
暖温带森林土壤酶活性对增温的响应及其环境解析
2017-02-23 07:43:36刘彦春刘世荣
林业科学研究 2017年1期
王 一,刘彦春 ,刘世荣
(中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室,北京 100091)
暖温带森林土壤酶活性对增温的响应及其环境解析
王 一,刘彦春 ,刘世荣*
(中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室,北京 100091)
[目的]模拟气候变化背景下土壤酶活性季节变化规律和土壤酶活性对环境因子变异的响应,以深入理解气候变化背景下微生物因素在调控土壤碳循环过程中的作用。[方法]以暖温带锐齿栎林土壤为研究对象,采用红外辐射增温技术和96孔-微平板法测定了生长季和非生长季土壤水解酶和氧化酶活性,解析环境因子在调控土壤酶活性季节变异中的作用。[结果]显示:增温处理在生长季显著提高土壤温度1.91℃,导致土壤有机碳含量显著降低了12.15%,微生物量碳氮显著增加了40.30%和61.29%;增温处理在非生长季显著提高土壤温度2.24℃,但对微生物量碳氮影响并不显著。增温处理对土壤专一酶活性的影响存在季节差异,生长季除N-乙酰-葡糖苷酶外增温处理降低了土壤专一酶活性;非生长季除β-葡糖苷酶和蛋白酶外增温处理增加了土壤专一酶活性。环境因子可以解释土壤专一酶活性变异的82.5%,MBN和NH4-N极显著影响土壤专一酶活性,二者共可以解释变异的59.0%。[结论]MBN和NH4-N是影响土壤专一酶活性的主要环境因子,其他环境因子:土壤水热配比、微生物量碳、微生物量碳氮比、溶解性有机碳和硝态氮等是影响土壤专一酶活性在处理间和季节间变异的主要驱动因子。
暖温带;锐齿栎林;土壤酶活性;增温;环境解析

河南省宝天曼国家级自然保护区位于北亚热带向南暖温带过渡区域,属气候变化敏感区。Luan等人在此研究表明:温度是影响土壤呼吸的重要环境因子,土壤异氧呼吸的温度敏感性高于自养呼吸[17],说明该区域微生物对气候变暖的响应更敏感,预测未来气候变暖背景下该区微生物分解作用导致土壤碳库更容易分解。通过海拔土柱置换试验模拟气候变暖发现气候变暖显著增加了土壤CO2通量并改变土壤有机质层的分解过程[18]。本研究以该区域落叶阔叶林土壤为研究对象,采用红外辐射增温模拟气候变暖进行进一步研究。旨在探明:(1)模拟气候变化背景下,土壤酶活性季节变化规律;(2)土壤酶活性对环境因子变异的响应,以深入理解气候变化背景下微生物因素在调控土壤碳循环过程中的作用。
1 研究区域
2 研究方法
2.1 样地设置
采用随机区组试验设计,于2011年3月在研究区域典型林分中布设3个区组,具体试验设计见文献[20]。简言之:在增温样地内,将长165 cm,宽15 cm的红外线辐射加热灯(MSR-2420,Kalglo Electronics Inc., Bethlehem, PA, USA)平行样地短边垂直悬挂于样地正中央上方,悬挂高度2.25 m,额定功率为2 000瓦,全年持续供电;在对照样地内,以相同的方式安装与红外线辐射加热灯形状和大小相同的假灯,以消除红外线辐射加热灯遮阴对试验造成的误差。
2.2 土壤样品采集及土壤温湿度测定
2.3 土壤酶活性及理化性质测定
土壤酶测定:把相当于1 g干土的鲜土或1.25 g鲜土加入125 mL的醋酸钠缓冲液(50 mmol·L-1,pH=5.0)中,在搅拌机中处理1 min制成均质土壤悬液。水解酶测定采用荧光微平板法[21]:样品控制(800 μL土壤悬液+200 μL酶底物),对照控制(800 μL土壤悬液+200 μL缓冲液),淬火控制(800 μL土壤悬液+200 μL标准液),底物控制(控制1∶200 μL缓冲液+50 μL酶底物,控制2∶200 μL缓冲液+50 μL标准底物),以上操作除底物控制加入96孔板(黑板)外,其余均加入96深孔板中;在25℃培养箱中暗培养3 h后加入10 μL NaOH(0.5 mol·L-1)停止反应,将96深孔板以5 000 r·min-1离心3 min后,用八通道微量取样器取250 μL加到96孔板中(黑板),标仪(PerkinElmer Enspire,USA)进行荧光扫描(波长:365 nm,450 nm)。氧化酶测定采用比色法:以25 mmol·L-1L-DOPA作为底物,酚氧化酶测定中样品控制(800 μL土壤悬液+200 μLDOPA),对照控制(800 μL土壤悬液+200μL缓冲液),底物控制(200 μL缓冲液+50 μLDOPA);过氧化物酶测定中样品控制(800 μL土壤悬液+200 μL DOPA+40 μLH2O2),对照控制(800 μL土壤悬液+200 μL缓冲液+40 μL H2O2),底物控制(200 μL缓冲液+50 μL DOPA+10 μL H2O2)(H2O2浓度3%)。以上操作除底物控制加入96孔板(黑板)外,其余均加入96深孔板中。室温震荡暗培养1 h后,将96深孔板以5 000 r·min-1离心3 min后,用八通道微量取样器取250 μL加到96孔板中(黑板),酶标仪(PerkinElmer Enspire,USA)进行吸光扫描(波长:465 nm)。(底物信息见表1,酶活性计算方法见文献[22])

表1 底物基本信息Table 1 The substrate basic information
土壤理化性质测定:土壤有机碳含量采用重铬酸钾外加热方法测定[23],土壤全氮含量采用凯氏定氮方法测定[24];土壤铵态氮、硝态氮含量通过2 mol KCL溶液提取过滤后采用流动分析仪测定[25];土壤pH采用玻璃电极法测定[26];微生物量碳氮采用氯仿熏蒸浸提法[27]。
2.4 数据统计分析
文章中所有酶活性均采用土壤专一酶活性,专一酶活性=土壤酶活性/微生物量碳[28]。采用水热配比(S/T)作为衡量土壤环境因子的指标(S/T=平均土壤湿度/平均土壤温度)。采用t检验比较增温处理和对照处理土壤理化性质、生长季和非生长季内增温处理和对照处理土壤活性碳库、氮库和专一酶活性以及相同处理下生长季和非生长季专一酶活性的差异。所有的数据都使用SPSS17.0进行统计分析,图表采用Sigmaplot12.5绘制,用Canoco4.5分析专一土壤酶活性与环境因子间的关系。
3 结果与分析
3.1 样地土壤基本理化性质
表2可知:与对照样地相比,增温处理土壤有机碳含量显著低于对照处理 (P<0.05),土壤有机碳含量降低12.15%;增温处理土壤总氮和pH值虽低于对照处理,但未达到显著水平;增温与对照处理对土壤碳氮比的影响也未达到显著水平。
3.2 增温处理对土壤环境和活性碳库与氮库的影响
表3和4表明:生长季和非生长季中,增温处理显著改变土壤环境,增温处理显著增加土壤温度和降低土壤湿度导致土壤水热配比显著低于对照样地,且增温处理导致非生长季水热配比下降幅度大于生长季的下降幅度;土壤溶解性有机碳含量在生长季和非生长季对增温处理的响应均未达到显著水平;生长季增温处理土壤微生物量碳和微生物量氮显著高于对照样地,分别增加了40.30%和61.29%,微生物量碳氮比显著低于对照样地12.64%;非生长季增温处理对土壤微生物量碳、微生物量氮和微生物量碳氮比的影响并不显著;土壤硝态氮和铵态氮含量在生长季和非生长季对增温处理的响应均未达到显著水平。
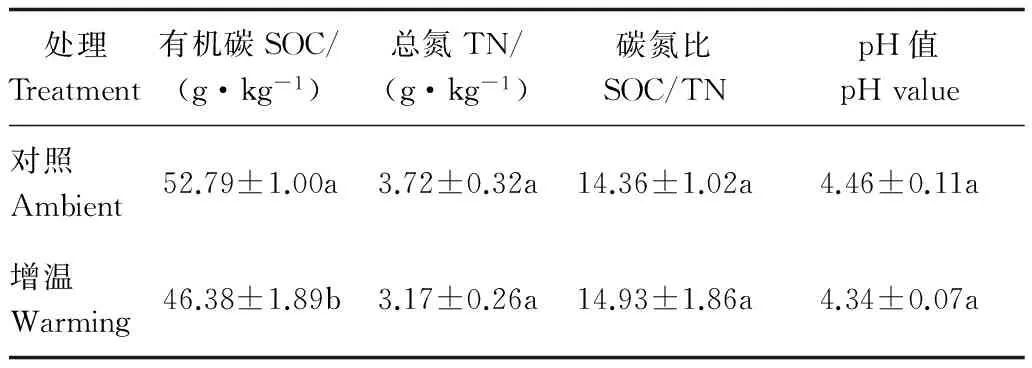
表2 样地土壤基本理化性质Table 2 Soil physical and chemical properties
(不同字母表示在0.05水平上显著差异)
(The different letters refer to significant effects atp<0.05)
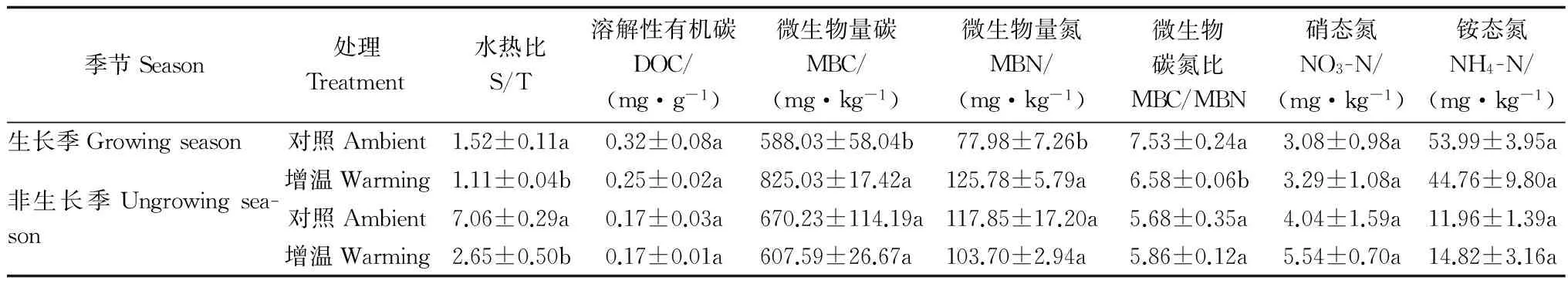
表3 土壤环境及土壤活性碳库和氮库的t检验结果Table 3 Results of t-test on soil labile carbon and nitrogen pools and environment
(不同字母表示在0.05水平上显著差异)
(The different letters refer to significant effects atp<0.05)

表4 增温和对照处理土壤温湿度的t检验结果Table 4 Results of t-test on soil moisture and temperature
(不同字母表示在0.05水平上显著差异)
(The different letters refer to significant effects atp<0.05)
3.3 增温处理对土壤酶活性的影响
增温处理改变了土壤专一水解酶和氧化酶活性(图1)。除NAG外,生长季增温处理降低了AG、BG、CB和LAP专一酶活性,但均未达到显著水平;增温处理显著降低了PER活性,未显著改变POX活性。非生长季增温处理增加了AG、CB、NAG、PER和POX专一酶活性,但降低了BG和LAP专一酶活性,且对LAP影响达到显著水平;与生长季不同,增温处理在非生长季不同程度地增加土壤专一酶活性,但未达到显著水平。图1和表5可知:除AG和LAP外,对照处理非生长季酶活性均低于生长季;增温处理非生长季酶活性均高于生长季,且非生长季AG活性显著高于生长季活性(P=0.019)。增温处理在生长季普遍降低了土壤专一酶活性,但非生长季增温处理则增加了土壤专一酶活性。说明土壤酶活性存在季节间变异,且增温处理在不同季节间对土壤酶的影响不同。
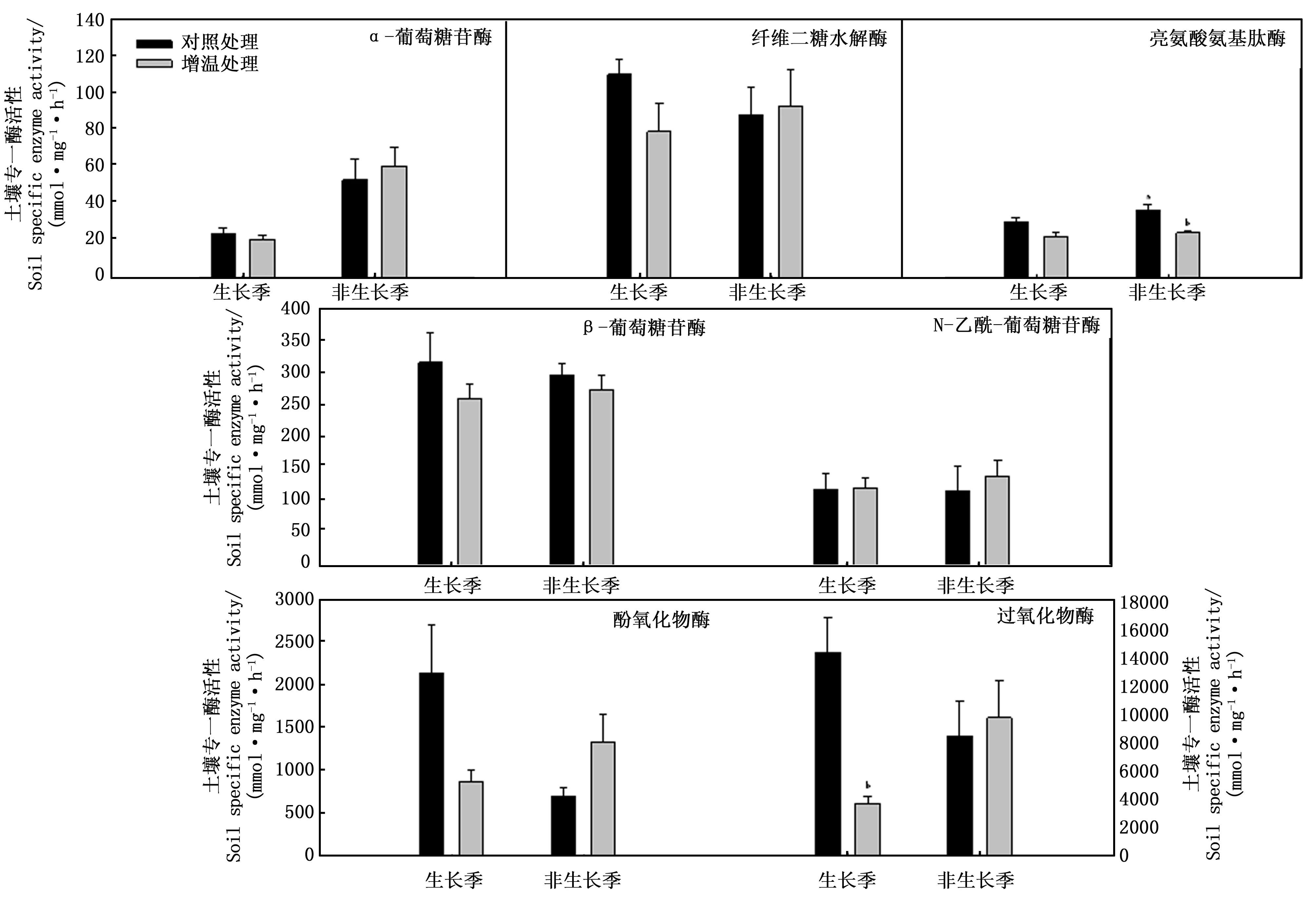
图1 生长季和非生长季土壤专一酶活性t检验结果Fig.1 Results of t-test on soil specific enzyme activity in growing and ungrowing seasons
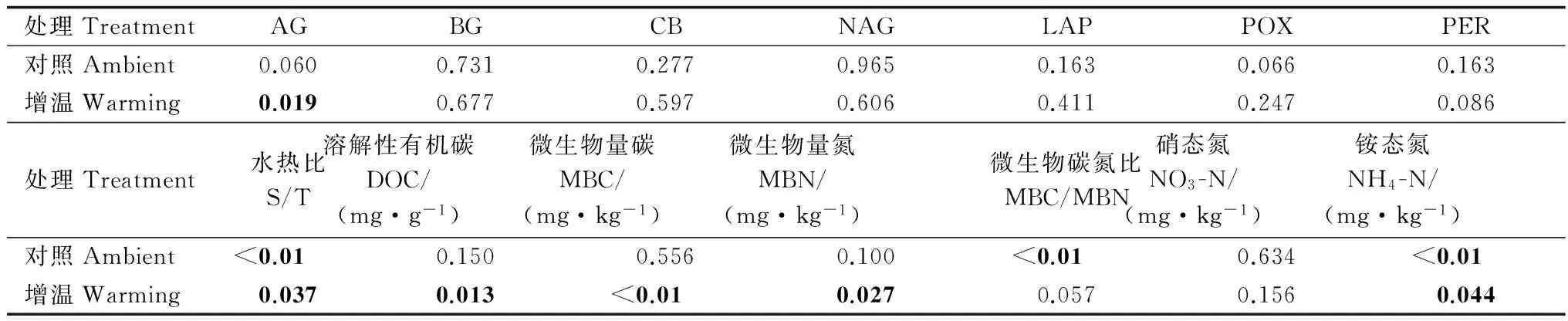
处理TreatmentAGBGCBNAGLAPPOXPER对照Ambient0.0600.7310.2770.9650.1630.0660.163增温Warming0.0190.6770.5970.6060.4110.2470.086处理Treatment水热比S/T溶解性有机碳DOC/(mg·g-1)微生物量碳MBC/(mg·kg-1)微生物量氮MBN/(mg·kg-1)微生物碳氮比MBC/MBN硝态氮NO3-N/(mg·kg-1)铵态氮NH4-N/(mg·kg-1)对照Ambient<0.010.1500.5560.100<0.010.634<0.01增温Warming0.0370.013<0.010.0270.0570.1560.044
(加粗字体表示在0.05水平上显著差异)
(The bold numbers refer to significant effects atp<0.05)
3.4 土壤酶活性的环境解析
冗余度分析(RDA)表明:9个环境因子变量中,有2个环境因子(NH4-N和MBN)与专一土壤酶活性显著相关。RDA分析中第一主成分和第二主成分共可以解释9个环境因子与7种土壤专一酶活性关系变异的82.5%(图2)。NH4-N和MBN共可以解释土壤专一酶活性变异的59.0%。RDA排序图说明主要环境因子和土壤专一酶活性之间的特异关系。其中LAP和AG与S/T和NO3-N呈正相关关系,POX与NH4-N、DOC和MBC/MBN呈正相关关系,NAG、PER、BG和CB则与DOC和MBC/MBN呈正相关关系。说明土壤专一酶活性变异受多种环境因子同时调控。
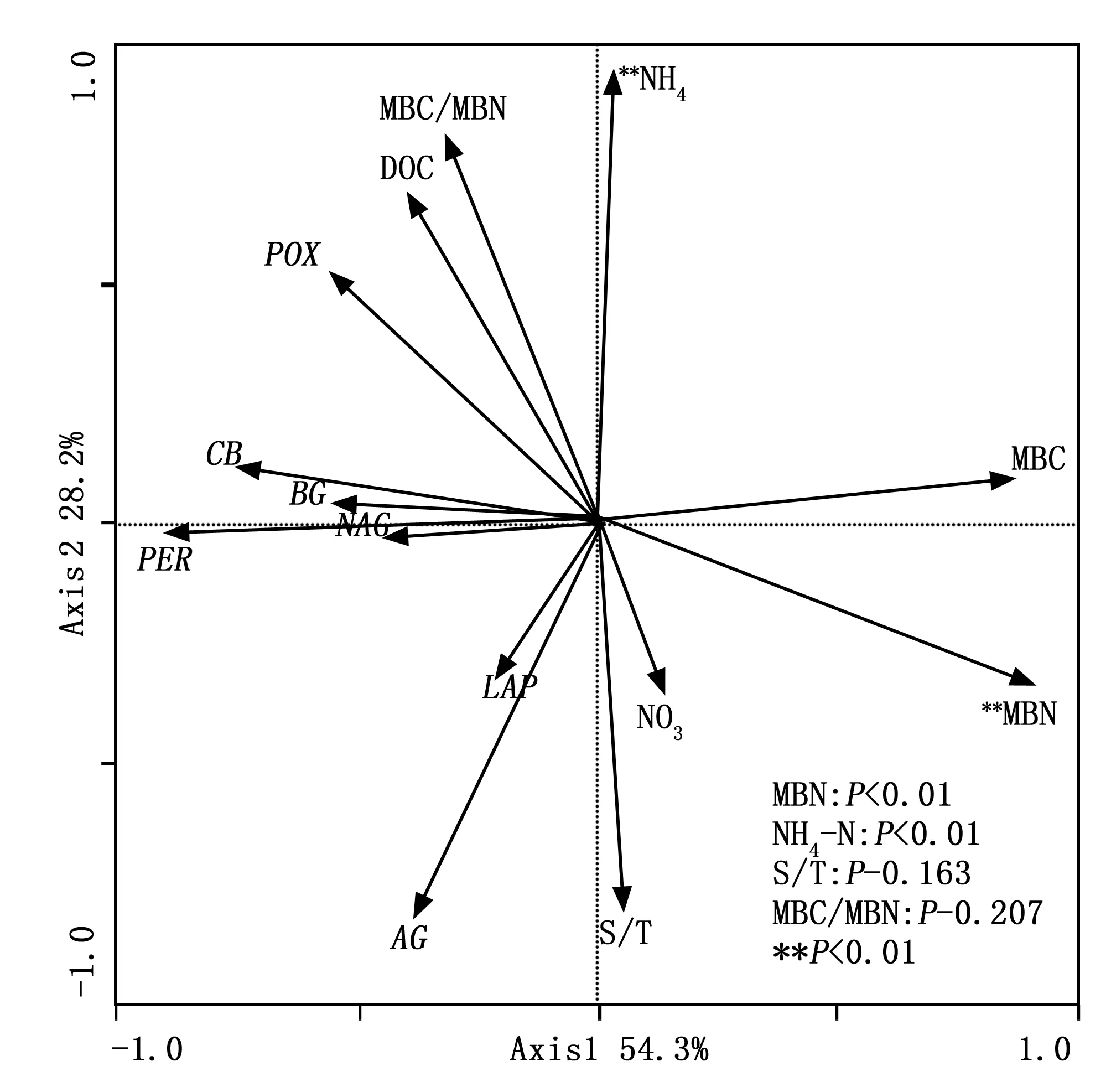
图2 环境因子与土壤专一酶活性RDA分析Fig.2 Redundancy analysis of soil specific enzyme activity and environmental factors
4 讨论
4.1 增温处理对土壤碳库和氮库的影响
作为土壤碳库和氮库中重要组成部分,活性碳库和氮库(微生物量碳、微生物量氮、溶解性有机碳和溶解性有机氮)虽然所占比例不高,却在调节土壤碳氮循环中发挥着重要的作用。研究表明:生长季增温处理显著提高了土壤中微生物量碳氮,但非生长季增温处理对微生物量碳氮的影响并不显著,说明微生物量碳氮对增温处理的响应受季节变化影响。Xu等人[32]开展增温试验也发现土壤活性碳库和氮库对增温处理的响应受季节变化影响显著。溶解性有机碳作为土壤微生物有机体可直接利用碳源,增温处理并未显著增加溶解性有机碳含量。Luo等人[33]研究表明,温度和湿度并不是影响土壤溶解性有机碳的主要因子,地下生物量等生物因子是调控土壤溶解性有机碳的重要因子。故增温处理虽显著增加土壤温度,但并不能增加土壤中溶解性有机碳的含量。微生物量碳氮比在粗略水平上可以看作微生物中细菌和真菌的相对丰度比[34]。生长季中增温处理显著降低了土壤微生物碳氮比,说明与对照样地相比增温样地的细菌群落占优势。已有整合分析研究表明[35]:增温处理会显著增加土壤净氮矿化作用和净硝化作用。本研究观察到增温处理虽导致土壤硝态氮和铵态氮(生长季除外)含量增加,但并未达到显著水平。这可能是因为增温处理导致细根生物量和土壤微生物量的增加促进了二者对土壤中无机氮的吸收和利用。
4.2 增温处理对土壤专一酶活性的影响
土壤酶作为土壤微生物活性和土壤理化性质的指示因子[11],在调控土壤碳氮循环中发挥着重要的作用。由于土壤酶活性与土壤性质、微生物特性等密切相关,所以增温处理下土壤酶活性可能出现季节性变化。在考察土壤酶活性的研究中,专一酶活性可以把微生物和土壤酶活性建立较好的联系。图1表明:生长季中除NAG外,增温处理不同程度降低其他4种水解酶专一酶活性,但均未达到显著水平。说明气候变暖情况下,酶催化反应变得更有效,单位微生物产生的酶量降低。黄雪蔓[36]研究发现:土壤微生物群落受土壤底物的数量(有机碳含量)和质量(碳氮比)影响。作为土壤微生物两大主要类群,通常认为细菌与水解酶活性相关,主要利用土壤中活性底物;真菌与氧化酶活性相关,主要利用土壤中惰性底物[37-38]。微生物量碳氮比作为粗略反映微生物群落中细菌和真菌相对丰度的指标,且真菌比细菌具有更高的碳氮比,所以微生物量碳氮比升高反映真菌相对丰富度更高。生长季中对照样地微生物量碳氮比(7.53)显著高于增温样地微生物碳氮比(6.58),说明对照样地中真菌相对丰富度比增温样地的高,真菌通常分泌氧化酶以分解大分子化合物,故生长季对照样地内过氧化物酶活性显著高于增温样地的。
环境因子变化可影响土壤底物扩散作用[39]和底物利用有效性[40]。非生长季中土壤温度因子成为限制性因子时,底物扩散作用和有效性受到限制,微生物利用底物效率和微生物活性也随之降低。增温处理显著提高了非生长季土壤温度,故非生长季增温样地微生物活性增强,微生物为满足自身生长需要,单位微生物量会产生更多的酶以提高其对土壤养分的利用,所以增温处理在非生长季对土壤专一酶活性多表现出促进作用。
4.3 环境因子变异对专一土壤酶活性的影响
除温度因子外,土壤湿度也影响土壤底物的扩散作用[39],作者引入水热比(S/T),考察温湿度因子共同作用对土壤专一酶活性的影响。图2可知:LAP和AG与S/T和NO3-N显著正相关,而温度因子对氮循环酶LAP的作用不明显[41],说明LAP活性受土壤湿度影响,该结论与Brzostek通过整合分析发现LAP酶活性受土壤湿度限制的结论一致[3];表5可知:增温样地和对照样地非生长季土壤NO3-N与生长季土壤NO3-N差异并不显著,但非生长季S/T显著高于生长季,故非生长季LAP和AG活性与生长季相比均表现出增加趋势(图1,表5)。RDA排序图表明NAG、BG和CB则与DOC和MBC/MBN呈正相关关系。表5可知:非生长季与生长季相比,对照样地MBC/MBN显著降低是引起非生长季对照样地酶(NAG、BG和CB)活性低于生长季的主要原因,说明微生物群落结构的变化可能是影响土壤专一酶活性季节变化的主要原因。You[42]在该区域研究也发现微生物群落结构与土壤酶活性之间具有很强的相关性,氧化酶活性与真菌相关而细菌与水解酶活性相关。这与我们得到的结论较为一致。图2可知,POX与NO3-N呈负相关关系;PER与NO3-N和NH4-N也呈负相关,与Sinsabaugh等人[37]研究结果一致,认为氧化酶活性与土壤无机氮可利用性呈负相关关系。我们把MBC作为分子将总土壤酶活性专一化,故RDA分析中土壤专一酶活性与MBC和MBN呈负相关关系。
5 结论
(1)增温处理显著提高了土壤温度,但并未显著降低土壤湿度。
(2)生长季增温处理引起土壤温度增加改变了酶催化反应效率进而改变了土壤专一酶活性;除PER外,增温处理导致土壤专一酶活性的降低并未达到显著水平。非生长季增温处理普遍增加了土壤专一酶活性,说明土壤专一酶活性对增温处理的季节性响应不同。
(3)除微生物量氮和铵态氮外,其他环境因子,诸如:土壤水热比、微生物量碳、微生物量碳氮比、溶解性有机碳和硝态氮等是影响土壤专一酶活性在不同处理间和季节间变异的主要原因。
[1] IPCC. Climate Change 2013: The Physical Science Basis[M]. Cambridge University Press, Cambridge, UK., 2013.
[2] Groffman P M, Driscoll C T, Fahey T J,etal. Effects of mild winter freezing on soil nitrogen and carbon dynamics in a northern hardwood forest.[J]. Biogeochemistry, 2001, 56(2): 191-213.
[3] Brzostek E R, Blair J M, Dukes J S,etal. The effect of experimental warming and precipitation change on proteolytic enzyme activity: positive feedbacks to nitrogen availability are not universal[J]. Global Change Biology, 2012, 18(8): 2617-2625.
[4] Baldrian P,najdr J, Merhautová V,etal. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change[J]. Soil Biology and Biochemistry, 2013, 56: 60-68.
[5] Weedon J T, George A Kowalchuk, Bodegom P M. No effects of experimental warming but contrasting seasonal patterns for soil peptidase and glycosidase enzymes in a sub-arctic peat bog[J]. Biogeochemistry, 2014, 117(1):55-66.
[6] Bastida F, Barberá G G, García C,etal. Influence of orientation, vegetation and season on soil microbial and biochemical characteristics under semiarid conditions[J]. Applied Soil Ecology, 2008, 38(1): 62-70.
[7] Wittmann C, Kähkönen M A, Llvesniemi H,etal. Areal activities and stratification of hydrolytic enzymes involved in the biochemical cycles of carbon, nitrogen, sulphur and phosphorus in podsolized boreal forest soils[J]. Soil Biology and Biochemistry, 2004, 36(3): 425-433.
[8] Burns R. Enzyme activity in soil: location and a possible role in microbial ecology[J]. Soil Biology and Biochemistry, 1982, 14(5): 423-427.
[9] Sowerby A, Emmett B, Beier C,etal. Microbial community changes in heathland soil communities along a geographical gradient: interaction with climate change manipulations[J]. Soil Biology and Biochemistry, 2005, 37(10): 1805-1813.
[10] Zogg G P, Zak D R, Ringelberg D B,etal. Compositional and functional shifts in microbial communities due to soil warming[J]. Soil Science Society of America Journal, 1997, 61(2): 475-481.
[11] Weedon J, Aerts R., George A. Kowalchuk,etal. Enzymology under global change: organic nitrogen turnover in alpine and sub-Arctic soils[J]. Biochemical Society Transactions, 2011, 39(1): 309.
[12] Wang X, Dong S, Gao Q,etal. Effects of short-term and long-term warming on soil nutrients,microbial biomass and enzyme activities in an alpine meadow on the Qinghai-Tibet Plateau of China[J]. Soil Biology and Biochemistry, 2014, 76:140-142.
[13] Zhou X, Chen C, Wang Y,etal. Warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland[J]. Science of the Total Environment, 2013, 444 (1):552-558.
[14] Steinweg J M, Dukes J S, Paul E A,etal. Microbial responses to mutli-factor climate change: effects on soil enzymes[J]. Frontiers in Microbiology, 2013,4(146):1-11.
[15] McDaniel M D, Kaye J P, Kaye M W. Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest forest[J]. Soil Biology and Biochemistry, 2013, 56:90-98.
[16] Bengtson P, Bengtsson G. Rapid turnover of DOC in temperate forests accounts for increased CO2production at elevated temperatures[J]. Ecology Letters, 2007, 10(9): 783-790.
[17] Luan J, Liu S, Wang J,etal. Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China[J]. Soil Biology and Biochemistry, 2011, 43(3): 503-512.
[19] 史作民, 程瑞梅, 刘世荣, 等. 河南宝天曼化香林特征及物种多样性[J].山地学报, 2005, 23(3): 374-380.
[20] 刘彦春, 暖温带锐齿栎林土壤呼吸及微生物群落结构对土壤增温和降雨减少的响应[D].北京: 中国林业科学研究院. 2013.
[21] Saiya-Cork K R, Sinsabaugh R L. The effects of long term nitrogen deposition on extracellular enzyme activity in anAcersaccharumforest soil[J]. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315.
[22] German D P, Weintraub M N, Grandy A S,etal. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies[J]. Soil Biology and Biochemistry, 2011, 43(7): 1387-1397.
[23] Nelson D W, Sommers L E, Sparks D L,etal. Total Carbon, Organic Carbon, and Organic Matter[M]//Methods of Soil Analysis.Part 2. Chemical and Microbiological Properties, 1982: 539-579.
[24] Bremner J M, Mulvaney C, Nitrogen-Total[M]//Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 1982: 595-624.
[25] Magill A H, Aber J D. Variation in soil net mineralization rates with dissolved organic carbon additions[J]. Soil Biology and Biochemistry, 2000, 32(5): 597-601.
[26] LY/T1228-999. 森林土壤 pH 值的测定[S]. 1999.
[27] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C[J]. Soil Biology and Biochemistry, 1987, 19(6): 703-707.
[28] Stone M M, DeForest J L, Plante A F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory[J]. Soil Biology and Biochemistry, 2014,75: 237-247.
[29] Post W M, Emanuel W R, Zinke P J,etal. Soil carbon pools and world life zones[J]. Nature,1982, 298:156-159.
[30] Shaver G R, Billings W D, Chapin F S,etal. Global change and the carbon balance of arctic ecosystems[J]. BioScience, 1992,42(6): 433-441.
[31] Rui Y, Wang S, Xu Z,etal. Warming and grazing affect soil labile carbon and nitrogen pools differently in an alpine meadow of the Qinghai-Tibet Plateau in China[J]. Journal of Soils and Sediments, 2011, 11(6): 903-914.
[32] Xu Z, Hu R, Xiong P,etal. Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China: nutrient availabilities, microbial properties and enzyme activities[J]. Applied Soil Ecology, 2010, 46(2): 291-299.
[33] Luo C, Xu G, Wang Y,etal. Effects of grazing and experimental warming on DOC concentrations in the soil solution on the Qinghai-Tibet plateau[J]. Soil Biology and Biochemistry, 2009, 41(12): 2493-2500.
[34] Yao H, Bowman D, Shi W. Seasonal variations of soil microbial biomass and activity in warm-and cool-season turfgrass systems[J]. Soil Biology and Biochemistry, 2011,43(7): 1536-1543.
[35] Bai E, Li S, Xu W,etal. A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics[J]. New Phytologist, 2013, 199(2): 441-451.
[36] 黄雪蔓, 刘世荣, 尤业明. 固氮树种对第二代桉树人工林土壤微生物生物量和结构的影响[J].林业科学研究,2014,27(5):612-620.
[37] Sinsabaugh R L, Gallo M E, Lauber C,etal. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition[J]. Biogeochemistry, 2005, 75(2): 201-215.
[38] Talbot J M, Bruns T D, Smith D P,etal. Independent roles of ectomycorrhizal and saprotrophic communities in soil organic matter decomposition[J]. Soil Biology and Biochemistry, 2013, 57: 282-291.
[39] Koch A L. Diffusion the crucial process in many aspects of the biology of bacteria, in Advances in microbial ecology[M]. 1990.Springer. 37-70.
[40] Koch O, Tscherko D, Kandeler E. Temperature sensitivity of microbial respiration, nitrogen mineralization, and potential soil enzyme activities in organic alpine soils[J]. Global Biogeochemical Cycles, 2007, 21(4): 497-507.
[41] Wallenstein M D, Mcmahon S K, Schimel J P. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils[J]. Global Change Biology, 2009, 15(7): 1631-1639.
[42] You Y, Wang J, Huang X,etal. Relating microbial community structure to functioning in forest soil organic carbon transformation and turnover[J]. Ecology and Evolution, 2014, 4(5): 633-647.
(责任编辑:彭南轩)
Response of Soil Enzyme Activities to Soil Warming and Explanation of Environmental Factors in Warm-temperate Oak Forest
WANGYi,LIUYan-chun,LIUShi-rong
(Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Key Laboratory of Forest Ecology and Environment, State Forestry Administration, Beijing 100091, China)
[Objective]To study the seasonal variation of soil enzyme activities and its response to environmental factors and to understand the effect of microbial factor in the regulation of soil carbon cycle in the context of climate change. [Method]The infrared radiation heater and the 96 micro-plate method were used to assay the activity of the hydrolytic and oxidative enzymes of the soil, and determine the role of environmental factors in addressing the seasonal variation in the activity of soil enzymes. [Result]During growing season, the warming treatment significantly increased the soil temperature by 1.91℃, leading to a 12.15% decrease of organic carbon content and significant increase of 40.30% and 61.29% of microbial biomass carbon (MBC) and nitrogen (MBN). During non-growing season, however, although the warming treatment increased the soil temperature by 2.24℃, no significant effect was found on MBC or MBN. The effects of warming treatment on specific enzyme varied in different seasons. During growing season, the activities of specific enzyme in warming treatment were lower than those in control except for NAG, while they were higher during non-growing season except for BG and LAP. [Conclusion]The environmental factors accounted for 82.5% of the total variations of soil enzyme activities, while the activities of specific enzyme were significantly affected by NH4-N and MBN. The other environmental factors (soil moisture, MBC, DOC, MBC/MBN and NO3-N) were the main reasons for the variations of specific enzyme activities in different treatments and seasons.
warm-temperate zone;Quercusalienavar.acuteserrata; soil enzyme activity; warming; explanation of environmental factors
10.13275/j.cnki.lykxyj.2017.01.016
2015-04-30
2016-02-20
国家自然科学基金重大项目“植物对森林生态系统碳-氮-水耦合循环的作用机制”(项目编号:31290223);中国林业科学研究院基本科研业务费专项资金项目“海内外杰出学者讲习计划”(项目编号:CAFYBB2011007)。
王 一(1986—),在读博士,主要研究方向为模拟气候变化背景下森林生态系统土壤碳氮循环。E-mail:wylx0830@126.com
* 通讯作者:E-mail: liusr9311@163.com
S714
A
1001-1498(2017)01-0117-08
猜你喜欢
土壤学报(2022年3期)2022-08-26 12:12:18
露天采矿技术(2020年6期)2021-01-05 00:37:54
环境卫生工程(2020年3期)2020-07-27 01:19:22
辽宁林业科技(2017年4期)2017-06-22 10:23:36
环境科技(2016年2期)2016-11-08 12:18:20
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
西南农业学报(2016年5期)2016-05-17 05:42:39
大气科学(2015年5期)2015-12-04 03:04:44
化工进展(2015年6期)2015-11-13 00:32:09
植物营养与肥料学报(2011年2期)2011-10-26 03:52:20
