质量分数,在不同器官C、N、P含量差异分析的基础上,通过化学计量比变化的对比分析研究幼树不同器官的C、N、P化学计量特征。[结果]闽北闽粤栲天然林主要优势树种幼树叶片中各元素的含量均高于其他器官,而根与干中的营养元素含量较低;不同器官的C∶P远高于C∶N与N∶P,其中,N∶P最小且因树种不同而有所变化,不同器官的C、N、P含量及化学计量特征均差异极显著。[结论]闽北闽粤栲天然林主要优势树种不同器官对C、N、P的选择与吸收存在差异,其叶片C含量最高且高于全球尺度和全国尺度,但N、P含量较低;幼树不同器官的C∶N、C∶P高于平均水平,而N∶P>16;闽粤栲天然林主要优势树种幼树对养分的利用效率与生长速率均较低,幼树的生长更新主要受N、P元素含量的影响,并可能受到P元素缺失的制约。
天然林;幼树;器官;营养元素;化学计量;闽粤栲
生态化学计量学是一门结合生物学、化学和物理学基本原理的综合学科[1],该理论认为元素是构成有机体的主要成分,并对有机体的许多行为进行有序调控[2-3]。2000年Elser等[4]明确提出生态化学计量学的概念,2002年,Sterner和Elser出版专著《Ecological Stoichimetry: The Biology of Elements from Molecules to the Biosphere》,成为生态化学计量学领域的第一部巨作,标志着生态化学计量学理论的确立与形成。在国外,生态化学计量学的研究十分活跃[5-7],国内对其研究起步则相对较晚,但已成为生态学研究的热点问题之一。生态化学计量学研究从最初的水生生物生态系统逐渐拓展到对陆地植物生态系统的研究,近十余年来,诸多植物的生态化学计量特征研究见于报道,其中,又以森林生态系统和草原生态系统的研究成果居多。施家月等[8]对天童山39种常见植物幼树各器官的氮磷养分特征进行了研究,发现不同器官吸收N和P存在正相关关系。常云妮等[9]在尤溪县的天然米槠林中分别对乔木层、灌木层和草本层的26个优势种的碳氮磷化学计量特征进行了研究,结果表明,尤溪县天然米槠(Castanopsiscarlesii(Hemsl.) Hayata)林内植物叶片C、N、P含量均偏低,P是限制该林分植物生产力最重要的元素。陈亚南等[10]结合纬度和坡向2个相关因素,对陕西黄土高原刺槐(RobiniapseudoacaciaLinn.)枯落叶的C、N、P生态化学计量特征进行研究,表明陕西黄土高原刺槐林的生长主要受氮素限制。这些研究集中体现了植物体对土壤C、N、P元素的吸收与转化及土壤C、N、P对植物生长发育具有的重要作用。幼树阶段是植物生活史中对环境最敏感的时期,幼树体内的养分含量在一定程度上反映了所处环境的养分条件[8]。栗忠飞等[11]、石贤萌等[12]分别对西双版纳热带雨林及哀牢山的幼树生态化学计量特征进行研究,发现幼树叶中C、N、P的生态化学计量比对海拔变化的响应较为显著,环境中N元素的变化影响幼苗对P元素的吸收,且不同物种对外源N添加具有差异性利用策略。幼苗根和茎中的N内稳性比叶片的更高,叶片对N沉降的响应更敏感。幼树体内的C、N、P养分含量变化直接影响其生长发育,改变幼树的生长发育速率,最终决定其在群落中的更新及生态适应策略。
闽粤栲(Castanopsisfissa(Champ. ex Benth.) Rehd. et Wils.)为壳斗科(Fagaceae)栲属(CastanopsisSpach)常绿乔木,中性偏阳的深根性树种,是营造水土保持林、水源涵养林、薪炭林及用材林的优良树种,也是南方集体林区最常见的主要阔叶树种之一。苏凌燕等[9]对闽粤栲林隙特征及干扰状态进行了研究,蓝文升等[12]对闽粤栲的空间结构、种群分布格局进行了研究,管大跃等[14]研究了闽粤栲天然林的生物量并构建了生物量预测模型,但对闽粤栲幼树幼苗更新及其生态化学计量研究未见报道。本研究以闽粤栲天然林中4种优势树种幼树为对象,对幼树各器官的养分含量进行研究,揭示闽粤栲天然林幼树C、N、P的生态化学计量特征,从而为闽粤栲天然林的更新与保护提供理论参考。
1 研究区概况
2 研究方法
2.1 野外调查及取样
于2014年7月,在顺昌县国有林场下属的高阳采育场经营区内,选择保护良好的闽粤栲天然林林分进行调查。在阴坡和阳坡的林分中分别设立1块50 m ×50 m的标准地。参考李俊清编著的《森林生态学》[16]中对森林植物群落垂直结构的划分标准,将林分分为乔木层、灌木层和草本层3个林层,分别对不同林层调查与记载各树种的种类、胸径(胸径小于5 cm的测量地径)、树高等因子。以林下选择样地上分布最多的闽粤栲、木荷、米槠和中华杜英4种林分优势树种幼树为研究对象,其幼树的概况见表1。4树种按幼树平均状况选取3株生长良好的幼树进行叶、枝、干、根取样,采集植株顶层成熟、健康叶片,当年新生枝条,高度位于植株1.3 m处的干及幼树植株的完整主根[17]为试验测定样本。

表1 样地中4种优势树种幼树概况Table 1 Average profiles of the four dominant young tree species in the sample plots
2.2 样品处理及测定
对采集的样品进行整理分类,包装好带回实验室,75 ℃烘干粉碎后封袋保存并做好标记,各样品重复测定3组, 1组空白为对照,植物C、N质量分数采用碳氮分析仪测定;P质量分数采用HNO3-HClO4消煮测定[18]。根据测定结果分析其 C、N、P 含量及化学计量特征。
3 结果与分析
3.1 主要优势树种幼树不同器官C、N、P含量
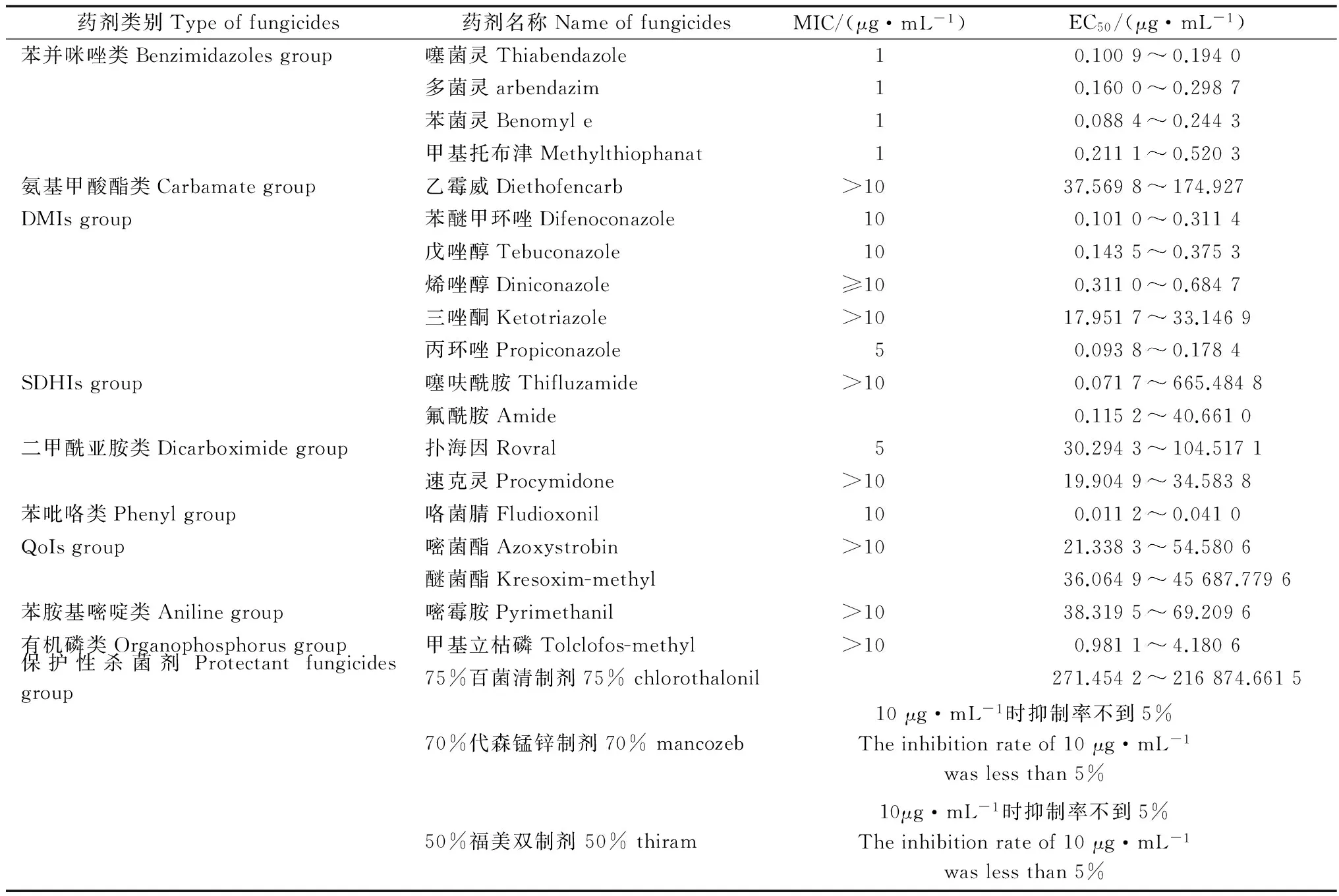
本研究分别测定闽北闽粤栲天然林林分中4种主要优势树种幼树不同器官的C、N、P含量,进而分析幼树各器官营养元素含量的差异性。测定结果(表2)表明:4种优势树种幼树叶片中各元素的含量均比其他器官的高,其中,叶片中的C含量最大,4种优势树种幼树不同器官的C含量变化均表现为叶>枝>干>根,N、P含量变化均表现为叶>枝>根>干。对不同器官各元素含量进行方差分析,结果(表3)表明:不同器官间各元素含量差异极显著(P<0.01)。

表2 4种优势树种幼树不同器官的C、N、P含量Table 2 Content of C, N and P in different organs of the four dominant young tree species

表3 4种优势树种幼树不同器官C、N、P含量 单因素方差分析结果Table 3 One-way ANOVA analysis results of the C, N and P content in different organs of the four dominant young tree species
3.2 4种优势树种幼树不同器官C、N、P的生态化学计量特征
为更好地说明闽粤栲天然林主要优势树种幼树的养分利用情况,进一步对幼树不同器官中的C、N、P的生态化学计量比进行分析,4种优势树种幼树不同器官C、N、P的生态化学计量特征(表4)表明:4种优势树种不同器官中的C∶P远高于C∶N与N∶P,C∶P由高到低依次为干>根>枝>叶,其次为C∶N,其排序与C∶P类似,N∶P最小且在不同树种中的排序有所变化,闽粤栲与米槠的N∶P由高到低依次为干>根>叶>枝,木荷的为干>叶>根>枝,中华杜英的为干>枝>根>叶。

表4 4种优势树种幼树不同器官C、N、P的 生态化学计量特征Table 4 Stoichiometry characteristics of C, N and P in different organs of the four dominant young tree species

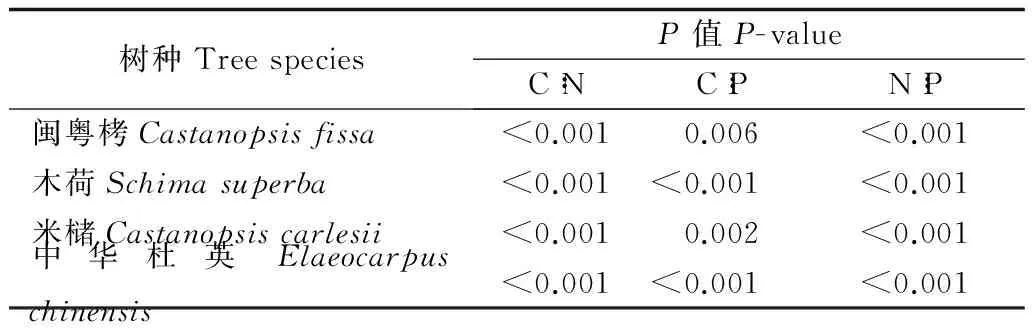
表5 4种优势树种幼树不同器官C、N、P生态化学 计量比单因素方差分析结果Table 5 One-way ANOVA analysis results of the stoichiometry ratio of C, N and P in different organs of the four dominant tree species
4 讨论
由于叶片是植物光合作用与代谢的主要场所,植物发育过程中将大量养分传递给叶片以满足植物生长的需要,幼树叶片中各营养元素含量最高,而幼树树干是高度木质化的器官,幼树根系分布尚浅,对吸收养分能力较弱。故而干和根中的营养元素含量较少,而枝作为联接树干和叶的主要营养传输器官,营养元素含量介于二者之间,因此,闽北闽粤栲天然林4种主要优势树种幼树叶的C、N、P元素含量最大,其次为枝,而后为干或根,该结果验证了Krieger等[19]的理论,认为木质部的养分大部分被运输至更需要的部位,植物的干和根的养分含量较低,也表明各优势树种幼树不同器官对C、N和P的吸收具有选择性与差异性,其结论也与常云妮等[9]对尤溪县米槠林内植物化学计量研究结果基本一致。在闽北闽粤栲天然林4种主要优势树种幼树不同器官中,叶片的C含量最高,平均为483.3 mg·g-1,大于全球492种陆生植物的平均C含量(464 mg·g-1)[4],也高于邻近地区同为壳斗科栲属的天然米槠林叶片的平均C含量(441.9 mg·g-1)[9];闽北闽粤栲天然林4种主要优势树种幼树叶片中的N平均含量(16.2 mg·g-1)低于全球N含量水平(18.34 mg·g-1)[20]和全国753种植物叶片的N含量(18.63 mg·g-1)[21],略低于中国东部南北样带654种植物叶片的N含量(17.55 mg·g-1)[22]。闽粤栲天然林4种主要优势树种幼树叶片的P含量为0.46 mg·g-1,明显低于全球水平及全国和中国东部南北样带的叶片P含量(1.42、1.21、1.28 mg·g-1)[20-22]。闽北闽粤栲天然林4种主要优势树种幼树叶片的N、P含量较低,其原因可能是由于该区域高温多雨和降水淋溶作用导致土壤中的N、P含量不足,间接导致了叶片中的N、P含量降低,幼树不同器官对各营养元素的选择与吸收存在差异,这将直接影响4种优势树种幼树的新陈代谢与更新。
幼树不同器官中的C、N、P化学计量比变化影响幼树生长速率,进而影响幼树的更新及生态适应策略[17]。闽粤栲天然林4种主要优势树种的C、N、P计量比在不同器官中的变化差异显著,由于较高的C含量及较低的N、P含量,导致不同器官的C∶N、C∶P均大于全球尺度内植物的C∶N和C∶P(22.5和232)[23],较高的C∶N与C∶P反映了幼树对营养的利用效率较低,植物体内可能由于N元素或P元素的缺失导致植物生长速率降低;而植物体内N∶P化学计量特征反映N和P 2种营养元素的供应状况和相对有效性[24],当植物N∶P<14时,植物生长主要受N限制;当N∶P>16时,植物生长主要受P限制;当14 影响植物N∶P化学计量特征的因素复杂且综合。在天童山常绿阔叶林中,不同物种间的N∶P差异较大,但在各种植物的演替过程中,N∶P总体变化特征基本一致[26]。中亚热带杉木人工林中,不同生活型植物的N∶P差异不显著[27]。因此,影响植物生态化学计量比的因素复杂多样,不同植物、不同群落的养分限制性受多种因素控制。西双版纳热带雨林幼树叶片中的生态化学计量比对海拔变化的响应显著[11],南亚热带中幼林针阔混交林生态化学计量特征与其土壤存在一定关系[28],温度对贡嘎山峨眉冷杉幼苗化学计量特征的影响差异显著[29]。这些研究集中体现了植物的生态化学计量特征与其所处海拔、土壤和温度等因素存在一定关系,闽北闽粤栲天然林4种优势树种幼树不同器官C、N和P化学计量特征的差异是否与这些因素有关还有待进一步研究。 (1)闽北闽粤栲天然林中4种优势树种幼树不同器官间各元素含量差异显著,其中,C含量最大,表现为叶>枝>干>根,而N、P含量表现为叶>枝>根>干。 (2)闽北闽粤栲天然林4种优势树种幼树不同器官中C∶P远高于C∶N与N∶P,其次为C∶N,N∶P最小,其化学计量特征在不同器官中差异极显著,幼树对养分的利用效率较低。 (3)闽北闽粤栲天然林4种优势树种幼树不同器官中的P含量均较低,且N∶P均显著高于16,其幼树生长更新主要受N、P元素含量的影响,并可能受P元素缺失的制约。 [1] 贺金生,韩兴国. 生态化学计量学:探索从个体到生态系统的统一化理论[J]. 植物生态学报,2010,34(1):2-6. [2] Schimel D S. All life is chemical[J]. BioScience,2003,53(5):521-524. [3] Michaels A F. The ratios of life. Science[J]. Science,2003, 300(5621): 906-907. [4] Elser J J, Fagan W F, Denno R F,etal. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812): 578-580. [5] McGroddy M E, Daufresne T, Hedin L O. Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios[J].Ecology, 2004, 85(9): 2390-2401. [6] Schade J D, Espeleta J F, Klausmeier C A,etal.A conceptual framework for ecosystem stoichiometry:balancing resource supply and demand[J]. Oikos, 2005, 109(1): 40-51. [7] Hall S R, Leibold M A, Lytle D A,etal. Inedible producers in food webs: Controls on stoichiometric food quality and composition of grazers[J]. American Naturalist, 2006, 167(5): 628-637. [8] 施家月,王希华,闫恩荣,等. 浙江天童常见植物幼树器官的氮磷养分特征[J]. 华东师范大学学报:自然科学版,2006,2006(2):121-129. [9] 常云妮,钟全林,程栋梁,等. 尤溪天然米槠林植物碳氮磷的化学计量特征及其分配格局[J]. 植物资源与环境学报,2013,22(3):1-10. [10] 陈亚南,马露莎,张向茹,等. 陕西黄土高原刺槐枯落叶生态化学计量学特征[J]. 生态学报,2014,34(15):4412-4422 [11] 栗忠飞,刘文胜,张 彬,等. 西双版纳热带雨林幼树C、N、P的生态化学计量比对海拔变化的响应[J]. 中南林业科技大学学报,2012,32(5):80-85. [12] 石贤萌,杞金华,宋 亮,等. 哀牢山中山湿性常绿阔叶林两种优势幼苗C、N、P化学计量特征及其对N沉降增加的响应[J]. 植物生态学报,2015,39(10):962-970. [13] 苏凌燕,郑德祥,钟兆全,等. 闽北闽粤栲天然林林隙特征及干扰状况[J]. 森林与环境学报,2015,35(2):125-130. [14] 蓝文升,钟兆全,郑德祥,等. 闽北天然闽粤栲林分空间结构研究[J]. 北华大学学报:自然科学版,2014,15(3):398-403. [15] 管大跃,黄国泉. 闽粤栲天然林生物量及预测模型研究[J]. 福建林业科技,2000,27(2):34-36. [16] 李俊清. 森林生态学[M]. 北京:高等教育出版社,2010:224-226. [17] 栗忠飞,郭盘江,刘文胜,等. 哀牢山常绿阔叶林幼树C、N、P生态化学计量特征[J]. 东北林业大学学报,2013,41(4):22-26. [18] 董 鸣. 陆地生物群落调查观测与分析[M]. 北京:中国标准出版社, 1997:154-155. [19] Krieger H, Schaefer H, Peng L. Growth dynamics of a plantedEucalyptusexseria(F. Muell) stand in south China: adaptation of generic a simulation model[M]. Kassel: Kassel University Press, 1990:10-15. [20] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. [21] Han W, Fang J, Guo D,etal. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2):377-385. [22] 任书杰,于贵瑞,陶 波,等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学,2007,28(12):2665-2673. [23] Elser J J, Sterner R W, Gorokhova E,etal. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6):540-550. [24] 邬 畏,何兴东,周启星. 生态系统氮磷比化学计量特征研究进展[J]. 中国沙漠,2010,30(2):296-302. [25] Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J]. Journal of Applied Ecology, 2003, 40(3):523-534. [26] 阎恩荣,王希华,周 武. 天童常绿阔叶林演替系列植物群落的N∶P化学计量特征[J]. 植物生态学报,2008,32(1):13-22. [27] 王恩熙,谢锦升,杨柳明,等. 中亚热带杉木人工老龄林林下植被叶片碳氮磷化学计量特征[J]. 亚热带资源与环境学报,2015,10(4):31-37. [28] 周 丽,张卫强,唐洪辉,等. 南亚热带中幼龄针阔混交林生态化学计量特征[J]. 生态环境学报,2014,23(11):1732-1738. [29] 羊留冬,杨 燕,王根绪,等. 短期增温对贡嘎山峨眉冷杉幼苗生长及其CNP化学计量学特征的影响[J]. 生态学报,2011,31(13):3668-3676. (责任编辑:徐玉秀) DOI:10.13275/j.cnki.lykxyj.2017.01.022 柿炭疽病防治药剂的筛选 邓全恩1,丁向阳2*,徐建强3,李建安1,谷战英1 (1.中南林业科技大学,经济林育种与栽培国家林业局重点实验室,经济林培育与保护教育部重点实验室,湖南 长沙 410004; 2.河南省林业科学研究院,河南 郑州 450008; 3.河南科技大学,河南 洛阳 471003) 收稿日期: 2016-05-10 关键词:柿;炭疽病;药剂;筛选 Abstract: [Objective]This study was designed to screen the fungicides for controlling pathogens of persimmon anthracnose by laboratory and field test. [Method]4 representative persimmon anthracnose strains collected from Qingzhou of Shandong Province, Gongcheng of Guangxi Zhuang Autonomous Region, Luanchuan of Henan Province and Fuping of Shanxi Province, were selected as the trial materials. By comparing the minimum inhibitory concentration (MIC) value and the concentration for 50% of maximal effect (EC50) value of 22 kinds of fungicides belonging to 10 categories to decide the inhibition activity of each fungicide. 5 kinds of fungicide were finally selected to conduct field trials. [Result]The results show that: benzimidazole, demethylation inhibitor and benzene-pyrrole acted better, the MIC values were below 10 μg·mL-1, and the EC50within the scope of 0.011 2-0.520 3 μg·mL-1. Field trials demonstrated that the inhibition efficacy of 30% difenoconazole WG (1500×), 240 g·L-1thifluzamide SC (2000×), 240 g·L-1pyraclostrobine SC (1000×), 25% prochloraz EC (2000×), 25 g·L-1fludioxonil SC (1000×) were between 18.72%-47.76% when investigated 10 days after the fourth spraying. [Conclusion]The speed of onset and final control effect of fludioxonil and prochloraz were better than the other three kinds of fungicides. Keywords: persimmon; anthracnose; fungicide; screening 柿(DiospyroskakiThunb.)为柿科(Ebenaceae)柿属(DiospyrosL.)落叶乔木或灌木,生态幅广、适应性极强[1],是我国主要的木本粮食树种,近年来随着国家对木本粮油产业的重视[2],柿重新受到了人们的关注,炭疽病是柿树上发生的主要侵染性病害,严重制约了柿产业的发展[3]。 该病主要发生在柿树叶片、枝条、果蒂和果实上,不同品种及同一品种的不同部位发病情况不同[4]。叶片发病时叶柄、叶脉变成黑色,形成坏死病斑,这也是柿炭疽病区别于其它柿叶部病害的明显标志;枝条发病时,主要表现在柿树的幼梢,严重时病斑互相融合,从而引起叶片脱落,甚至导致枝条死亡[5];柿蒂发病时,表现为黑色坏死现象;果实发病时变红变软,提前脱落。柿炭疽病的病菌主要以菌丝体在幼龄枝条病组织及芽眼中越冬,也可在病果、叶痕和冬芽中越冬,翌年条件适宜时以孢子的形式借风雨和昆虫传播,主要侵染新梢及幼果。一般新梢6月上旬开始发病,雨季发病较为严重,果实6月下旬至7月中下旬发病。Weir等基于分子生物学和形态学的差异将柿树炭疽病菌的表位型界定为柿盘长孢菌GloeosporiumkakiHori,而分类学上将其归类为柿树炭疽菌Colletotrichumhoriinom. nov[6]。 炭疽病在多种果树、农作物上均有发生和研究报道[7-11],柿炭疽病的研究相对较少,主要集中在部分品种炭疽病菌的分离鉴定和分子序列分析[12-14]、侵染过程、发生发展规律及综合防治技术[5,15],对于柿炭疽病防治药剂的筛选还处于初步研究阶段[16-17],研究涉及的药剂种类较少或者为早期药剂,加之炭疽病在多种植物上发生,所以生产中使用的防治药剂种类繁杂、效果不一、安全性不确定,因此对柿炭疽病防治药剂的筛选显得格外重要。本文针对柿炭疽病菌选择了10类22种杀菌剂,室内测定各种药剂对柿炭疽病菌的抑制作用,筛选出抑制力强的药剂进行田间防治试验。 1.1 常见杀菌剂对柿炭疽病病菌室内毒力测定 选取采集于山东青州、广西恭城、河南栾川及陕西富平的4个具有代表性的菌株;杀菌剂包括苯并咪唑类、三唑类、二甲酰亚胺类等10类22种。(1)MIC值测定:将供试菌株接种在含1、5、10 μg·mL-1杀菌剂的PSA平板上,不含药的PSA平板为对照,每皿2个菌株,每处理2次重复,25℃培养4 d后,采用十字交叉法测量各处理的菌落直径,求平均值后,计算抑制率,得出药剂对炭疽病菌的最低抑制浓度(Minimum inhibitory concentration,MIC)。抑制率=(对照菌落直径-处理菌落直径/(对照菌落直径-5)×100%;(2)EC50值测定:采用菌丝生长速率法[18]进行测定,采用MIC测定中的抑制率公式求出各个药剂浓度对菌丝生长抑制率,所得数据经Finney概率分析后用DPS v12.01统计软件求出毒力回归方程式及EC50。实验步骤包括: 1)配置杀菌剂母液。将备试原药用丙酮溶解, 配制成10 g·L-1的母液备用。 2)炭疽菌株进行活化。把炭疽的菌丝接种于培养皿的PSA培养基平面中央,培养4 d左右待用。3)制备带毒培养基。依据前期各杀菌剂对柿炭疽病菌MIC值的测定结果,设置系列浓度梯度(表1),根据浓度梯度加入一定量的杀菌剂母液,制备系列浓度的带毒培养基。 4)炭疽病菌接种。将活化后的炭疽病菌用打孔器在菌落边缘打出直径为0.5 cm的菌饼,然后接种于含药剂系列浓度的PSA培养基上。 5)病菌抑制效果观测。炭疽病菌于25℃培养箱中培养6 d左右,观察菌丝生长状况,测量菌落直径,算出抑制率。以不加药剂的PSA平板为空白对照,每个药剂浓度3个重复。 1.2 药剂田间防治试验 1.2.1 试验地情况 试验在广西平乐县(110°38'35.91″ E,24°38'0.32″ N)沙子镇一个10年生水柿园进行,株行距为3.5 m×4 m。柿园地形为平地,红壤,pH值6.2,肥力中等,管理水平中等,整个园区发病情况基本一致。 1.2.2 试验方法 结合已经得到的室内防治药剂筛选结果,选择其中效果较好和未做室内试验但较新型药剂共5种进行柿炭疽病室外防治药剂的筛选试验,设5个药剂处理(表2)和1个清水对照,每处理3次重复,共18个小区,每小区3株柿树,每个处理9棵树喷1喷雾器药液(22 kg)。施药时间为2015年8月10日、9月1日、9月20日和10月8日,皆为晴天或多云,喷药后3 d内均未下雨。前两次施药时间为下午4:00左右,后两次施药时间为下午3:00左右。 1.2.3 调查方法 在每株调查柿树的树冠中等偏上部位东、西、南、北4个方向定枝4个,每枝果数不少于10个(广西水柿发生该病主要表现在果实上,所以本试验不调查叶片及果蒂),于每次喷药前调查病果率(由于柿果发生炭疽病导致的果实过早脱落 表1 杀菌剂来源及处理浓度梯度 Table 1 Sources of fungicides and gradient concentration of the treatment 杀菌剂Fungicides生产厂商Manufacturer浓度梯度Concentrationgradient/(μg·mL-1)苯醚甲环唑Difenoconazole山东潍坊双星农药有限公司ShandongWeifangShuangxingPesticideCo.,Ltd.0、0.1562、0.3125、0.6250、1.2500、2.5000、5.0000戊唑醇Tebuconazole江苏克胜股份有限公司JiangsuKeshengCo.,Ltd.0、0.1562、0.3125、0.6250、1.2500、2.5000、5.0000噻菌灵Thiabendazole浙江绿州生物技术有限公司ZhejiangLvzhouBiotechnologyCo.,Ltd.0、0.0156、0.0312、0.0625、0.1250、0.2500、0.5000多菌灵Carbendazim山东禾宜生物科技有限公司ShandongHeyiBiotechnologyCo.,Ltd.0、0.0156、0.0312、0.0625、0.1250、0.2500、0.5000甲基托布津Methylthiophanate南京柏蓝科技有限公司NanjingBailanTechnologyCo.,Ltd.0、0.0312、0.0625、0.1250、0.2500、0.5000、1.0000苯菌灵Benomyl南京柏蓝科技有限公司NanjingBailanTechnologyCo.,Ltd.0、0.0312、0.0625、0.1250、0.2500、0.5000、1.0000速克灵Procymidone日本住友化学株式会社JapanSumitomoChemicalCorp0、0.6250、1.2500、2.5000、5.0000、10.0000、20.0000扑海因Rovral拜耳作物科学(中国)有限公司BayerCropScience(China)Co.,Ltd.0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000嘧菌酯Azoxystrobin中国农科院植保所农药厂ThePesticideFactoryofIPPofCAAS0、0.4688、0.9375、1.8750、3.7500、7.5000、15.0000噻呋酰胺Thifluzamide利民化工股份有限公司LiminChemicalCo.Ltd.0、0.6250、1.2500、2.5000、5.0000、10.0000、20.0000氟酰胺Amide拜耳作物科学(中国)有限公司BayerCropScience(China)Co.,Ltd.0、0.6250、1.2500、2.5000、5.0000、10.0000、20.0000乙霉威Diethofencarb海利尔药业集团股份有限公司HailierPharmaceuticalGroupCo.,Ltd.0、2.5000、5.0000、10.0000、15.0000、20.0000、40.0000嘧霉胺Pyrimethanil中国农科院植保所农药厂ThePesticideFactoryofIPPofCAAS0、10.0000、20.0000、40.0000、50.0000、60.0000、80.0000吡唑醚菌酯Pyraclostrobin德国巴斯夫公司GermanBasfInc0、0.4688、0.9375、1.8750、3.7500、7.5000、15.0000丙环唑Propiconazole陕西标正作物科学有限公司ShaanxiBiaozhengCropScienceCo.,Ltd.0、0.0781、0.1563、0.3125、0.6250、1.2500、2.5000烯唑醇Diniconazole江苏剑牌农化股份有限公司JiangsuSwordAgriculturalCo.,Ltd.0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000三唑酮Ketotriazole江苏剑牌农化股份有限公司JiangsuSwordAgriculturalCo.,Ltd.0、0.6250、1.2500、2.5000、5.0000、10.0000、20.0000甲基立枯磷Tolclofos-methyl青岛瀚正益农生物科技有限公司QingdaoHanzhengyinongAgroSciencesBiotechnologyCo.Ltd.0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000咯菌腈Fludioxonil瑞士先正达作物保护有限公司SyngentaCropProtectionCo.Ltd.0.0049、0.0098、0.0195、0.0391、0.0781、0.1563百菌清Chlorothalonil利民化工股份有限公司LiminChemicalCo.Ltd.0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000福美双Thiram河北赞峰生物工程有限公司HebeiZanfengBiologicalEngineeringCo.,Ltd.0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000代森锰锌Mancozeb美国杜邦公司DuPontEngineeringPolymers0、0.3125、0.6250、1.2500、2.5000、5.0000、10.0000 注:以上杀菌剂均按有效成分计算质量浓度,按照每种农药的说明用量范围而定。 Note: The concentration of above fungicides are calculated according to the effective composition, in accordance with the scope of the description of each pesticide dosage. 表2 田间防治试验药剂Table 2 Information about the fungicides of field treatment 和病斑的大小无关,所以本试验调查时只调查病果率,不调查发病等级),考虑到每次病果率调查时由于发生炭疽病导致的果实脱落情况,病果率的计算方法为:病果率=[调查当次树上病果数+(初次调 查果实总数-当次调查果实总数)-其它因素落果数]/初次调查果实总数。最后一次调查时间为喷药后的第10天即10月18日。防治效果=(对照病果率-处理病果率)/对照病果率×100%。 1.3 数据处理 应用Microsoft Excel 2007、SPSS19.0和DPS v12.01统计软件对数据进行处理和制表、制图等。 2.1 室内不同药剂对柿炭疽病菌的抑制作用 供试的10类22种杀菌剂对采集于山东青州、广西恭城、河南栾川及陕西富平的4个菌株的抑制作用见表3,对柿炭疽病菌菌丝生长抑制效果较好的杀菌剂有苯并咪唑类、脱甲基抑制剂类(三唑类、DMIs)、苯吡咯类3类杀菌剂,它们有的品种已应用在柿炭疽病的化学防治上,有的则正在作为复配剂进行开发。琥珀酸脱氢酶抑制剂类(SDHIs)及甲氧基丙烯酸酯类杀菌剂(QoIs)对柿炭疽病菌毒力差异较大,需作进一步的探索。其中琥珀酸脱氢酶抑制剂类(SDHIs)中的噻呋酰胺对于广西水柿炭疽病病菌抑制作用较强,所以选用该药进行室外试验。三唑类药物虽然对于柿炭疽病病菌抑制作用较强,但其代表性药剂多菌灵、苯菌灵等能诱发病原菌的抗性,同类品种间有正交互抗性,应注意与其它类型杀菌剂混用或轮换使用,所以室外试验未选择三唑类药剂。结合表2与市场上的新型杀菌剂使用情况,室外试验选择了苯醚甲环唑、咪鲜胺、咯菌腈、吡唑醚菌酯、噻呋酰胺5种杀菌剂。 表3 不同杀菌剂对柿炭疽病菌的抑制作用Table 3 Inhibition effects of different fungicides on pathogens of anthracnose of persimmon 2.2 不同药剂田间防治效果 表4 不同杀菌剂对柿炭疽病的田间防治效果Table 4 Inhibition effects of different fungicides on anthracnose of persimmon in field treatment 注:不同小写字母表示P≤0.05水平下显著性差异。 Note: The different small letters represent significant difference of P level less than or equal to 0.05. 本试验选择了10类22种杀菌剂,基本上覆盖了近年来市场上病害防治的常用药剂,并采用室内试验和田间试验相结合的方法进行药剂筛选,为制定较为有效的柿炭疽病防治方法打下了基础。室内试验中发现三唑类对炭疽病菌抑制效果较好,但由于三唑类杀菌剂的作用机理是抑制病原菌体内甾醇的脱甲基化过程,作用位点相对单一,因此存在较高的抗药性风险[19],所以田间试验未选择该类药剂,这符合试验方案中寻找有效、可持续药剂的目标和理念。田间试验结果显示虽然进行了药剂处理,但最终发病率都超过了30%,这可能因为试验柿园往年发病较重且未经过正规的防治,所以出现了前期防效较好,但随着时间推移炭疽病的发病率呈现上升的趋势。后续工作包括:(1)补测咪鲜胺和吡唑醚菌酯对柿炭疽病病菌的室内毒力数据,并与其它药剂的室内毒力数据和大田试验结果进行比对分析;(2)从药剂使用时间对最终防治效果的影响方面开展研究,如发病前防治和发病后防治的效果差异来比较不同药剂的防治效果和预防效果,进而探讨不同药剂的最佳喷药时期[20];(3)通过不同药剂的配合使用筛选出效果较好的药剂组合,为复配新型药剂做好准备工作。由于时间限制本次室外防治试验结果是1年得出的,后续研究需关注连续多年使用的效果差异以比较不同药剂的长期效果;(4)对各种杀菌剂进行综合比较,诸如对植物生长的影响、对环境的影响、成本分析等,力求经济合理地施用高效低毒杀菌剂。 参考文献: [1] 邓全恩,龚榜初,吴开云,等. 柿果生理性病害顶腐病发病规律调查研究[J]. 林业科学研究,2014,27(5):689-696. [2] 谭晓风,马履一,李芳东,等. 我国木本粮油产业发展战略研究[J]. 经济林研究,2012,30(1):1-5. [3] 刘开启,牟惠芳,刘凤英. 柿炭疽病的研究[J]. 山东农业大学学报,1988, (4): 69-71. [4] Geffroy V,Sévignac M,De Oliveira J C,etal. Inheritance of partial resistance againstColletotrichumlindemuthianuminPhaseolusvulgarisand co-localization of quantitative trait loci with genes involved in specific resistance[J].Mol Plant Microbe Interact,2000,13(3):287-296. [5] 张敬泽,徐 同. 柿树炭疽病菌在越冬枝条上的菌态及数量[J]. 植物保护学报,2003,30(4):438-439. [6] Weir B S,Johnston P R. Characterisation and neotypification ofGloeosporiumkakiHori asColletotrichumhoriinom. nov[J].Mycotaxon,2010,111(1): 209-219. [7] 杨光道,段 琳,束庆龙,等. 油茶果皮花青素、糖含量和PAL 活性与炭疽病的关系[J]. 林业科学,2007,43(6):100-104. [8] 雷新涛, 赵艳龙, 姚全胜,等. 杧果抗炭疽病种质资源的鉴定与分析[J]. 果树学报,2006, 23(6): 838-842. [9] 吴良庆,朱立武,衡 伟,等. 砀山梨炭疽病病原鉴定及其抑菌药剂筛选[J]. 中国农业科学,2010,43(18):3750-3758. [10] 王惠哲,李淑菊,刘秀峰,等. 与黄瓜抗炭疽病相关基因连锁的AFLP标记的筛选[J]. 园艺学报,2007, 34 (1): 213-216. [11] 孙春英,毛胜利,张正海,等. 辣椒抗炭疽病遗传与育种研究进展[J]. 园艺学报2013,40(3):579-590. [12] 余贤美,侯长明,王 洁,等. 山东牛心柿炭疽病菌的分离鉴定及致病性[J]. 林业科学,2015,51(4):126-133. [13] 余贤美,侯长明,王 洁,等. 次郞甜柿炭疽病菌的分离鉴定及其rDNA-ITS序列分析[J]. 经济林研究, 2014,32(1):45-50. [14] 张敬泽,徐 同,何黎平. 浙江无核柿炭疽病菌鉴定及附着胞形成过程中的核相变化[J]. 菌物学报,2005,24(3):446-456. [15] 贾克锋,陈 雁,王利忠. 日本甜柿炭疽病发生规律及防治技术[J]. 浙江林学院学报,1997, 14 (1): 45-49. [16] 丁向阳, 徐建强, 邓全恩, 等. 柿炭疽病化学防治药剂的初步筛选[J]. 经济林研究,2015,33(4):148-151, 156. [17] 曲健禄, 武海斌,范 昆,等. 柿树炭疽病菌的生物学特性及几种杀菌剂对其的抑制作用[J]. 农药学学报,2012,14(5): 503-509. [18] Wang Y H,Chen J, Zhu Y C,etal. Susceptibility to neonicotinoids and risk of resistance development in the brown plant hopper,Nilaparvatalugens(Stal) (Homoptera: Delphacidae)[J].Pest Manag Sci,2008,64(12):1278-1284. [19] 潘金菊,慕 卫,翟如环,等. 9 种杀菌剂对苹果斑点落叶病菌和轮纹病菌的毒力比较[J]. 农药科学与管理,2006,27 (6):16-18. [20] 刘永刚,李佳佳,李昭煜,等. 枸杞炭疽病防治药剂筛选[J]. 植物保护,2016,42(1):230-234 (责任编辑:金立新) DOI:10.13275/j.cnki.lykxyj.2017.01.023 山核桃不同无性系果实性状及营养成分分析 常 君1,任华东1*,姚小华1,王开良1,周 燕2,周振琪2 (1.中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,浙江 杭州 311400; 2.浙江省建德市林业局,浙江 建德 311600) 收稿日期: 2016-03-14 关键词:山核桃;无性系;果实性状;营养成分 Abstract: [Objective]To study the nut fruit shape, the composition, the nutrient contents and their variation ofCaryacathayensisclone. [Method]11 trait and nutrient indices ofC.cathayensisnut fruit from trial forest were analyzed by the methods of variance analysis and multiple comparison based on the national standard of China. [Result]The results showed that there was a significant difference in nut length between the fresh fruits and the nuts, and no significant difference was found in the rest indices ofC.cathayensis. The kernels ofC.cathayensiswere rich in fats (52.64%-58.51%), proteins (55.07-65.50 mg·g-1), soluble sugars (0.26%-0.68%) and mineral elements such as K, Ca, Na, Mg, Zn, Mn, and Fe. 17 kinds of amino acid were identified in the kernels ofC.cathayensis. There were extremely significant differences in the contents of amino acids among the clones ofC.cathayensis. [Conclusion]According to the characteristics of the nut nutrients ofC.cathayensis, the directions of breeding and utilizing ofC.cathayensisare suggested. Three clones are comprehensively selected as the clones of high yield and high quality; one clone is selected as the clone suitable for edible oil production; and two clones are selected as the clones with high contents of amino acids. Keywords:Caryacathayensis; clones; fruit characteristics; nutrients 山核桃(CaryacathayensisSarg.),又称山核、山蟹、小核桃,属胡桃科(Juglandaceae)、山核桃属(CaryaNutt.)[1-3]物种,其坚果种仁富含脂肪、蛋白质及大量人体必需矿质元素,风味独特,深受消费者喜爱,是我国南方重要的特色干果和木本油料树种,也是我国特有的坚果树种。我国浙皖天目山区是山核桃天然分布区,也是山核桃的主产区,主要包括浙江杭州地区的临安市、淳安县、桐庐县、富阳市、建德市和湖州市的安吉县,安徽黄山市的歙县和宣城市旌德县、绩溪市、宁国市等县(市)[4-6]。种植山核桃经济效益好,年亩产值可达万元以上,是产区群众主要的收入来源,随着栽培面积不断扩大,山核桃产业已成为主产区重要支柱产业。 长期以来我国山核桃生产以实生栽培为主,生产林分个体分化严重,存在产量、品质不稳定等问题。实现种植品种无性化是山核桃产业持续高效发展的必然选择,也是我国山核桃产业亟待解决的科学问题。有关山核桃无性系品种选育的研究国内仅有无性扩繁方面的研究报道[7-12],未见成功的无性系选育研究报道,对山核桃果实性状及营养成分分析与评价也多是基于实生优株或实生资源果实生长发育规律、蛋白质及氨基酸含量与变异、土壤微生物量碳氮的变化、山核桃油提取工艺、脂肪酸组成及加工炒制过程中营养成分、脂肪酸氧化及抗氧化能力变化中的部分指标进行分析[13-22]。作者单位在攻克无性繁殖技术后成功开展了山核桃无性系测定试验,在浙江省金华市东方红林场建立了山核桃无性系测定试验林。本文以该试验林无性系为研究对象,对11个参试无性系的果实性状及营养成分组成和含量进行了测定与分析,旨在探明山核桃无性系果实形态结构特征、内含物营养成分组成与含量及其变异规律,并依据各参试无性系的果实性状和种仁营养成分特点,探讨山核桃无性系选育方向并筛选适宜不同利用方向的优良无性系。 1.1 试验材料 试验材料取自浙江省金华市东方红林场8年生山核桃无性系试验林,试验地为丘陵缓坡地,普通红壤,立地条件较一致,土地肥力一般。试验采用随机区组设计,5株小区3次重复,参试无性系共11个,分别为大源2号、大源4号、大源5号、高岭1号、高岭3号、高岭5号、高岭7号、高岭8号、徐坑24号、徐坑40号和徐坑89号。 1.2 测定方法 采用山核桃鲜仁为测定材料,按照以下方法进行相关指标的测定:粗脂肪含量按照GB/T14772-2008《食品中粗脂肪的测定》方法测定,蛋白质含量按照GB 5009.5-2010《食品中蛋白质的测定》方法测定,可溶性糖含量按照NY/T 1278-2007《蔬菜及其制品中可溶性糖的测定 铜还原碘量法》方法测定,单宁含量按照NY/T 1600-2008《水果、蔬菜及其制品中单宁含量的测定 分光光度法》方法测定,铬含量按照GB 5009.123-2014《食品安全国家标准 食品中铬的测定》方法测定,钙含量按照GB/T 5009.92-2003《食品中钙的测定》方法测定,铜含量按照GB/T 5009.13-2003《食品中铜的测定》方法测定,锌含量按照GB/T 5009.14-2003《食品中锌的测定》方法测定,钾、钠含量按照GB/T 5009.91-2003《食品中钾、钠的测定》方法测定,镁、锰、铁含量按照GB/T 5009.90-2003《食品中铁、镁、锰的测定》方法测定,参照GB/T 5009.124-2003《食品中氨基酸的测定》方法测定氨基酸含量。 1.3 数据处理 所有数据采用Excel 2007、DPS 14.5数据处理软件进行统计分析。 2.1 山核桃无性系果实形态特征性状变异分析 表1为11个山核桃参试无性系果实形态特征性状的变异分析结果。从分析结果可以看出,山核桃果实在青果单果质量、青果果长、青果果宽、坚果单果质量、出籽率、坚果果长、坚果果宽、坚果壳厚、果仁质量及出仁率10个测定性状上无性系间均存在一定程度的变异,但多数性状变异幅度不大,变异幅度最大的性状是坚果壳厚(变异系数为7.15%),坚果果宽变异幅度最小(变异系数为0.83%)。方差分析结果表明,除青果果长与坚果果长无性系间表现出显著差异外,其它8个指标无性系间差异均未达到显著水平,表明山核桃果实形态无性系间总体上比较一 致,但山核桃青(坚)果果长性状在参试无性系间确实存在本质上的差异,差异具有一定的遗传稳定性,说明在无性系水平上进行山核桃果长特征的选择改良是可能的,有可能取得良好的效果。进一步对参试无性系的SSR多重比较表明:高岭7号、高岭8号及高岭3号3个无性系的青果果长显著高于其它无性系,其中高岭7号无性系青果果长最长(为35.95 mm),高出最短青果果长大源4号无性系(为32.84 mm)9.47%。坚果果长SSR多重比较结果表明:高岭8号、高岭7号2个无性系坚果果长显著高于其它无性系,其中高岭8号无性系坚果果长最长(为26.67 mm),较最短坚果果长大源4号无性系(为24.98 mm)高出6.77%。 表1 山核桃无性系果实形态特征变异分析结果Table 1 Variation analysis of characteristics of C. cathayensis clones 注:显著水平为5%,字母相同为差异不显著,字母不同为差异显著,*为差异显著。 Note: At the significant level of 5%, no significant difference for the same letter, and different letters for the significant difference, *mean the differences was significant. 2.2 不同山核桃无性系种仁内含物含量比较分析 粗脂肪、蛋白质、可溶性糖和单宁是影响山核桃种仁品质的重要质量指标。对11个山核桃参试无性系种仁粗脂肪、蛋白质、可溶性糖和单宁含量的测定分析结果表明(表2):种仁粗脂肪、蛋白质、可溶性糖和单宁含量在参试无性系间均存在一定的变异,变异程度差异较大,种仁可溶性糖含量变异程度最大,无性系间变异系数高达39.67%,粗脂肪含量无性系间变异较小,变异系数仅为2.76%。进一步的方差分析结果表明,可溶性糖含量参试无性系间的差异达极显著差异水平,说明山核桃无性系在种仁可溶性糖含量上存在显著的遗传变异。经SSR多重比较结果表明:高岭7号、徐坑40号与高岭8号3个无性系种仁可溶性糖含量极显著高于其它无性系,种仁可溶性糖含量最高的无性系是高岭7号,含量达0.68%,高出参试无性系总体均值74.00%,是含量最低无性系高岭1号(含量0.29%)的2.34倍。种仁中的单宁是导致种仁涩口的主要物质,其含量高低直接影响山核桃仁的风味,种仁单宁含量在参试无性系中也表现出较大的差异,无性系间的变异系数达13.7%,其含量最高的是徐坑40号(单宁含量12.43 mg·g-1),其含量高出所有参试无性系总体均值(9.87 mg·g-1)25.9%,是含量最低无性系大源4号(单宁含量8.11 mg·g-1)的1.53倍。 表2 山核桃不同无性系营养成分指标变异分析Table 2 Variation analysis of nutrients of C. cathayensis clones 注:显著水平为1%,字母相同为差异不显著,字母不同为差异显著,**为差异极显著。 Note: At the significant level of 1%, no significant difference for the same letter, and different letters for the significant difference, ** mean the difference was extremely significant. 2.3 山核桃不同无性系矿质元素含量比较分析 植物籽粒微量元素质量分数常受土壤母质性质的巨大影响,并可能由此影响农产品品质[23],矿质元素含量的高低是评鉴山核桃种仁品质的主要指标。山核桃种仁中富含钾、钙、钠、镁、锌等人体必需矿质营养物质,与郑春霞等[24]测定的新疆9种干果微量元素比较可以发现,山核桃种仁中的锰、铜、锌含量明显大于新疆地区9种干果的含量,这也与钱新标等[23]研究结果一致。表3为11个山核桃参试无性系种仁中9种矿质元素含量变异分析结果,从中可以看出:山核桃种仁矿质元素总量及各元素含量在无性系间均存在一定程度的差异,种仁中矿质元素总量参试无性系总体均值为4 530.49 ug·g-1,变异系数6.51%,总量最高的是徐坑24号(5 073.78 ug·g-1),最低的是高岭7号(3 906.71 ug·g-1),两者相差1 167.07 ug·g-1;山核桃种仁中含量最高的矿质元素是钾(K)(平均含量为3 087.66 ug·g-1),其次为镁(Mg)(平均含量为818.98 ug·g-1)。9种矿质元素中,无性系间变异最大的是钠(Na)和铬(Cr),变异系数分别高达46.15%和24.00%,说明铬(Cr)和钠(Na)在参试无性系间存在较大的差异。对参试无性系种仁矿质元素总量及各元素含量的方差分析结果表明,山核桃种仁中钠(Na)含量参试无性系间差异达极显著水平,钙(Ca)含量参试无性系间差异达显著水平。经SSR多重比较,大源4号无性系钠(Na)含量高达11.53 ug·g-1,极显著高于其它无性系,是所有参试无性系总体均值(5.36 ug·g-1)的2.15倍;徐坑24号和徐坑89号钙(Ca)含量显著高于其它无性系,其中徐坑24号无性系钙(Ca)含量最高,其含量(507.93 ug·g-1)高出所有参试无性系总体均值(424.89 ug·g-1)19.54%,是含量最低无性系高岭7号(322.53 ug·g-1)的1.57倍。 表3 山核桃不同无性系的矿质元素含量方差分析、多重比较及变异分析Table 3 Variance analysis, multiple comparison and variation analysis of mineral element contents ofC. cathayensis clones ug·g-1 注:显著水平为1%,字母相同为差异不显著,字母不同为差异显著,*为差异显著,**为差异极显著。 Note: At the significant level of 1%, no significant difference for the same letter, and different letters for the significant difference, *mean the differences was significant, ** mean the difference was extremely significant. 2.4 山核桃不同无性系种仁蛋白氨基酸组分含量比较分析 氨基酸是蛋白质的组成成分,在人体营养和生理上占有重要地位,为满足人体新陈代谢的需要,人们必须从食物中摄取人体所需的氨基酸[25]。山核桃种仁中富含17种氨基酸,包含人体所必需8种氨基酸中的7种。表4为11个山核桃参试无性系种仁氨基酸含量的分析测定结果,从中可以看出:参试无性系种仁中氨基酸总量总体均值为53.99 mg·g-1,7种人体必需氨基酸总量总体均值为18.39 mg·g-1,占氨基酸总量的34.06%。山核桃种仁所含的17种氨基酸中,大源2号谷氨酸和精氨酸含量最高,分别为12.02 mg·g-1和7.51 mg·g-1。在山核桃种仁所含人体所必需的7种氨基酸中,大源2号亮氨酸含量最高,含量为4.59 mg·g-1,占7种人体必需氨基酸总量的24.96%。 对11个山核桃参试无性系果实形态10个指标的测定分析结果表明:参试无性系平均青果单果质量为17.99 g,平均坚果单果质量为5.57 g,较解红恩等[26]对山核桃实生起源林分测定的平均鲜果单果质量13.58 g和贾晓东等[27]平均坚果单果质量3.81 g高出32.47%和46.19%,平均坚果果长和坚果果宽分别为26.05 mm和21.46 mm,较贾晓东等[27]平均坚果果长21.85 mm和坚果果宽19.82 mm分别高出19.22%和8.27%,主要原因可能是山核桃无性系化种植果实大小较实生种植更为均一,品质更好。在10个果实形态测定指标中,仅青果果长和坚果果长在无性系间存在显著水平的差异,在11个山核桃参试无性系中,高岭8号、高岭7号2个无性系坚果果长显著高于其它无性系。 对11个山核桃无性系果实种仁蛋白内含氨基酸组分及含量测定分析结果表明:山核桃种仁含有17种氨基酸,包含人体所必需8种氨基酸中的7种,这与王冀平等[13]、刘力等[15]、李新委等[17]和章亭洲等[29]研究结果一致。参试无性系种仁氨基酸总量均值为53.99 mg·g-1,必需氨基酸总量均值为18.39 mg·g-1,占总氨基酸含量的34.06%,与WHO/FAO规定的标准(40%)[30]差距较小,在所检测出的氨基酸中,谷氨酸和精氨酸含量显著高于其它氨基酸含量,精氨酸含量次之。谷氨酸和天冬氨酸是鲜味氨基酸,其中谷氨酸还参与脑组织代谢,具有健脑作用,能促进脑细胞进行呼吸,有利于脑组织中的氨排除,天冬氨酸还具有降血压、抗疲劳等多种生理功能[29,33],精氨酸可促进生长发育、创伤愈合和增强细胞免疫功能等效用[13,34],因此山核桃可作为优质的蛋白质资源。对11个山核桃无性系种仁氨基酸含量的方差分析结果表明,氨基酸总量、人体必需氨基酸总量及各氨基酸含量在参试无性系间均表现出极显著差异。在参试无性系中,大源2号、大源4号和高岭7号3个无性系的氨基酸总量和人体必需氨基酸总量极显著高于其它无性系;大源2号、大源4号和高岭7号3个无性系的天冬氨酸、丝氨酸、谷氨酸、甘氨酸、丙氨酸、缬氨酸、异亮氨酸、亮氨酸、赖氨酸和精氨酸10种氨基酸含量极显著高于其它无性系;大源2号无性系的苏氨酸、胱氨酸和组氨酸含量极显著高于其它无性系;徐坑40号无性系蛋氨酸含量极显著高于其它无性系;大源4号无性系酪氨酸含量极显著高于其它无性系;大源2号和大源4号2个无性系的苯丙氨酸和脯氨酸含量极显著高于其它无性系。 同核桃种仁[31]、薄壳山核桃种仁[32]一样,山核桃种仁也富含蛋白质、可溶性糖、矿质元素和各种氨基酸,具有较高的营养和保健价值。山核桃作为浙皖天目山区最重要的特色干果和木本油料树种,对山核桃无性系的筛选评价,在优先考虑丰产性的同时,应对果实形态结构及种仁内含物营养指标进行综合分析与评价。本文对11个山核桃参试无性系果实性状及营养成分研究结果表明:大源4号、高岭8号和高岭4号3个无性系的坚果大小、出籽率、出仁率、果仁质量等均高于其它几个无性系,具有优良的干果利用性状特点,可以作为山核桃干果利用无性系予以重点关注,徐坑89号无性系粗脂肪含量明显高于其它参试无性系,可作为山核桃高档食用油定向开发的重要资源。大源2号和大源4号无性系谷氨酸、精氨酸和天冬氨酸含量相对最高,可以作为氨基酸类产品开发的无性系进行进一步的评价。 参考文献: [1] 张若惠,路安民.中国山核桃属研究[J].植物分类学报,1979,17(2):40-44. [2] 郑万钧.中国树木志 第2卷[M].北京:中国林业出版社,1985:2379. [3] 马良进,林君阳,李 桥,等.山核桃外果皮中的抑菌活性成分[J].林业科学,2009,45(12):90-94. [4] 姚小华.山核桃高效栽培技术[M].北京:金盾出版社,2012:1-9. [5] 王 静,吕芳德.我国山核桃属植物研究进展[J].经济林研究,2012,30(1):138-142. [6] 金志凤,赵宏波,李 波,等.基于GIS的浙江山核桃栽植综合区划[J].浙江农林大学学报,2011,28(2):256-261. [7] 黎章距,钱莲芳,钱光林.山核桃保花保果技术研究[J].林业科学,1993,29(4):360-365. [8] 常 君,姚小华,王开良,等.山核桃异砧嫁接技术研究[J].浙江林业科技,2013,33(6):28-31. [9] 洪 旗,叶浩然,周 燕,等.山核桃本砧嫁接技术研究[J].经济林研究,2013,31(4):203-205. [10] 余 琳,张卫斌,余兵妹.山核桃不同砧穗组合嫁接苗造林效果及结实情况分析[J].南京林业大学学报:自然科学版,2009,33(3):143-145. [11] 王白坡,程晓建,喻卫武,等.山核桃嫁接育苗成活率探讨[J].浙江林学院学报,2002,19(3):231-234. [12] 余 琳,余忠敏,余家中,等.山核桃嫁接育苗技术研究与造林试验[J].浙江林业科技,2006,26(5):20-23. [13] 王冀平,李亚南,马建伟.山核桃仁中主要营养成分的研究[J].食品科学,1998,19(4):44-46. [14] 张 鹏,钟海雁,姚小华,等.四种山核桃种仁含油率及脂肪酸组成比较分析[J].江西农业大学学报,2012,34(3):499-504. [15] 刘 力,龚 宁, 夏国华,等.山核桃种仁蛋白质及氨基酸成分含量的变异分析[J].林业科学研究,2006,19(3):376-378. [16] 葛林梅,郜海雁,穆宏磊,等.山核桃加工过程脂肪酸氧化及抗氧化能力变化研究[J].中国粮油学报,2014,29(1):61-65. [17] 李新委,谢世友,马 燕,等.山核桃营养价值与种植经济效益分析[J].农学学报,2015,5(2):51-56. [18] 吴晓骏,吴 威,赵余庆.山核桃化学成分和生物活性研究进展[J].食品研究与开发,2013,34(12):127-130. [19] 王鸿飞,徐 超,周明亮,等.山核桃油改善小鼠记忆功能的研究[J].中国粮油学报,2012,27(7):63-66. [20] 邵亮亮,徐佳杰,张 亮,等.炒制对山核桃仁营养成分的影响[J].食品科学,2010,31(24):424-426. [21] 余兆硕,丁宏武,唐 琦,等.山核桃油提取工艺优化及脂肪酸组成分析[J].农产品加工,2016, (1):19-23. [22] 邵香君,徐建春,吴家森,等.山核桃集约经营过程中土壤微生物量碳氮的变化[J].水土保持通报,2016,36 (02):72-75. [23] 钱新标,徐温新,张圆圆,等.山核桃果仁微量元素分析初报[J].浙江林学院学报,2009,26(4):511-515. [24] 郑春霞,王文全,阮 晓.新疆干果中微量元素含量分析[J].光谱学与光谱分析,2000,20(4):543-544. [25] 王小生.必需氨基酸对人体健康的影响[J].中国食物与营养,2005,(7):48-49. [26] 解红恩,黄有军,薛霞铭,等.山核桃果实生长发育规律[J].浙江林学院学报,2008,25(4):527-531. [27] 贾晓东,王 婵,莫正海,等.美国山核桃与浙江山核桃果实品质的比较研究[J].天津农业科学,2013,19(3):28-31. [28] 李旭玫.茶叶中的矿质元素对人体健康的作用[J].中国茶叶,2002,24(2):30-31. [29] 章亭洲.山核桃的营养、生物学特性及开发利用现状[J].食品与发酵工业,2006,32(4):90-93. [30] Pellett P L,Young V R.Nutritional evaluation of protein foods[J].Food and Nutrition Bulletin,1980(S4):167. [31] 张 琦,程 滨,赵瑞芬,等.不同品种核桃仁的脂肪酸与氨基酸含量分析[J].山西农业科学,2011,39(11):1165-1169. [32] 于 敏,徐宏化,王正加,等.6个薄壳山核桃品种的形态及营养成分分析[J].中国粮油学报,2013,28(12):74-77. [33] 王 芳,高瑜珑,阮 琴,等.山茶花氨基酸组成分析及营养价值评价[J].浙江师范大学学报:自然科学版,2015,38(3):342-347. [34] Bauer I,Graessle S,Loidl P,etal.Novel insights into the functional role of three protein arginine methyltransferases in Aspergillus nidulans[J].Fungal Genetics and Biology,2010,47(6):551-561. (责任编辑:金立新) DOI:10.13275/j.cnki.lykxyj.2017.01.024 钙浓度对马尾松幼苗生长和生理特征的影响 李德燕1,2,3,周运超1,3* (1.贵州大学贵州省森林资源与环境研究中心,贵州 贵阳 550025; 2.安顺学院,贵州 安顺 561000; 3.贵州大学林学院,贵州 贵阳 550025) 收稿日期: 2016-06-07 关键词:马尾松; Ca2+浓度;生长;生理特性 中图分类号:S791.248 文献标识码:A 文章编号:1001-1498(2017)01-0174-07 Abstract:[Objective]To select the optimal Ca2+concentration forPinusmassonianaseedling growth and to provide evidence for plantation cultivation and rational application of calcium fertilization by studying the effects of Ca2+concentration on growth and physiological characteristics ofP.massonianaseedlings. [Method]A greenhouse sand culture experiment was carried out to study the growth and physiological characteristics of half-year-old seedlings ofP.massonianain different Ca2+concentrations (0.0, 0.4, 1.0, 2.0, 3.0 mmol·L-1and 4.0 mmol·L-1(CK)). [Result]The results showed that both the height and stem increments ofP.massonianaunder 1.0—2.0 mmol·L-1Ca2+for five months were higher, and seedlings grew best in 2.0 mmol·L-1Ca2+. The adaptable strong-to-weak sequence ofP.massonianaseedlings was 2.0, 1.0, 0.4, 3.0, 0.0 mmol·L-1and 4.0 mmol·L-1(CK) of Ca2+. At different growth periods, these indices of pine needles, including O2-generation rate, H2O2content, protective enzyme activities (SOD, POD, CAT and APX), the contents of GSH, AsA and proline, decreased at first and then increased subsequently along with the increasing Ca2+concentration, and all minimums occurred at 1.0 mmol·L-1or 2.0 mmol·L-1of Ca2+. [Conclusion]P.massonianaseedlings is suitable to grow in the soil with Ca2+concentration of 1.0—2.0 mmol·L-1according to the experiment results. Based on comprehensive analysis to soil available calcium ofP.massonianaforestland, the growth and physiological characteristics of seedlings, it is necessary to applying rationally calcium fertilization so as to boost the productivity ofP.massoniana. Keywords:Pinusmassoniana; calcium concentration; growth; physiological properties 马尾松(PinusmassonianaLamb.)作为中国南方的主要造林树种,占中国用材林面积的17.6%,具有较高的经济效益、社会效益和生态效益,为避免马尾松林生产力下降,实现马尾松人工林可持续经营已成为当前的一项迫切任务。然而,长期种植马尾松会造成林地水土流失加重,土壤养分如Ca、Mg和K等流失加快,引起林地养分缺乏,导致马尾松生产力降低[7-8];而周期性采伐使大量养分元素随木材及其他器官的收获而被移出,造成林地养分净消耗,影响了钙在生态系统中的循环,致使钙流失,从而导致地力衰退[9-12]。此外,酸雨作用也会导致林地土壤酸化,使土壤中盐基离子淋失,特别是Ca2+的流失[13-14]。研究结果表明,在马尾松林中土壤钙含量降低可能是导致马尾松林分衰退及生产力降低的重要原因之一。 前人对黔中地区马尾松生长的影响因子进行研究,结果表明,马尾松幼树的株高生长与土壤中CaO、Fe2O3含量及pH值呈负相关,土壤中钙含量对马尾松生长发育影响较大[15]。目前,专门针对钙与马尾松相关的研究主要集中在养分含量分析[16-18]及种源[19]等方面,而有关不同Ca2+水平对马尾松生长及生理特征的影响尚不清楚。本文以半年生马尾松幼苗为研究对象,用不同Ca2+水平的Hoagland营养液对马尾松幼苗进行砂培,研究其生长情况及针叶生理指标的变化特征,掌握马尾松适宜生长的钙浓度范围,阐明不同供Ca2+水平下马尾松生理指标的变化规律及响应机理,为不同Ca2+水平下马尾松人工林培育及合理施用钙肥提供参考。 1.1 材料处理及试验设计 1.2 研究方法 参照汤绍虎等[22]的方法测定功能叶的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性以及丙二醛(MDA)和脯氨酸含量,参照张亚宏等[23]的方法测定抗坏血酸过氧化物酶(APX)活性,参照汤章城[24]和彭建等[25]的方法测定活性氧(ROS)中氧阴离子自由基(O2-)产生速率和过氧化氢(H2O2)浓度,参照Queval等[26]的方法测定抗坏血酸(AsA)及谷胱甘肽(GSH)的含量,参照李合生等[27]的方法测定类胡萝卜素的含量。 1.3 数据处理 试验数据为平均值±标准差(3次重复),使用Excel进行数据统计与作图,SPSS(V20.0)进行数据分析。采用隶属函数法对马尾松生理指标进行综合评定;采用80 d时各处理的生理和生长指标进行相关性分析。 2.1 不同供Ca2+水平下马尾松苗的生长特征 从表1可看出:随着供Ca2+水平的增加,马尾松幼苗株高增量和地径增量均呈先增加后降低的趋势,其中,供Ca2+水平为2.0 mmol·L-1的株高增量和地径增量均最大,分别比CK高51.84%和47.78%,而供Ca2+水平为4.0 mmol·L-1(CK)和0.0 mmol·L-1的株高增量和地径增量最小。供Ca2+水平为2.0 mmol·L-1的株高增量和地径增量除与1.0 mmol·L-1处理的差异不显著外,均显著高于其他处理。 表1 不同供Ca2+水平下马尾松的生长情况Table 1 Growth of P. massoniana seedling under different Ca2+ concentrations 注:同列不同小写字母表示差异显著(P<0.05),下同。 Note: Data with different lowercase letters in the same column are highly significantly different(P<0. 05). The same below. 2.2 不同供Ca2+水平下马尾松针叶生理指标的变化 2.2.1 ROS生成的变化 2.2.1.1 O2-产生速率 图1 A表明:不同供Ca2+水平下,马尾松在相同的生长期内O2-产生速率均呈先降低后升高的趋势。处理20、40、60、80 d时,针叶中O2-产生速率均在2.0 mmol·L-1供Ca2+水平时最低,比CK分别降低了5.12%、5.33%、5.92%和10.02%。处理20 d时,O2-产生速率在0 mmol·L-1供Ca2+水平时最高,且比CK高4.26%;而处理40、60、80 d时,O2-产生速率均在4.0 mmol·L-1供Ca2+水平时最高。相同供Ca2+水平的马尾松针叶内的O2-产生速率随生长期的延长呈不断上升趋势。 2.2.1.2 H2O2含量 图1 B表明:不同供Ca2+水平下,马尾松幼苗针叶内的H2O2浓度随生长期的延长呈不断增加的趋势。处理20、40、60、80 d时,针叶内的H2O2浓度均在1.0 mmol·L-1供Ca2+水平时最低,分别比对照低7.63%、25.75%、28.72%、20.88%;均在4.0 mmol·L-1供Ca2+水平时最高,而0.0 mmol·L-1供Ca2+水平的次之,仅比4.0 mmol·L-1(CK)处理依次低0.67%、2.48%、1.55%和4.87%。 2.2.2 MDA含量的变化 从图2看出:在相同的生长期内,马尾松幼苗针叶内的MDA含量随供Ca2+水平的增加均呈先降低后增加的趋势。处理20、40、60、80 d时,MDA含量均在2.0 mmol·L-1供Ca2+水平最低,分别比4.0 mmol·L-1(CK)下降了11.98%、10.25%、14.69%、13.25%;除处理80 d外,MDA含量的最高值均在0.0 mmol·L-1供Ca2+水平,分别比4.0 mmol·L-1(CK)增加了0.31%、0.14%、0.20%,而处理80 d时,MDA含量最高值在4.0 mmol·L-1(CK)供Ca2+水平。相同供Ca2+水平的马尾松幼苗针叶中的MDA含量随着生长期的延长呈上升趋势。2.2.3 不同供Ca2+水平对马尾松抗氧化系统的影响 图1 不同供Ca2+水平下马尾松针叶O2-产生速率和H2O2含量的变化(图中不同小写字母表示在相同生长期不同供Ca2+水平间差异显著(P<0.05),下同)Fig.1 Change of O2- generation rate and H2O2 content of P. massoniana seedling under different Ca2+ concentrations(Date with different lowercase letters are highly significantly different on the column top(P<0.05).The same below.) 图2 不同供Ca2+水平下马尾松针叶MDA含量的变化Fig.2 Change of MDA content of P. massoniana seedling under different Ca2+ concentrations 图3 不同供Ca2+水平下马尾松SOD、POD、CAT和APX活性的变化Fig.3 Change of SOD, POD, CAT and APX activities of P. massoniana seedling under different Ca2+ concentrations 2.2.4 脯氨酸含量 在相同生长期内,马尾松幼苗针叶中的脯氨酸含量均随供Ca2+水平的增加先降低后增加,最小值均出现在2.0 mmol·L-1Ca2+处理中,在20、40、60、80 d时,分别比4.0 mmol·L-1(CK)降低了5.01%、7.16%、7.92%、18.09%;马尾松幼苗针叶中的脯氨酸含量最大值均出现在0.0 mmol·L-1Ca2+处理中,在20、40、60、80 d时,分别比4.0 mmol·L-1(CK)升高了4.21%、3.90%、4.73%、1.23%。相同供Ca2+水平下,马尾松幼苗针叶内的脯氨酸含量随处理时间的延长总体呈上升趋势,而对照80 d时的脯氨酸含量比60 d时的增加24.27%(图4D)。 图4 不同供Ca2+水平下马尾松AsA、GSH、类胡萝卜素和脯氨酸含量的变化Fig.4 Change of AsA,GSH,carotenoids and proline contents of P.massoniana seedling under different Ca2+ concentrations 2.3 不同供Ca2+水平对马尾松生理影响的综合评价 利用模糊数学的隶属函数法对生理指标进行分析,马尾松幼苗生理指标受不同供Ca2+水平影响由强到弱的供Ca2+浓度顺序为:4.0(CK)>0.0>3.0>0.4>1.0>2.0 mmol·L-1(表3)。 3.1 马尾松生长适宜的钙浓度 表2 马尾松幼苗生理指标与生长的相关性分析Table 2 Correlation analysis between physiological indexes and growth indexes of P.massoniana seedling 注: *、** 分别表示在0.05水平(双侧)和0.01水平(双侧)上显著相关。 Note: *、** means that correlation is significant at the 0.05 level(2-tailed),0.01 level (2-tailed),respectively. 表3 马尾松生理指标的平均隶属函数均值Table 3 Synthetical membership function value ofP. massoniana seedling in physiological indices 3.2 不同供Ca2+水平对马尾松生理的影响 (1)马尾松幼苗在Ca2+水平为1.0 mmol·L-1和2.0 mmol·L-1的株高增量和地径增量较高,其中,Ca2+水平2.0 mmol·L-1的株高增量和地径增量最高。马尾松幼苗对Ca2+水平适应能力强弱顺序为2.0>1.0>0.4>3.0>0>4.0 mmol·L-1(CK)。 (2)在不同生长期内,随着供Ca2+水平的增加,马尾松针叶中ROS(O2-产生速率、H2O2浓度)、MDA含量、抗氧化酶(SOD、POD、CAT和APX)活性、抗氧化物质(GSH、AsA)含量、脯氨酸含量均先降低后增加,最小值均出现在供Ca2+水平1.0 mmol·L-1或2.0 mmol·L-1处理中;类胡萝卜素含量先增加后降低,其最高值均出现在供Ca2+水平2.0 mmol·L-1处理中。 (3)马尾松幼苗的株高增量和地径增量与类胡萝卜素含量关系不明显,而与其他生理指标均呈显著或极显著负相关。 综上,本文研究钙浓度对马尾松生长及生理指标的影响,明确了适宜马尾松生长的钙浓度及生理指标的变化规律,为马尾松人工林培育及合理施用钙肥提供一定的理论支持。 参考文献: [1] McLaughlin S B,Wimmer R.Calcium physiology and terrestrial ecosystem processes[J].The New Phytologist,1999,142(3):373-417. [2] 赵其国.中国东部红壤地区土壤退化的时空变化、机理及调控[M].北京:科学出版社,2002. [3] 周 卫,林 葆.土壤中钙的化学行为与生物有效性研究进展[J].土壤肥料,1996(5):19-22,44. [4] 吴飞华,刘廷武,裴真明.酸雨引起森林生态系统流失研究进展[J].生态学报,2010,30(4):1081-1088. [5] White P J,Broadley M R.Calcium in plants[J].Annals of Botany,2003,92(4):487-511. [6] 陈兴业,叶林茂,张 硌.土壤水分植物生理与肥料学[M].北京:海洋出版社,2010. [7] 唐万鹏,潘 磊,崔鸿侠,等.三峡库区马尾松天然林林分结构特征分析[J].林业科学研究,2015,28(5):681-685. [8] 赵汝东,樊剑波,何园球,等.退化马尾松林下土壤障碍因子分析及酶活性研究[J].土壤学报,2011,48(6):1287-1292. [9] Stupak I,Nordfjell T,Gundersen P.Comparing biomass and nutrient removals of stems and fresh and predried whole trees in thinnings in two Norway spruce experiments[J].Canadian Journal of Forest Research,2008,38:2660-2673. [10] Lattimore B,Smith C T,Titus B D,etal.Environmental factors in woodfuel production:Opportunities,risks,and criteria and indicators for sustainable practices[J].Biomass and Bioenergy,2009,33(10):1321-1342. [11] Schaberg P G,DeHayes D H,Hawley G J.Anthropogenic calcium depletion:a unique threat to forest ecosystem health?[J].Ecosystem Health,2001,7(4):214-228. [12] Tomlinson G H.Acidicdeposition,nutrient leaching and forest growth[J].Biogeochemistry, 2003,65(1):51-81. [13] 凌大炯,章家恩,欧阳颖.酸雨对土壤生态系统影响的研究进展[J].土壤,2007,39(4):514-521. [14] 刘菊秀,周国逸.土壤累积酸化对鼎湖山马尾松林物质元素迁移规律的影响[J].浙江大学学报:农业与生命科学版,2005,31(4):381-391. [15] 周政贤.中国马尾松[M].北京:中国林业出版社,2001. [16] 季孔庶,邹永梅,陈天华,等.不同种源马尾松灰分矿质营养变异的研究[J].南京林业大学学报,1996,20(1):22-25. [17] 郭 峰,周运超.不同密度马尾松林针叶养分含量及其转移特征[J].南京林业大学学报:自然科学版,2010,34(4):93-96. [18] 周运超,王德炉,谌红辉.不同营林措施对马尾松针叶养分的影响[J].浙江林业科技,2001,21(1):5-8. [19] 旷远文,温达志,周国逸,等.鼎湖山异龄马尾松针叶长度序列元素分布[J].植物生态学报,2006,30(1):33-39. [20] 宫杰芳,周运超,李小永,等.马尾松生理指标对高锰胁迫的响应[J].生态学杂志,2012,31(3):520-525. [21] 毛达如.植物营养研究法[M].北京:中国农业大学出版社,2005. [22] 汤绍虎,罗 充.植物生理学实验教程[M].重庆:西南师范大学出版社,2012. [23] 张亚军,孙万仓,魏文慧,等.自交对甘蓝型油菜叶片SOD,CAT,APX活性的影响[J].华北农学报,2008,23(1):105-108. [24] 汤章城.现代植物生理学实验指南[M].北京:科学出版社,1999. [25] 彭 建,王丹英,徐春梅,等.钼酸铵法测定水稻过氧化氢酶活性[J].中国农学通报,2009,25(16):61-64. [26] Queval G,Issakidis-Bourguet E,Hoeberichts F A,etal.Conditional oxidative stress responses in theArabidopsisphotorespiratory mutantcat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression,and define photoperiod as a crucial factor in the regulation of H2O2-induced cell death[J].Plant Journal,2007,52:640-657. [27] 李合生,孙 群,赵世杰,等.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000. [28] 潘 伟,王彦辉,于澎涛,等.石灰石粉混施剂量对酸雨区马尾松细根特征的影响[J].林业科学院研究,2009,22(6):840-845.[29] Gill S S,Tuteja N.Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J].Plant Physiology and Biochemistry,2010,48(12):909-930. [30] 秦晓佳,丁贵杰.磷胁迫对不同种源马尾松氮钾吸收与利用的影响[J].中南林业科技大学学报,2012,34(4):32-36. [31] Bavei V,Shiran B,Arzani A.Evaluation of salinity tolerance in sorghum (SorghumbicolorL.) using ion accumulation,proline and peroxidase criteria[J].Plant Growth Regulation,2011,64:275-285. (责任编辑:徐玉秀) DOI:10.13275/j.cnki.lykxyj.2017.01.025 赤霉素GA4+7处理下白桦无性系生长及 差异基因表达分析 刘 宇,徐焕文,刘桂丰,姜 静* (林木遗传育种国家重点实验室,东北林业大学,黑龙江 哈尔滨 150040) 收稿日期: 2015-12-23 摘要:[目的]为研究赤霉素GA4+7对白桦生长的调控机制。[方法]试验利用1个白桦无性系的分株材料,连续2年分别叶面喷施浓度为0.2 g·L-1(T1)、0.4 g·L-1(T2)的GA4+7溶液,对实验苗木的生长情况进行跟踪调查,并对相应顶端生长组织进行转录组测序和差异基因分析。[结果]对2个年份的苗高、地径等性状分析发现,不同处理间苗高与地径的差异均达到显著或极显著水平,GA4+7对高生长有明显的促进作用,2年后的T2白桦苗高较CK提高了19.65%。于6月28日取GA4+7连续处理2年的白桦顶芽开展RNA-seq分析,结果显示:0.4 g·L-1GA4+7(T2)处理与对照(CK)间的差异基因数量最多,即上调表达的基因有181个,下调表达的基因有55个,这些差异基因在叶绿体类囊体膜、红光、远红光及蓝光的细胞响应等方面富集明显。Pathway富集分析显示:氧化磷酸化途径与光合途径中的8条基因呈上调表达,认为施加0.4 g·L-1GA4+7处理后,上述基因的上调表达加快电子传递进程、催化ATP的合成,从而促进白桦光合作用增强,提高苗期生长量。[结论]对白桦外源施加赤霉素GA4+7后,可上调氧化磷酸化途径和光合途径相关基因的表达,进而促进苗木的高和地径生长。研究结果可为白桦生长相关途径基因的克隆提供参考。 关键字:GA4+7;白桦无性系;转录组;生长 中图分类号:S792.153 文献标识码:A 文章编号:1001-1498(2017)01-0181-09 Abstract: [Objective]To explore the regulation mechanism of gibberellin GA4+7on the growth of birch.[Method]The experiment was carried out by using two treatments from a tissue-cultured plantlet. The leaf surface was treated by foliar spraying GA4+7solution with concentrations of 0.2 g·L-1(T1) and 0.4 g·L-1(T2) for two consecutive years, respectively. Then the differential gene and transcriptome sequencing of the corresponding top growth tissue were analyzed. [Result]It was found that the difference of height and ground diameter of the plantlet under different treatments reached significant or extremely significant level after two years. It is proved that the GA4+7has a significant promoting effect on height growth. The height of birch seedlings under T2 treatment increased by 19.65% compared with the CK. The results of RNA-seq analysis of birch bud for treated by GA4+7for continuous 2 years showed that the number of different gene was the most between the 0.4 g·L-1of GA4+7(T2) treatment and the CK, a total of 181 up-regulated expression genes and 55 down-regulated expression genes were observed. These genes presented significantly enrichment in the chloroplast thylakoid membrane, red light and far red light and blue light cell response. Pathway enrichment analysis showed that 8 genes in the oxidative phosphorylation pathway and photosynthetic pathway were up-regulated, indicating that applying on 0.4 g·L-1GA4+7treatment, the up-regulated gene expression above promoted the electron transfer process, catalyzing ATP synthesis, thereby promoting birch photosynthetic enhancement, and improved seedling growth.[Conclusion]The application of exogenous gibberellin GA4+7can upregulate the expression of related genes, which participates oxidative phosphorylation pathway and photosynthetic pathway, thus contributing to seedling height and diameter growth. Keywords: GA4+7, birch clones, transcriptome, growth 赤霉素是控制植物生长并作用于植物整个生命周期的一种激素,属于四环二萜类化合物。目前已知的赤霉素种类至少有130种,但仅有部分赤霉素的活性较高,GA4与GA7是其中活性很高的两种。由于这两种赤霉素分子结构相似,分离困难,常以混合物即GA4+7的形式存在[1]。迄今为止,有关GA4+7影响树木生长报道最多的是促进杉科(Taxodiaceae)、松科(Pinaceae)等针叶树种开花结实的研究[2-5]。如Almqvist等[6]人采用GA4+7茎干注射处理21年生的挪威云杉(Piceaabies),显著增加了雌雄花的数量;赵鹏等[7]对15年生落叶松喷洒0.5 g·L-1的赤霉素GA4+7,喷洒的枝条雌球花量是对照枝条的2.9倍。另外,在果树中研究发现,GA4+7有效促进植株营养生长,并且大幅提高开花率及座果率[8-9]。目前,GA4+7在植物生长调节和促进开花结实两方面所起的作用已被证实,但是在国内利用GA4+7促进阔叶用材树种的生长相关报道还较少,主要集中在施加GA3或其它易于提取的赤霉素,研究其对植物的生长影响,其主要原因是由于GA4+7难以获得,且生产价格昂贵,限制了其广泛应用[1],所以GA4+7对于阔叶树种的处理方式、处理时间以及作用效果尚未明确。但随着GA4+7合成途径的确定,生产菌株的获得,使其使用成本大幅减少,大大推动GA4+7在针阔叶树种中的应用。因此本试验以1个白桦(B.platyphylla×B.pendula)无性系为研究对象,采用不同浓度赤霉素GA4+7处理,在生长最旺盛的6月末取材测定基因的转录组,目的是为了研究GA4+7对白桦生长的影响及分子机制,也为GA4+7在白桦中的应用提供参考。 1.1 材料 1.2 方法 参考Philipson的方法[10]设置0.2 g·L-1GA4+7(T1处理)、0.4 g·L-1GA4+7(T2处理)2种浓度,以不喷施任何溶液为空白对照(CK),每个处理9株,共计27株白桦无性系。2014年4月20日至7月20日期间对参试苗木进行叶面喷施处理,每隔15 d处理1次,10月初待苗木封顶时调查1年生苗高、地径。2015年4月20日开始重复上述GA4+7处理,并于2015年6月28日分别取T1、T2、CK的白桦顶端组织(包含顶芽及第一片幼叶),迅速置于液氮中带回实验室。10月初待苗木封顶时调查2年生苗高、地径。 1.2.1 RNA的提取及cDNA文库的构建 采用CTAB法提取样品总RNA后,用DNaseⅠ(Promega,USA)消化去除DNA污染,采用Qubit 2.0、Aglient 2100方法检测RNA样品的纯度、浓度和完整性,从3个RNA样品中各取20 μg由百迈克生物科技有限公司进行cDNA文库的构建。分别使用Qubit2.0和Agilent 2100 Bioanalyzer对文库的浓度和插入片段大小进行检测,使用Q-PCR方法对文库的有效浓度进行准确定量,以保证文库质量。库检合格后,用HiSeq2500进行测序并进行有参考基因组的转录组分析。 将测序所得Raw Data去除测序接头以及引物序列,过滤低质量值数据,获得高质量的Clean Data。将其与白桦基因组进行比对,比对上的reads称为Mapped Data。通过检验插入片段在基因组上的分布,评估mRNA片段化的随机性及mRNA的降解情况;通过插入片段的长度分布,评估插入片段长度的离散程度;通过绘制饱和度图,评估文库质量。 1.2.2 差异基因的筛选、GO、pathway分析 使用EBSeq进行差异表达分析,获得2个样品之间的差异表达基因集,采用RPKM(Reads Per Kb per Million reads)作为衡量转录本或基因表达水平的指标[11],计算得到的基因表达量可直接用于比较不同样品间的基因表达差异。在差异表达基因检测过程中,将Fold Change≥2且FDR(false discovery rate)<0.01作为筛选标准定义差异表达基因。得到差异表达基因之后,对差异表达基因做GO功能分析和KEGG Pathway分析。 RPKM计算公式为: RPKM = 106C/(NL /103) 设RPKM(A)为geneA的表达量,则C为唯一比对到gene A的reads数,N为唯一比对到所有gene的总reads数,L为gene A的碱基数,RPKM法能消除基因长度和测序量差异对计算基因表达的影响。 1.2.3 qRT-PCR验证 采用CTAB法分别提取T1、T2以及CK白桦顶端分生组织的总RNA,经DNaseI(RNase free)消化后使用ReverTre Ace®qPCR RT Kit(Toyobo,Osaka,Japan)将提取的总RNA进行反转录。反转录产物稀释10倍,用作定量PCR的模板,以BpUBC和BpSAND作为内参基因,进行实时定量PCR 扩增(所有试样进行3次重复)。PCR的反应体系为:SYBR® Premix ExTaqTMⅡ(2×)10 μL,引物各1 μL(10 μmol·L-1),水6 μL,模板2 μL,反应参数为95℃预变性30 s,95℃变性5 s,58℃退火延伸34 s,循环40 次,绘制溶解曲线,温度由95℃ 15 s,60 ℃ 1 min,至95℃ 15 s止。以上反应在ABI PRISM® 7500 荧光定量PCR仪上完成。选取未处理白桦(CK)作为对照,用2-ΔΔCT方法[12]对定量PCR的结果进行分析,引物序列见表1。 1.3 遗传参数分析 运用MicrosoftExcel进行数据处理,用SPSSv16.0软件进行方差分析及多重比较。 2.1 不同浓度赤霉素GA4+7对白桦无性系生长的影响 对GA4+7处理白桦无性系获得的2个年度生长性状进行方差分析以及主要遗传参数分析(表2)。在第1个年度,发现不同浓度的GA4+7对苗高的生长有极显著促进作用(P<0.01),而对地径的影响不明显(P>0.05);在连续处理的第2个年度,GA4+7不但延续了对高生长的极显著促进作用(P<0.01),而且对地径生长的促进作用也达到了显著水平(0.01 表1 qRT-PCR引物序列Table 1 qRT-PCR primer sequence 对各处理的生长性状进行多重比较发现(见图1),白桦的苗高生长在各处理间的差异达到显著水平(0.01 2.2 不同浓度GA4+7处理下白桦转录组数据统计及文库质量评价 对CK、T1、T2等白桦样品测序数据评估显示,Q30碱基百分比均不低于91.94%,并且3个样品与基因组的比对率在79.00%以上,说明转录组数据满足后续信息分析的需求及数据分析的可靠性。通过对插入片段在基因组上的分布及插入片段的长度分布进行检测、饱和图的绘制,表明该转录组文库可以用于后续转录组测序。 表2 赤霉素GA4+7处理白桦无性系苗高、地径方差分析及主要遗传参数Table 2 Analysis of variance and genetic parameters among gibberellin GA4+7 treatment birch clones on height and base diameter 注:图中a、b和c表示差异显著程度,不同字母表示差异显著(n =3,P<0.05,Duncan多重比较分析,误差线表示标准偏差),相同字母表示差异不显著Note: a, b and c in the figure showed significant difference degree, different letters mean significant difference (n=3, P<0.05, Duncan multiple comparison analysis, error line shows the standard deviation), the same letter indicates no significant difference.图1 赤霉素GA4+7处理白桦无性系苗高、地径生长情况Fig.1 Height and base diameter growth of gibberellin GA4+7 treatment birch clonal 表3 样品测序数据统计及比对效率Table 3 Samples sequencing data statistics and mapped ratio 2.3 不同浓度GA4+7处理下白桦无性系的差异表达基因 根据RPKM获得CK、T1、T2处理的白桦3个样本转录组文库的基因表达量,比较每2样本间的差异表达基因(图2),CK_vs_T1有145个差异基因(95个上调,50个下调),CK_vs_T2有236个差异基因(181个上调,55个下调),T1_vs_T2有82个差异基因(46个上调,36个下调),其中T1_vs_T2的差异基因最少。绘制维恩图分析3个处理差异表达基因的分布(图3),发现有6个基因在CK_vs_T1、CK_vs_T2、T1_vs_T2间差异表达,有29个基因在CK_vs_T1和T1_vs_T2中差异表达,有33个基因在CK_vs_T2和T1_vs_T2中差异表达,有73个基因在CK_vs_T1和CK_vs_T2中差异表达。 图2 不同处理间上调和下调差异基因数量Fig.2 The number of up-regulated and down-regulated differential expressed genes between the different treatments 2.4 差异基因的GO分类及富集分析 GO功能显著性富集分析能确定差异表达基因行使的主要生物学功能,可将基因分为细胞组成、生物过程、分子功能3个大类,因此对3个处理的差异表达基因进行了功能分类,结果见图4。差异表达基因主要集中在细胞组成的细胞(cell)、细胞器(organelle)、细胞部分(cell part);分子功能的催化活性(catalytic activity)、结合(binding);生物过程的代谢过程(metabolic process)、细胞过程(cellular process)、单一有机体过程(single-organism process)、刺激响应(response to stimulus)、生物调控(biological regulation)。由图2可知,CK_vs_T1的差异基因数量明显少于CK_vs_T2的差异基因数量,说明施加高浓度的GA4+7引起的差异基因数目增多。 其中G0: CK_vs_T1;G1:CK_vs_T2;G2:T1_vs_T2 注:细胞组成:C1 胞外区,C2 细胞,C3 拟核,C4 细胞膜,C5 病毒体,C6 细胞连接,C7 细胞外基质,C8 膜包围的内腔,C9 高分子复合物,C10 细胞器,C11 细胞外部分基质,C12 细胞外部分基质,C13 部分细胞器,C14 部分病毒体,C15 部分细胞膜,C16 部分细胞。分子功能:M1 蛋白结合转录因子活性,M2 核酸结合转录因子活性,M3 催化活性,M4 受体活性,M5 甲脒基核苷酸交换因子活性,M6 结构因子活性,M7 运输活性,M8 结合,M9 电子载体活性,M10 抗氧化活性,M11 通道调控活性,M12 金属伴侣活性,M13 酶调节活性,M14 标签蛋白,M15 翻译调节活性,M16 营养容量活性,M17分子翻译活性。生物过程:B1 复制,B2 细胞杀伤,B3 免疫系统过程,B4 代谢过程,B5 细胞过程,B6 生殖过程,B7 生物附着,B8 信号,B9 多细胞有机体过程,B10 发育过程,B11 生长,B12 移动,B13 单一有机体过程,B14生物学时期,B15 节律过程,B16 响应刺激,B17 定位,B18 多有机体过程,B19 生物调节,B20 细胞组成组织起源。Note: Cellular component: extracellular region, cell, nucleoid, membrane, virion, cell junction, extracellular matrix, membrane-enclosed lumen, macromolecular complex, organelle, extracellular matrix part, extracellular region part, organelle part, virion part, membrane part, cell part. Molecule function: protein binding transcription factor activity, nucleic acid binding transcription factor activity, catalytic activity, receptor activity, guanyl-nucleotide exchange factor activity, structural molecule activity, transporter activity, binding, electron carrier activity, antioxidant activit , channel regulator activity, metallochaperone activity, enzyme regulator activity, protein tag, translation regulator activity, nutrient reservoir activity, molecular transducer activity. Biological process: reproduction, cell killing, immune system process, metabolic process, cellular process, reproductive process, biological adhesion, signaling, multicellular organismal process, developmental process, growth, locomotion, single-organism process, biological phase, rhythmic process, response to stimulus, localization, multi-organism process, biological regulation, cellular component organization or biogenesis.图4 不同样品间表达差异基因的GO分类Fig.4 GO classification of differentially expressed genes between the different birch samples 对每2个处理样品间的差异基因进行GO富集分析,以CK_vs_T2为例(图5),差异基因在叶绿体类囊体膜(chloroplast thylakoid membrane)、红光的细胞响应(cellular response to red light)、远红光的细胞响应(cellular response to far red light)、蓝光的细胞响应(cellular response to blue light)等方面富集显著。 注:分子功能:醌结合活性、杜松烯合酶活性、电子运输转运活性、右旋大根香叶烯合酶活性、germacradienol合酶活性、叶绿素结合;细胞组成:质体球、叶绿体类囊体膜;生物过程:光系统Ⅱ组装、光合作用光反应、杜松烯生物合成过程、花青素组成、卡里金响应、高亮度细胞响应、红光响应、紫外光的细胞响应、光保护、红光的细胞响应、远红光的细胞响应、蓝光的细胞响应。Note: Molecule function: quinone binding,(+)delta cadinene synthase activity, electronic transporter transfer activity, germacrene-D synthase activity, germacradienol synthase activity,chlorophyⅡ binding; Cellular component: plastoglobule, chloroplast thylakoid membrane; Biological process: photosysemⅡassembly, photosynthesis light reaction, cadinene biosynthesis process ,anthocyanin-containing compound ,response to karrikin ,cellular response to high light ,response to red light ,cellular response to UV-A ,photoprotection ,cellular response to red light, cellular response to far red light, cellular response to blue light.图5 CK_vs_T2差异基因的富集分析Fig.5 Analysis of CK_vs_T2 difference gene enrichment 2.5 赤霉素处理下白桦无性系生长相关基因的初步分析 Pathway显著性富集分析能确定差异表达基因参与的主要生物过程,因此用KEGG数据库对差异基因进行Pathway富集分析,研究富集的主要生物学过程。 2.5.1 光合作用途径相关基因 根据白桦无性系生长情况发现,GA4+7处理对白桦苗期高生长有显著促进作用,且0.4 g·L-1GA4+7对白桦的促进作用较0.2 g·L-1的促进作用更显著。由于植物生物量与光合作用呈正相关关系[14],因此对光合作用途径相关基因进行深入挖掘。Pathway富集分析结果显示:GA4+7处理白桦苗木后,CK_vs_T2的光合作用途径有4个差异基因上调表达,基因的表达量见图6,这些差异基因编码的蛋白分别是光系统Ⅱ蛋白D1(photosystem II protein D1)、光系统ⅠP700叶绿素A脱辅基蛋白A2(photosystem I P700 chlorophyll A apoprotein A2)、细胞色素b6f亚组4(cytochrome b6-f complex subunit 4)、叶绿体中CF1ɑ亚组ATP合成酶(ATP synthase CF1 alpha subunit (chloroplast))。 注:BP028367编码光系统Ⅱ蛋白D1;BP028372编码光系统ⅠP700叶绿素A脱辅基蛋白A2; BP028382编码细胞色素b6f亚组4;BP029736编码叶绿体中CF1ɑ亚组ATP合成酶。Note: BP028367 Ⅱcoding photosystem II protein D1;BP028372 coding photosystem I P700 chlorophyll A apoprotein A2;BP028382 coding cytochrome b6-f complex subunit 4;BP029736 coding ATP synthase CF1 alpha subunit (chloroplast)图6 光合作用途径相关基因在不同样品间的表达水平Fig.6 Related gene expression levels among different samples in photosynthesis pathway 2.5.2 氧化磷酸化途径相关基因 氧化磷酸化途径是有机物在分解过程中的氧化步骤所释放的能量,驱动ATP合成的过程,即ATP的生成过程,ATP是植物的各项生命活动直接能源物质[15]。Pathway富集结果表明:CK_vs_T2的氧化磷酸化途径有4个差异基因上调表达,基因的表达量见表4,它们分别编码ATP合成酶F1亚组1(ATP synthase F1 subunit 1)、细胞色素c氧化酶亚组1(cytochrome oxidase subunit 1)、NADH-质体醌氧化还原酶亚组K(NAD(P)H-quinone oxidoreductase subunit K)、NADH锟氧化还原酶亚组2(NAD(P)H-quinone oxidoreductase subunit 2)。 2.5.3 qRT-PCR验证 为验证转录组测序结果的可靠性,分别以T1、T2和CK白桦顶尖组织的RNA为试材,对光合作用途径表达差异显著的4条基因和氧化磷酸化途径表达差异显著的4条基因进行qRT-PCR分析,结果显示:与CK相比,T1、T2中上述8条基因均呈上调表达,与Illumina/Solexa测序结果具有相同的表达趋势(图7),说明测序结果是可靠的。 表4 氧化磷酸化途径相关基因在不同处理间的 表达水平(RPKM值)Table 4 Related gene expression levels between different treatment in oxidative phosphorylation pathway (RPKM value) 注:相对表达水平由实时定量PCR以BpUBC和BpSAND为内参计算而来,每个基因3次重复。Note: the relative expression level by real-time quantitative PCR with BpUBC and BpSAND as reference genes to calculated, repeat 3 times each gene.图7 实时定量PCR验证各样品之间候选基因的表达差异Fig.7 Real-time quantitative PCR to verify differentially expressed between each candidate gene samples 调节树木生长,使其短时间内获得较大生长增益从而缩短育种周期是林木育种的重要课题,人们通常采取施肥、环割、断根、施加外源激素等措施促进树木生长[7,16-18]。试验证明,对于果材兼用树种施加GA4+7在促进开花及生长、提高座果率方面效果显著,因此,备受育种工作者的重视[8-9]。 为了探讨GA4+7在促进白桦生长方面的作用,试验利用遗传组成完全相同的1个白桦无性系为材料,分别喷施0.2 g·L-1和0.4 g·L-1GA4+7,结果表明,施加GA4+7对白桦苗期生长具有促进作用,在苗高方面,2个浓度的GA4+7均能显著促进苗高生长,0.2 g·L-1和0.4 g·L-1GA4+7处理2年后无性系苗高均值分别高于CK的9.05%、19.65%;在地径方面,在GA4+7连续处理2年后地径均值显著高于CK,GA4+7浓度为0.4 g·L-1时对白桦苗期生长促进效果好。这一实验结果与Bottini等人[9]对苹果(Maluspumila)树的研究结果相吻合,即赤霉素促进植株的营养生长。 GA4+7处理下白桦无性系生长量的增加,必然与生长相关基因的上调表达密切关联,对白桦苗木生长旺盛期的转录组测序也证明了0.4 g·L-1GA4+7(T2)处理苗高增长显著,同时与对照(CK)之间差异基因的数量也最多,即上调表达的基因有181个,下调表达的基因有55个(图2),这些差异基因在叶绿体类囊体膜、红光、远红光及蓝光的细胞响应等方面富集明显(图5),进一步Pathway富集分析发现:叶绿体中CF1ɑ亚组ATP合成酶、细胞色素b6f亚组4、光系统Ⅱ蛋白D1、光系统ⅠP700叶绿素A脱辅基蛋白A2等4条光合代谢相关的基因呈上调表达,这些基因参与光合作用的光反应过程。另外还有氧化磷酸化途径的ATP合成酶F1亚组1、细胞色素c氧化酶亚组1、NADH-质体醌氧化还原酶亚组K、NADH锟氧化还原酶亚组2等4条基因呈上调表达,上述8条基因表达量的升高,可能是通过加快电子传递过程、催化ATP的合成,而促进植物的光合作用增强[19-25]。总之,GA4+7处理后白桦通过上调光合途径和氧化磷酸化途径的相关基因的表达,加快其细胞内有机物的合成,进而促进其苗高、地径的生长。由于参试的白桦无性系放置于塑料大棚外,按常规管理这些苗木进入开花结实期还需2~3年的时间,GA4+7处理后能否加快白桦营养生长向生殖生长转变进程还有待于后续研究,按照前期试验设计继续施加GA4+7,观察试验组是否缩短童期、是否增加开花数量提高种子产量,进而在基因表达水平上探讨GA4+7对白桦生长的影响。 本研究外源施加赤霉素GA4+7可促进白桦苗木的高生长和地径生长。转录组测序结果表明:赤霉素GA4+7处理前后的差异基因主要富集在叶绿体类囊体膜、红光、远红光及蓝光的细胞响应等方面;同时赤霉素GA4+7可上调氧化磷酸化途径和光合途径相关基因的表达,研究结果可为深入了解赤霉素GA4+7对白桦生长的调控机制及生长相关途径重要基因的克隆提供参考。 参考文献: [1] 李佼佼,杨文革,胡永红.赤霉素A4+7的分子生物学研究进展[J]. 河南农业科学,2013,42(5):1-4. [2] Paula M P. Eastern white pineflowering in response to spray application of gibberellin A4/7or ProConeTM[J]. Northern Journal of Applied Forestry, 2002,19(2):68-72. [3] Cherry M L,Anekonda T S,Albrecht M J,etal. Flower stimulation in young miniaturized seed orchards of Douglas-fir(Pseudotsuga menziesii)[J]. Canadian Journal of Forest Research, 2007,37(1):1-10. [4] Owens J N,Chandler L M,Bennett J S,etal. Cone enhancement inAbiesamabilisusingGA4/7, fertilizer, girdling and tenting[J]. Forest Ecology and Management, 2001,154(2001):227-236. [5] 邵顺流,陈小荣,唐陆法,等.GA4/7对百山祖冷杉开花促进和子代遗传活力恢复研究[J]. 浙江林业科技,2007,27(5):21-24. [6]Almqvist C. Timing ofGA4/7application and the flowering ofPinussylvestrisgrafts in the greenhouse[J].Tree Physiology, 2003,23(6):413-418. [7]赵 鹏.油松无性系种子园开花结实习性及激素处理促进成花的研究[D].杨凌:西北农林科技大学,2007:38-46. [8]Curry E. Increase in epidermal planar cell density accompanies decreased russeting of ‘golden delicious’ apples treated with Gibberellin A4+7[J]. HortScience, 2012,47(2):232-237. [9]Bottini R,Cassán F,Piccoli P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase[J]. Appl Microbiol Biotechnol,2004,65(5):497-503. [10] Philipson J J. The promotion of flowering in large field-grownSitkaspruceby girdling and stem injections of gibberellin A4/7[J].Canadian Journal of Forest Research, 1985,15(1):166-170. [11]Mortazavi A,Williams B A,McCue K,etal. Mapping and quantifying mammalian transcriptomes by RNA-Seq[J]. Nature Methods, 2008,5(7):621-628. [12] Livak K J,Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCtmethod[J].Methods, 2001,25(4):402-408. [13]刘 宇,徐焕文,姜 静,等.基于种子活力及苗期生长性状的白桦四倍体半同胞家系初选[J]. 北京林业大学学报,2014,36(2):74-80. [14] 孔国辉,刘世忠,吴 彤,等.油页岩废渣场26种木本植物光合作用和生长的差异[J].热带亚热带植物学报,2006,14(6):467-476. [15]潘瑞炽主编.植物生理学[M]第6版.北京:高等教育出版社,2008:116-119. [16]杨传平,刘桂丰,魏志刚,等.白桦强化促进提早开花结实技术的研究[J].林业科学,2004,40(6):75-78. [17]刘福妹,姜 静,刘桂丰.施肥对白桦树生长及开花结实的影响[J]. 西北林学院学报, 2015,30(2):116-120. [18]Daoust G, Plourde A, Beaulieu J. Influences of crown size and maturation on flower production and sex expression in Picea glauca treated with gibberellin A4/7[J]. Tree Physiology, 1995,15(7-8):471-475. [19]Anbudurai P R,Mor T S,Ohad I,etal. The ctpA gene encodes the C-terminal processing protease for the D1 protein of the photosystem II reaction center complex[J]. Proceedings of the National Academy of Sciences, 1994,91(17):8082-8086. [20]Russell A W,Critchley C,Robinson S A,etal. Photosystem II regulation and dynamics of the chloroplast D1 proteinin Arabidopsis leaves during photosynthesis and photoinhibition[J].Plant Physiology, 1995,107(3):943-952. [21]Kurisu G,Zhang H M,Smith J L,etal. Structure of the cytochrome b6f complex of oxygenic photosynthesis: tuning the cavity[J]. Science, 2003,302(5647):1009-1014. [22]Scheller H V,Jensen P E,Haldrup A,etal. Role of subunits in eukaryotic Photosystem I[J]. Biochimica et Biophysica Acta (BBA)-Bioenergetics, 2001,1507(1):41-60. [23]Meng B Y,Tanaka M,Wakasugi T,etal. Cotranscription of the genes encoding two P700 chlorophyll a apoproteins with the gene for ribosomal protein CS14: determination of the transcriptional initiation site by in vitro capping[J]. Current Genetics, 1988,14(4):395-400. [24]Nielsen J,Hansen F G,Hoppe J,etal. The nucleotide sequence of the atp genes coding for the F0subunits a,b,c and the F1subunit δ of the membrane bound ATP synthase of Escherichia coli[J]. Molecular and General Genetics MGG, 1981,184(1):33-39. [25]杨建国.杨树叶绿体NADH脱氢酶基因的克隆与序列分析[D].太谷:山西农业大学,2014:19-24. (责任编辑:张 研) Stoichiometric Characteristics of C, N and P in the Dominant Species Sapling Organs ofCastanopsisfissaNatural Forest in Northern Fujian ZHENGDe-xiang1,CAIYang-xin1,YANGYu-jie1,ZHONGZhao-quan2,MIAOSan-hua1,WUWen-bin1 (1.College of Forestry, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China;2.State-owned Forest Farm of Shunchang, Shunchang 353200, Fujian, China) [Objective]Sample plots were set inCastanopsisfissanatural forest in Northern Fujian. The saplings of four dominant species were used to study the stoichiometric characteristics of C, N and P in different organs, so as to lay a theoretical basis for the regeneration and protection ofC.fissanatural forest. [Method]The mass fractions of C, N and P in stem, root, branch and leaf of the sampling trees were determined with carbon-nitrogen analyzer and HNO3-HClO4heating digestion method. Based on variance analysis of C, N and P contents in different organs, the stoichiometric characteristics of C, N and P were studied with the comparison of the changing stoichiometric ratio. [Result]The contents of C, N and P in leaf were higher than in other organs, whereas those in root and stem were lower for the saplings of dominant species ofC.fissanatural forest in Northern Fujian. Moreover, the C∶P ratio in different organs was higher than the C∶N ratio and N∶P ratio. The N∶P ratio was the minimum and varied among different tree species. There were highly significant differences in the contents and the stoichiometric characteristics of C, N and P in different organs of the tree species. [Conclusion]Compared with N and P, the C content was higher in different organs of the dominant tree species inC.fissanatural forest in Northern Fujian. The C∶N ratio and C∶P ratio were higher than the average level while the N∶P ratio was higher than 16, indicating that the nutrient utilization efficiency and growth rate of the saplings of dominant species inC.fissanatural forest were suboptimal. The growth and the regeneration of the saplings were influenced by the content of N and P and restricted by the missing of P. natural forest; young tree; organ; nutrient element; stoichiometric;Castanopsisfissa Screening of Fungicides to Pathogens of Persimmon Anthracnose DENGQuan-en1,DINGXiang-yang2,XUJian-qiang3,LIJian-an1,GUZhan-ying1 (1.Central South University of Forestry and Technology, Key Laboratory of Non-Wood Forest Product of State Forestry Administration, Key Laboratory of Cultivation and Protection for Non-Wood Forest Trees of Ministry of Education, Changsha 410004, Hunan, China; 2.Henan Academy of Forestry, Zhengzhou 450008, Henan, China; 3.Henan University of Science and Technology, Luoyang 471003, Henan, China) Nut Fruit Characteristics and Nutrients ofCaryacathayensisClones CHANGJun1,RENHua-dong1,YAOXiao-hua1,WANGKai-liang1,ZHOUYan2,ZHOUZhen-qi2 (1.Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Key Laboratory of Tree Breeding of Zhejiang Province,Hangzhou 311400, Zhejiang, China; 2.Jiande Forestry Bureau, Jiande 311600, Zhejiang, China) Effects of Calcium Concentration on Growth and Physiological Characteristics ofPinusmassonianaSeedling LIDe-yan1,2,3,ZHOUYun-chao1,3 (1.Research Center of Forest Resources and Environment in Guizhou Province, Guizhou University, Guiyang 550025, Guizhou, China; 2.College of Agronomy, Anshun University, Anshun 561000, Guizhou, China; 3.College of Forestry, Guizhou University, Guiyang 550025, Guizhou, China) Analysis of Clonal Growth and Differences in Gene Expression ofBetulaplatyphylla×B.pendulaunder GA4+7Treatment LIUYu,XUHuan-wen,LIUGui-feng,JIANGJing (State Key Laboratory of Tree Genetics and Breeding, Northeast Forestry University, Harbin 150040, Heilongjiang, China) 10.13275/j.cnki.lykxyj.2017.01.021 2016-05-03 福建省教育厅资助项目(JA12112);福建农林大学校重点项目建设专项(6112C035H)资助 郑德祥(1975—),男,福建仙游人,博士,副教授,主要从事森林经理学研究.E-mail:fjzdx@126.com S718.55 A 文章编号:1001-1498(2017)01-0154-06 63.1 文献标识码:A 1001-1498(2017)01-0160-06 文献标识码:A 文章编号:1001-1498(2017)01-0166-085 结论
基金项目: 国家公益性行业(农业)科研专项“现代柿产业关键技术研究与试验示范”(201203047)。
作者简介: 邓全恩(1987—),男,河南商丘人,在读博士研究生,主要从事经济林栽培研究.
* 通讯作者:丁向阳,研究员,主要从事柿子育种及栽培研究. E-mail:dxydd001@163.com1 材料与方法


2 结果与分析
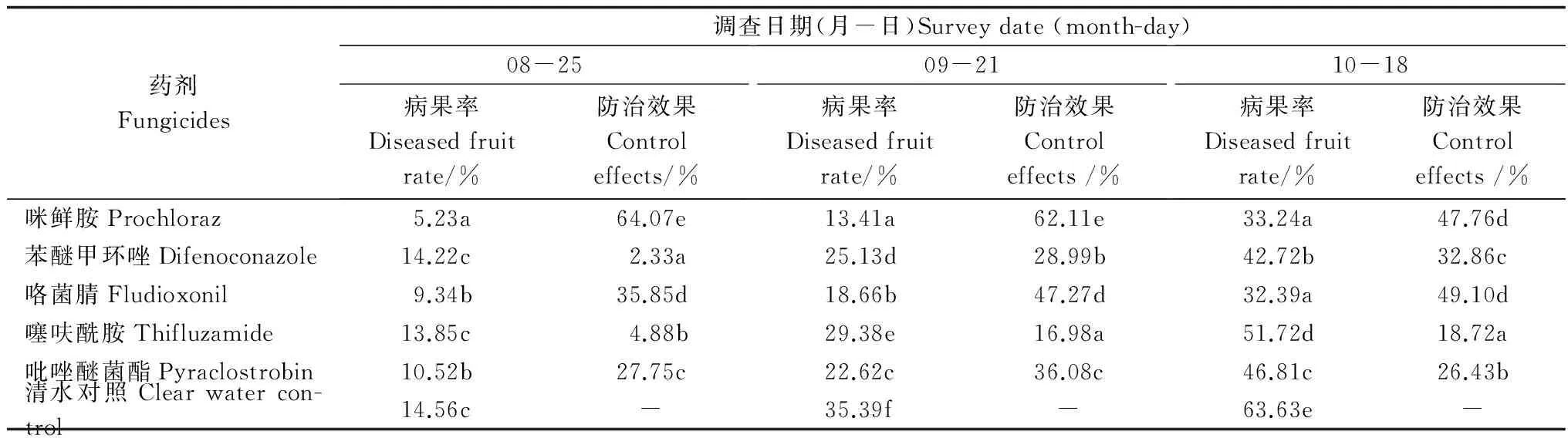
3 讨论
4 结论
基金项目: “十二五”国家科技支撑计划(2013BAD14B0104);国家林业局科技发展中心山核桃遗传资源调查项目(GR-2015-16)。
作者简介: 常 君(1981—),男,山西怀仁人,助理研究员,硕士,从事经济林栽培与育种研究.
* 通讯作者:任华东(1963—),男,浙江永康人,副研究员,从事经济林栽培与育种研究. E-mail:renhd@163.com
1 材料与方法
2 结果与分析



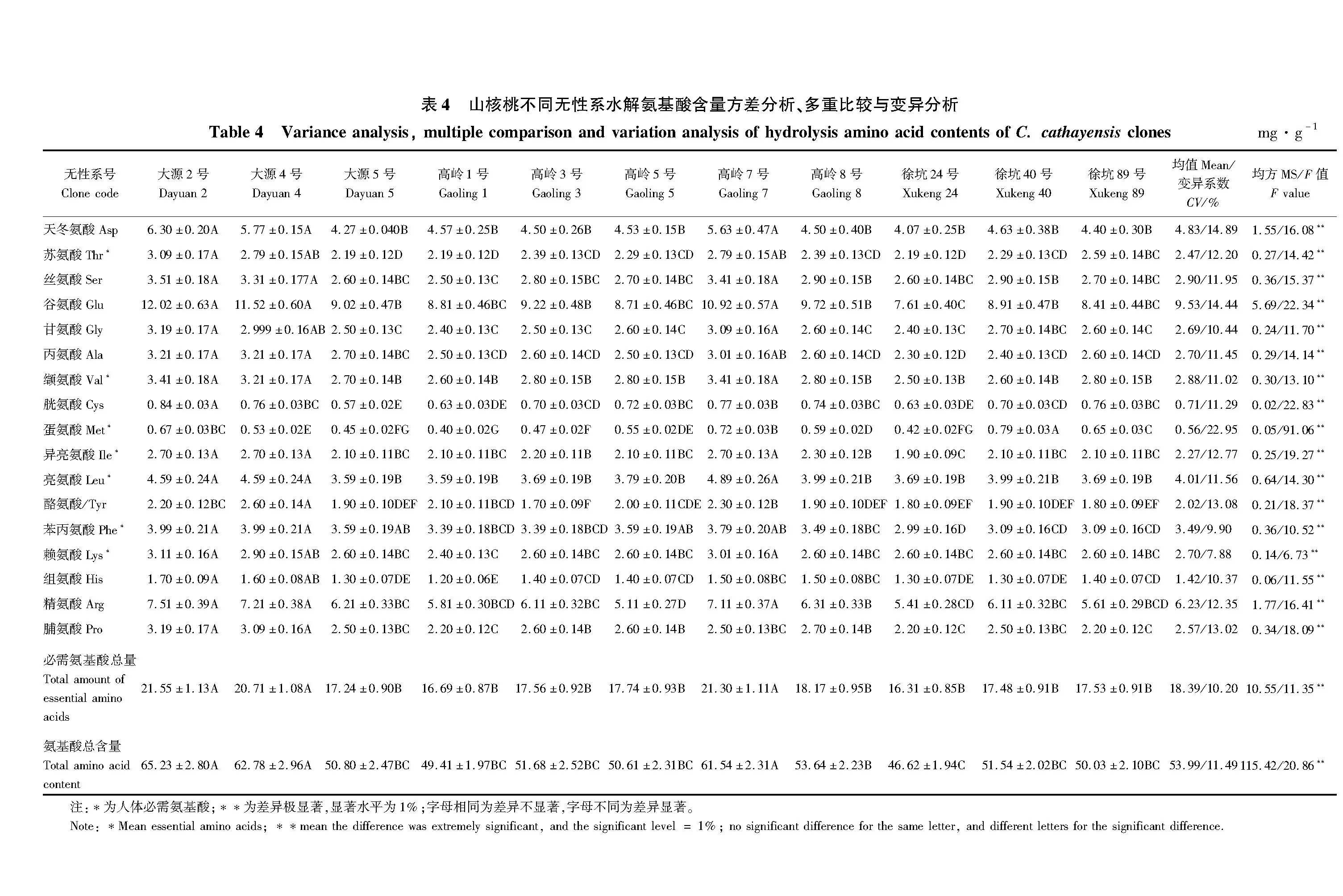
3 讨论
4 小结
基金项目: 贵州省重大专项“马尾松多目标培育及产业关键技术研究与示范”(黔科合重大专项字(2012)6001号);“百”层次人才计划(黔科合人才(2015)4022)
作者简介: 李德燕(1978—),女,贵州福泉人,博士研究生,主要从事植物生理生态方面的研究.E-mail:lideyan1234@163.com
* 通讯作者:周运超(1964—),教授,博导.E-mail:yczhou@gzu.edu.cn1 材料与方法
2 结果与分析

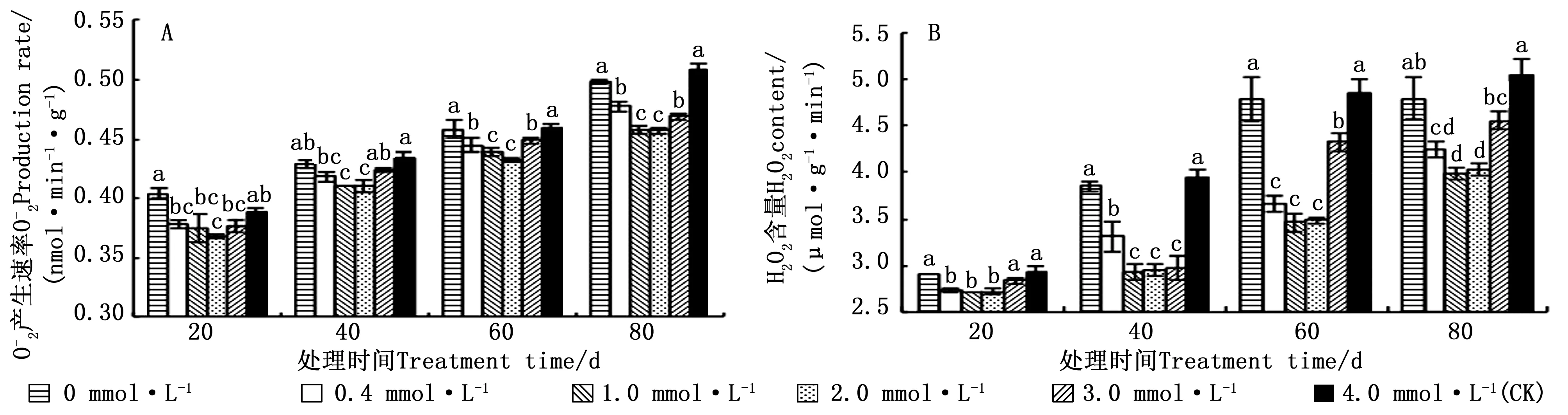
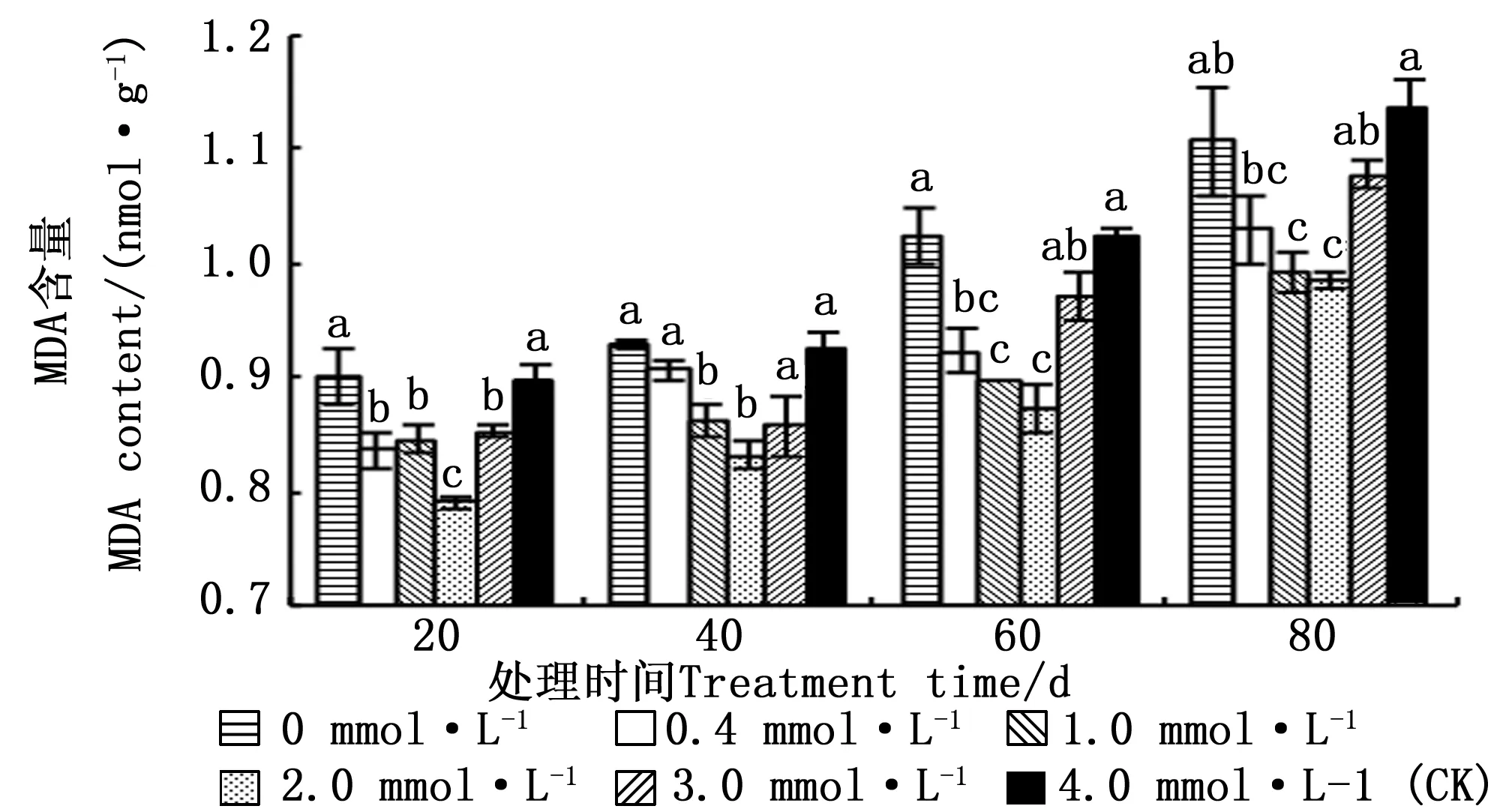



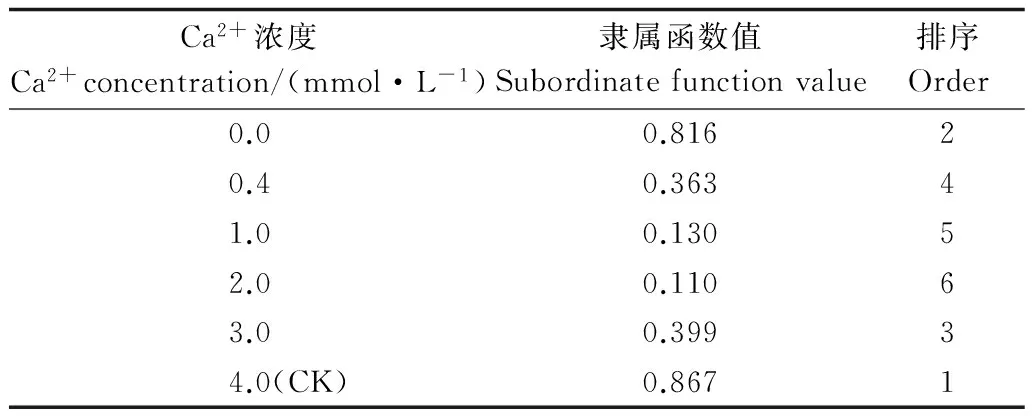
3 讨论

4 结论
基金项目: 珍贵用材树种新品种选育技术研究(2012BAD01B05)。
作者简介: 刘 宇(1990—),男(汉族),黑龙江齐齐哈尔人,硕士研究生,主要从事林木遗传育种研究
* 通讯作者:姜 静(1960—),女(汉族),黑龙江哈尔滨人,教授,主要从事林木遗传育种研究。1 材料与方法
2 结果与分析




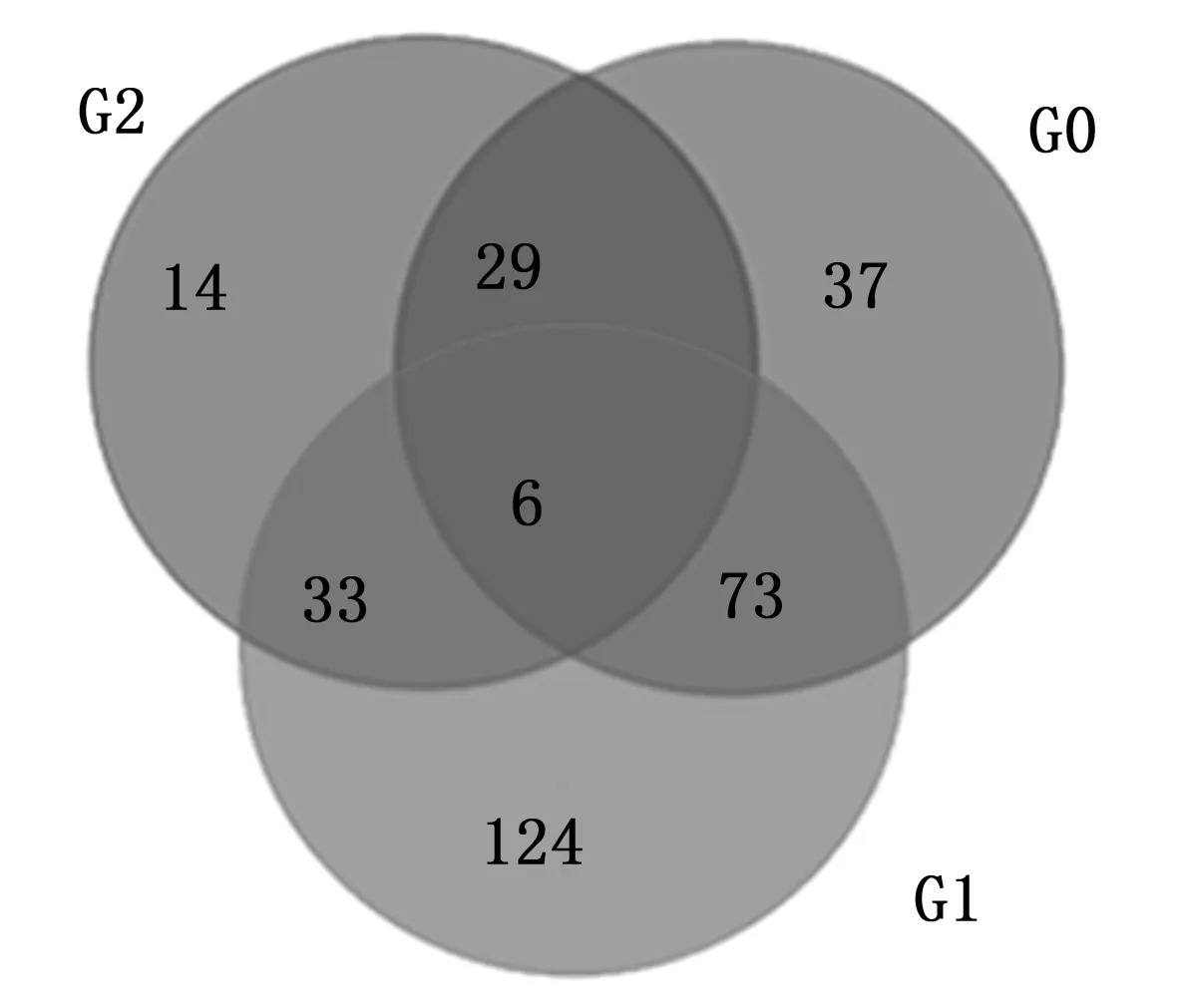
图3 不同处理下白桦差异表达基因维恩图
Fig.3 Venn diagram of differentially expressed genes of birch under different treatments



3 讨论
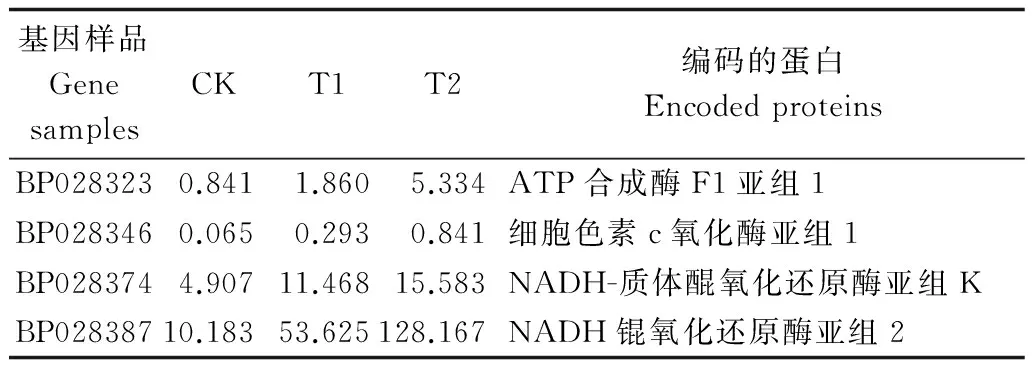
4 结论
* 通讯作者.
猜你喜欢
数学大王·中高年级(2021年3期)2021-03-10 11:19:30安徽农学通报(2017年1期)2017-02-15 18:28:07现代养生·下半月(2016年5期)2017-01-09 12:10:15安徽农学通报(2016年20期)2016-12-26 21:23:33现代农业科技(2016年20期)2016-12-20 14:47:52热带农业科学(2016年10期)2016-12-12 01:50:05中国科技博览(2016年15期)2016-08-23 00:39:42食品界(2016年4期)2016-02-27 07:37:06作文与考试·小学高年级版(2015年3期)2015-05-30 10:48:04作文与考试·小学高年级版(2015年5期)2015-05-30 10:48:04
