
塔里木盆地鱼类入侵及区系演变趋势
2017-02-08 05:58陈国柱仇玉萍李丽萍
生态学报 2017年2期
陈国柱, 仇玉萍, 李丽萍
西南林业大学, 国家高原湿地研究中心, 昆明 650224
塔里木盆地鱼类入侵及区系演变趋势
陈国柱*, 仇玉萍, 李丽萍
西南林业大学, 国家高原湿地研究中心, 昆明 650224
鱼类入侵是塔里木盆地土著鱼类濒危的关键性影响因素之一,结合历史文献数据及近年调查数据,系统综述了塔里木盆地鱼类入侵现状,并就鱼类入侵引起的鱼类区系均一化问题进行了分析。结果显示,该地区记录鱼类63种,其中土著种19种,入侵种44种。尽管入侵鱼类提升了塔里木盆地鱼类群落属级及科级的多样性水平,但相应的G-F多样性指数却明显下降,表明该区鱼类群落从物种组成简单、分化明显的特点演变为物种成分复杂、分化贫乏的特点。入侵种中源于长江和额尔齐斯河分别有30种(68.2%)和7种(15.9%),源于美欧和亚洲其他区域的有7种(15.9%)。入侵鱼类的进入引起塔里木盆地与其相关区域鱼类区系相似性显著上升(P<0.01),并导致该区鱼类区系从单一的中亚高原山区复合体演变为多区系混合体。进一步利用鱼类个体生态矩阵分析方法研究显示,外来种在塔里木盆地适应性广泛,能够占据水域各种栖息生境,显著压缩了土著种的栖息空间,进而通过捕食、食物竞争及其他作用途径严重危及土著鱼类种群的续存,提示须采取有力措施限制入侵种的进一步扩散及对濒危土著物种迅速开展人工保育工作。
塔里木盆地;生物入侵;鱼类;物种濒危
鱼类是全球生物多样性的关键组分,更是多数水域生态系统的核心功能成分,近年来,鱼类入侵引起的生态问题引起了社会及学术界的广泛关注[1-5]。我国青藏、云贵及蒙新高原区域在历史上演化出众多独特而脆弱的珍稀鱼类物种,是我国生物多样性不可或缺的成员,然而近代以来在外来鱼类引种潮所引发的入侵种问题影响下这些原生物种许多已然处于灭绝或濒危边缘[6-8]。系统性研究入侵鱼类对我国高原区域水域生态系统的影响,特别是对它们对土著鱼类的危害及作用机制进行深入探索,进而采取有力措施限制其进一步扩散,从而切实保育现存的珍稀土著鱼类资源是当前我国生物多样性保育工作中一项极具挑战性的任务。迄今为止,国内尚没针对外来鱼类入侵引起区域性大尺度鱼类物种多样性及鱼类区系区域变化等基础性工作进行过专门研究,本文将首先对外来鱼类入侵引起的我国典型内流河区域塔里木盆地鱼类物种多样性及鱼类区系区域剧变问题展开初步研究。
塔里木盆地位于我国新疆维吾尔自治区南部,据欧亚大陆的中央,远离海洋,面积约有91万km2,平均海拔约1000 m,周围被天山、阿尔金山、昆仑山等大山脉所包围。盆地内气候极为干旱,仅能依靠周围高山冰雪融水形成有限的地表径流,这些径流进入盆地后大部分汇集入我国最大的内流河塔里木河(干流约1321 km,总长约2350 km,支流众多,海拔约650—4150 m),而最终所有地表径流均消失在盆地中央的塔克拉玛干沙漠[9-11](图1)。特殊的自然地理环境形成了该盆地有别于我国其他地区的独特鱼类区系(为塔里木亚区),物种组成简单,多样性低而独特性高,主要由裂腹鱼类及高原鳅类所组成,特有种有扁吻鱼(Aspiorhynchuslaticeps)、塔里木裂腹鱼(Schizothorax(Racoma)biddulphi)、隆额高原鳅(Triplophysabombifrons)和叶尔羌高原鳅(Triplophysayarkandensis)等4种(亚种)[6, 12],物种分化十分显著[12-13]。在经历了20世纪60年代以来运动式引种后,塔里木盆地出现了大量鱼类入侵物种,这些入侵物种已经成为导致土著鱼类濒危的关键性影响因素之一[6,14-16]。由于塔里木盆地的封闭性,鱼类入侵引发的生态学问题具有独特性及典型性,该区是研究我国鱼类入侵问题较为理想区域之一。
王德忠[6]最早报道了塔里木河流域外来鱼类问题,他指出由于为外来鱼类的引入,塔里木河流域的鱼类由原来的15种和亚种增加到41种和亚种,其区系成份从只有中亚高山复合体一种成分,增加至5个复合体成分。尔后,马燕武等[15]将塔里木河流域鱼类的土著鱼类订为12种,而程勇等[16]统计鉴定了塔里木河干流上游及附属水体中已建群的入侵鱼类共18种。其他研究,如武云飞和吴翠珍[17]针对喀刺昆仑山-昆仑山等塔里木盆地南缘的鱼类物种组成进行了研究,列出了10种入侵鱼类,已经初步认识到外来鱼类对土著鱼类渔业的影响;郭焱等[14]列出了博斯腾湖中栖息的鱼类,30种鱼类绝大部分为入侵鱼类,原有的扁吻鱼等5种土著鱼类已经在湖体消失;马燕武等[18]对塔里木河支流开都河流域的调查显示,现存3种土著鱼类局限于分布在外来种尚未占据优势的上游区域,该流域入侵鱼类共计16种;马燕武等[19]对扁吻鱼现存的唯一的自然栖息地塔里木河另一支流木扎提河(克孜尔水库)的调查显示,该区共有9种入侵鱼类栖息,已占现存鱼类群落物种数的47.4%。王智超等[20]发现了新入侵种河川沙塘鳢(Odontobutispotamophila)在塔里木干流成功定殖。近年来,塔里木盆地外来鱼类物种呈现出逐渐增加、分布区域日益扩大、危害越发明显的趋势。
多年来,虽然研究者对塔里木盆地外来鱼类入侵问题多所涉猎,但尚未能对该区入侵鱼类问题进行过系统性研究。究竟目前该盆地有多少鱼类入侵物种,这些物种来源组成如何,造成了什么样关键性生态影响等基础问题人们对之尚缺乏深入了解,极大制约了该区对鱼类入侵物种治理及土著濒危鱼类物种保护实践的开展。
本文综合分析文献资料及结合实地调查资料,针对塔里木盆地入侵鱼类物种编目、多样性特征、来源组成、生态特征及鱼类区系演变等问题展开研究,力图初步揭示塔里木盆地鱼类入侵问题概况,为系统性研究塔里木盆地鱼类入侵生态学问题奠定基础。
1 材料与方法
1.1 材料
有关塔里木盆地鱼类物种记录及个体生态学资料主要收集自公开发表的论文、专业书籍,部分补充资料来自本单位自2010—2015年在塔里木盆地区域内5个国家湿地公园(试点)建设总体规划设计过程中对这些区域鱼类本底资源调查结果。
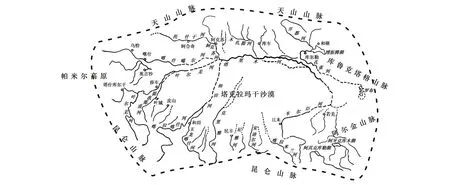
图1 塔里木盆地水系示意图[12]Fig.1 Drainage systems of Tarim Basin, adapted from reference[12]
1.2 研究方法
塔里木盆地鱼类物种组成及物种多样性变化。根据鱼类分布及物种组成资料分析综合编制塔里木盆地鱼类名录,在此基础上采用G-F指数方法来评估塔里木河段的鱼类多样性变化。
G-F指数计算方法[21-22]:
F指数(DF)(科的多样性)
式中,Pi=Ski/Sk,Ski为名录中k科i属中的物种数,Sk为名录中k科中的物种数,n为k科中的属数,m为名录中鱼类的科数。
G指数(DG)(属的多样性)
式中,qj=sj/S,sj为j属中的物种数,S为名录中鱼类的物种数,P为名录中鱼类的属数。
G-F指数计算(DG-F)
DG-F=1-DG/DF
式中,G-F指数是 0-1的测度,非单种的科越多,G-F指数越高,反之,该指数值就越低。G-F指数应用价值在于能够仅从属、种水平和单种科的多寡来反映某一地区较长一段时间的物种多样性,而不必考虑种群数量和均匀度[21]。
塔里木盆地鱼类群落与相关区域鱼类群落组成相似性分析。利用群落相似性指数(Jaccard′s相似性指数)分析塔里木盆地与伊犁河谷[23]、青藏高原区域的西藏[8,24-25]、青海[26-27]等、长江中下游主要湖泊鄱阳湖[28]、洞庭湖[29]、太湖[30]等鱼类群落的组成的相似性,揭示由鱼类入侵引起的鱼类区系均一化演变问题。
相似性指数I计算方法[31]
I=j/(a+d-j)
式中,a为群落A中的物种数,b为群落B中的物种数,j为群落A和B中的共有种数。当I为0.75—1.0时为极相似;为0.5—0.75时为中等相似;为0.25—0.5时为中等不相似;为0—0.25时,为极不相似。
塔里木盆地鱼类个体生态矩阵分析。鱼类个体生态需求矩阵(R)计算公式[32]:
R=(S×A)/ns
式中,S为鱼类群落物种名录矩阵,A为鱼类个体生态特征矩阵,ns为鱼类群落种类数,即以研究区域鱼类物种名录为纵列、该区域鱼类所具有的生态特征指标为横列,组成二联列表(矩阵),求取相应的鱼类环境需求指数R[R1,R2,R3,…,Ri],Ri表示某一样点中鱼类群落对环境某一具体因子的需要程度[32-33]。鱼类个体生态需求矩阵是将已经收集到的鱼类生态特征按目的进行整理、筛选,从而用来评价鱼类群落变化与环境关系的指标,是以0—1为量度,某种生态特征对应的Ri值越高,则说明具有此特征的鱼类受此相关环境因子变化的影响越大[32-33]。应用鱼类个体生态学矩阵分析方法时,鱼类个体生态学指数包括了群落中每种鱼类已知生态学特征如生活史、产卵条件、相应的水质要求等等[34],当某种鱼类某些生态特征不明确时,则它的该项特征在矩阵中相应缺如。它的优点及作用主要在于一方面可以避免由于随机抽样带来的数量差异影响,对鱼类功能群整体进行分析,尤其对鱼类资源历史调查资料信息能充分利用;而另一方面可通过分析鱼类个体生态需求矩阵计算出鱼类的环境需求,分析样点鱼类群落的环境需求,进而推断出研究目标区域的生态环境历史变化[33]。
2 结果
2.1 塔里木盆地鱼类群落演变
综合文献及实地调查资料分析显示,塔里木盆地原有土著鱼类共19种,外来鱼类44种,鱼类物种数共计63种(附表1),外来鱼类所占比例为69.8%。原有鱼类组成中仅有1目2科6属,为鲤形目,鲤科、鳅科及裸重唇鱼属、扁吻鱼属、裂腹鱼属、重唇鱼属、裸裂尻鱼属、高原鳅属等;现今鱼类组成有5目16科42属,数量显著上升。显示经历了半个多世纪外来鱼类的持续性引入,塔里木盆地鱼类群落中物种组成出现了显著变化。
原有土著鱼类群落F指数(DF)(科的多样性)为1.30,G指数(DG)(属的多样性)为0.74,G-F指数为0.43。当前鱼类群落总体F指数为4.21,G指数为3.38,G-F指数为0.20,群落中单种科的数目较原土著鱼类群落显著上升。
与此同时,塔里木盆地鱼类群落组成与周边相关区域的相似性也发生了明显变化(表1)。通过非参检验分析显示,相对历史时期,当今塔里木盆地鱼类群落与周边相关区域的相似性指数显著上升(Mann-Whitney U检验,P<0.01)。与最为接近的伊犁河流域鱼类群落对比分析,在外来鱼类被大规模被引入新疆前,两者的共有种仅有4种,分别为新疆裸重唇鱼(Gymnodiptychusdybowskii)、斑重唇鱼(Diptychusmaculatus)、新疆高原鳅(Triplophysastrauchii)、斯氏高原鳅(Triplophysastoliczkae),相似系数为仅为0.143;而今,两者共有种已达22种,其中18种为外来种,相似系数达到了0.275。与西藏区域的比较与之类似,历史上,两者共有种为5种,分别为斑重唇鱼、高原裸裂尻鱼(Schizopygopsisstoliczkai)、斯氏高原鳅、细尾高原鳅(Triplophysastenura)和小眼高原鳅(Triplophysamicrops),相似系数为0.065;当前两者的共有种为12种,其种8种为外来种,相似系数为0.098。与青海省相比,两者在历史记录上土著鱼类群落共有物种有2种,分别为细尾高原鳅和小眼高原鳅,相似系数为0.030;在青海省也经历了持续性的外来种入侵后,两者共有种已达19种,其中外来种17种,相似系数为0.154。与入侵种主要来源地长江中下游区域的比较发现,塔里盆地土著鱼类群落在历史记录中与该区代表性湖泊鄱阳湖、洞庭湖及太湖等无任何共有种,而到目前为止,塔里木盆地与这3个湖泊的共有种已经分别达到27种、24种、22种,相似系数分别为0.157、0.160、0.218,显示出与之鱼类区系均一化态势。
2.2 塔里木盆地入侵鱼类来源及组成
在44种外来鱼类中,起源于长江中下游区域的有30种,占68.2%;起源于额尔齐斯河的有7种,占15.9%,分别为贝加尔雅罗鱼(Leuciscusbaicalensis)、河鲈(Percafluviatilis)、湖拟鲤(Rutilusrutiluslacustris)、白斑狗鱼(Esoxlucius)、黑鲫(Carassiuscarassius)、丁鱥(Tincatinca)、花丁鮈 (Gobiogobiocynocephalus);起源于黑龙江流域有2种,为池沼公鱼(Hypomesusolidus)、北方泥鳅(Misgurnusbipartitus);起源于北美洲的2种,为虹鳟(Oncorhynchusmykiss)、云斑鮰(Ictalurusnebulosus);起源于欧洲的2种,为散鳞镜鲤(Cyprinuscarpio)、东方欧鳊 (Abramisbramaorientalis);起源于日本的1种,为白鲫(Carassiusauratuscuvieri)(附表1)。共计起源于国内的39种,占88.6%,起源于国外的5种,占11.4%。
所有入侵鱼类中,鲤形目27种,占61.4%;鲈形目9种,占20.5%;鲑形目4种,占9.1%;鲶形目3种、占6.8%;鳉形目1种,占2.2%。
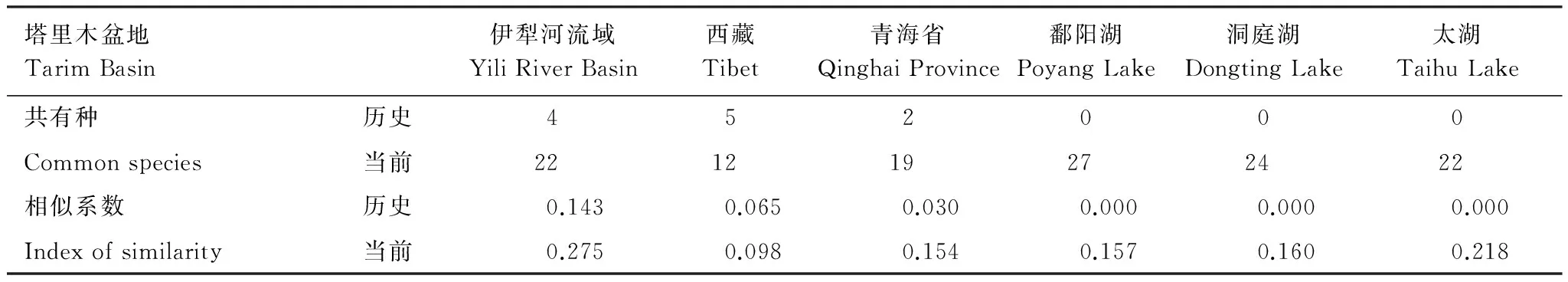
表1 塔里木盆地鱼类群落组成与相关区域相似性系数
伊犁河流域原土著鱼类物种有13种,当前共有鱼类物种39种[23];西藏原有土著鱼类物种63种,现有鱼类物种共72种[8, 24-25];青海省原有土著鱼类物种49种,现有鱼类物种79种[26-27];鄱阳湖、洞庭湖、太湖鱼类物种分别有136种、111种、60种[28-30];具体物种名录见相应参考文献
2.3 塔里木盆地土著与入侵鱼类生态特征比较
从营养纬度及空间维度分析,塔里木盆地原有鱼类群落与入侵鱼类群落的物种生态特征组成有一定差别,原有土著鱼类尚没有演化出专门以浮游生物为食的虑食性物种,同时也没有演化出以水生植物为食物的物种。外来鱼类中上述两类群均有代表性物种进入了塔里木盆地水体(表2)。无论土著鱼类及入侵鱼类均以杂食性、定居性、喜栖息于缓流生境的物种为主,同时水体各层均有物种占据(表2)。
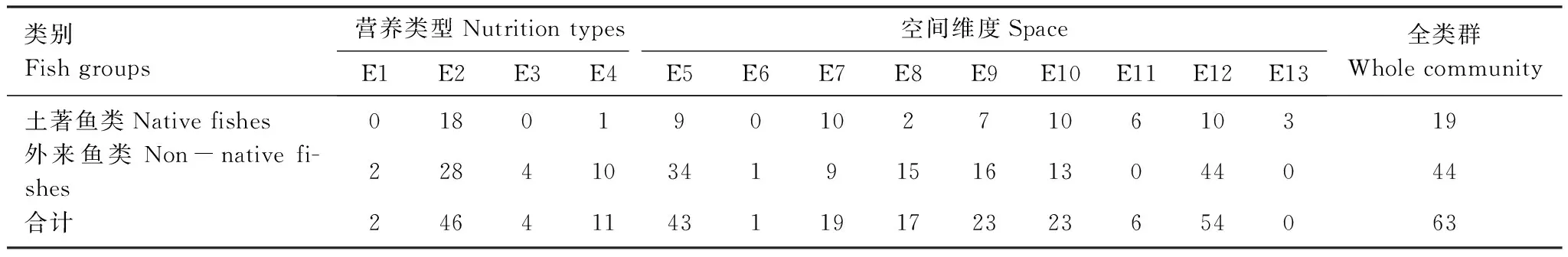
表2 塔里木盆地土著鱼类与外来鱼类生态特征构成
E1:滤食性 Filter feeder;E2:杂食性 Omnivore;E3:植食性 Herbivorous;E4:肉食性 Carnivore;E5:定居 Sedentary;E6:洄游 Migratory;E7:半洄游 Semi-migratory; E8:中上层 Mid and super stratum;E9:中下层 Mid and sub stratum;E10:底层 Benthic;E11:急流型 Riffle;E12:缓流型 Slow current;E13:泛栖型 Inhabit both in riffle and slow current
进一步以鱼类个体生态矩阵分析,显示历史上塔里木盆地演化形成的土著鱼类物种是以适应于杂食性(R=0.974)、口下位(R=0.947)、底层栖息(R=0.526)、产粘性卵(R=0.526)及沉性卵(R=0.474)、生活在缓流型水体(R=0.579)、具有半洄游习性(R=0.526)的鱼类为主(表3)。而入侵鱼类虽然也是以适应于杂食性(R=0.614)物种为主,但其他食性物种均有一定的比例(表4),尤其是适应于肉食性物种比例较高(R=0.250)。与土著鱼类群落不同的是,入侵鱼类口端位比例极高(R=0.955),主要以适应于水体中上及中下层物种为主,底栖比例较低(R=0.250)。所有入侵鱼类均适应栖息于缓流生境(R=1.000),无适应于急流生活的物种,且以产粘性卵物种为主(R=0.727)(表4)。大部分入侵种适应于定居生活(R=0.795),部分种类在繁殖季节有半洄游习性,需到达上游或急流区域产卵场产卵(R=0.182)。
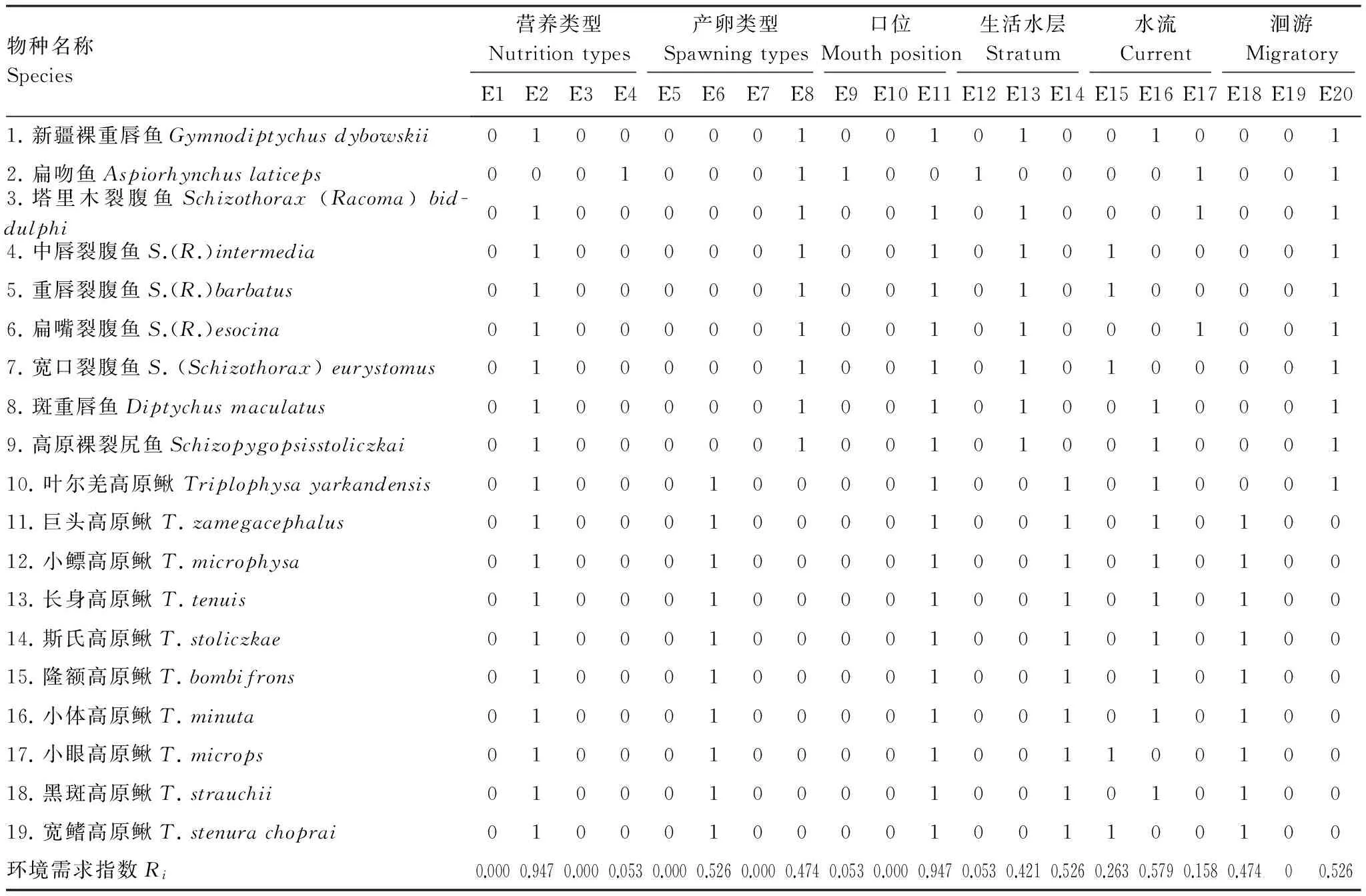
表3 塔里木盆地土著鱼类个体生态学矩阵
黑斑高原鳅Triplophysastrauchii=新疆高原鳅;宽鳍高原鳅Triplophysastenurachoprai=细尾高原鳅Triplophysastenura(记录于喀什噶尔河乌恰吉根); E1:滤食性 Filter feeder;E2:杂食性 Omnivore;E3:植食性 Herbivorous;E4:肉食性 Carnivore;E5:浮性 Pelagic eggs;E6:粘性 Adhesive eggs;E7:漂流 Drifting eggs;E8:沉性 Demersal eggs;E9:端位 With terminal mouth;E10:上位 With upper mouth;E11:下位 Hypostomatous;E12:中上层 Mid and super stratum;E13:中下层 Mid and sub stratum;E14:底层 Benthic;E15:急流 Riffle;E16:缓流 Slow current;E17:泛栖型 Inhabit both in riffle and slow current;E18:定居 Sedentary;E19:洄游 Migratory;E21:半洄游 Semi-migratory

表4 塔里木入侵鱼类个体生态学矩阵
河鲈Percafluviatilis产胶质卵,一个卵带可有上百卵子,能半漂浮,本文归为飘浮性卵类;中华青鳉Oryziassinensis产卵后,卵子成串挂在雌鱼身后,发育一段时间通过卵膜上的纤丝挂在水草上,本文归入粘性卵类;E1—E12的标注同表3
3 讨论
3.1 入侵鱼类导致塔里木盆地鱼类组成及鱼类区系演变
土著鱼类区系及分布详细本底资料的缺失给我国鱼类入侵生态学研究带来极大困难。塔里木盆地(主要针对塔里木河流域)土著鱼类物种确切数目及分布多年来一直存在不同意见[6, 12, 15, 35],王德忠记载有15种[6],王智超等也记载有15种[35],但两者间物种组成存在差异;马燕武等认为仅有12种[15],谢春刚等与之持相同意见[12]。由于近半世纪来入侵种的持续扩散,土著种分布持续的退缩,当今要厘清这个问题似乎不太可能了。尽管如此,依据不同时期文献资料的综合分析,剔除同物异名及同名异物的影响,依据最新研究结论研判,本文勘定塔里木盆地土著鱼类应有19种,与之前发表的资料存有差异[6,12,15,35]。这可能与本文针对研究区域为塔里木盆地,而此前研究者所针对研究区域为塔里木河流域,对处于塔里木盆地区域但与塔里木河缺乏直接联系的区域关注不足有关,例如武云飞和吴翠珍[17]早期对塔里木盆地南侧的喀刺昆仑山-昆仑山的调查工作实际上并未为其他研究者所足够重视;对分布区域甚为狭窄的巨头高原鳅(Triplophysazamegacephalus)[36]也甚少受到关注。诚然,随着新的研究资料出现,本文结论还可能发生一些变动,但这对本文的数据分析及其相关结论不会造成致命性的影响。
另一方面,综合大量的文献资料,塔里木盆地现今已发现入侵鱼类44种。与云南高原湖泊经历鱼类入侵引起土著种大量灭绝后果不同[7],塔里木盆地外来鱼类入侵并未导致土著鱼类的完全灭绝,这可能与该区生境广阔、复杂多样、能够为土著种提供足够避难所有关。例如,在开都河的调查显示,土著鱼类通过退缩在河流的上游区域栖息而得以继续生存[18];在持续性外来种的影响下扁吻鱼目前在克孜尔水库尚能残存唯一一个扁吻鱼自然种群[19]。因此,现今塔里木盆地拥有土著及入侵鱼类物种共计63种,而入侵鱼类所占比例则高达69.8%。
入侵鱼类的进入引起塔里木盆地鱼类群落物种数量的显著上升,进而极大改变了塔里木盆地鱼类群落的物种多样性状况。从G-F指数研究结果分析,原有土著鱼类群落F指数(DF)(科的多样性)为1.30,G指数(DG)(属的多样性)为0.74,科、属的多样性指数较江河平原区域鱼类群落显著偏低,如在珠江流域鱼类群落的科、属指数在东江中下游分别为7.39、3.39[31]、漓江分别为10.03、4.05[37]。但塔里木盆地土著鱼类群落G-F指数较高,达0.43,显示塔里木盆地具有鱼类区系组成简单而物种分化较为强烈的特点[12]。而经历入侵鱼类的入侵后,当前该区鱼类群落总体F指数上升为4.21,G指数也上升为3.38,表明入侵种的进入已经显著提升了塔里木盆地鱼类物种的多样性。然而,目前该区鱼类群落G-F指数为仅为0.20,这主要为入侵种中单种科所占比例较高所致,这提示过去人们在对鱼类进行引种的过程中主要以经济性状为选择因素,在各科属中经济性状突出的少数物种被首先引入,从而引致当前塔里木盆地内入侵种单种科比例高、总体上降低其鱼类群落G-F指数数值的现象。
另一方面,进入塔里木盆地的鱼类入侵种主要来源于长江中下游地区(68.2%)、额尔齐斯河流域(15.9%)、黑龙江流域、亚洲其他地方、欧洲及美洲等不同地理区域,导致了该区鱼类区系组成从单一的中亚高原山区复合体演变为混合5个鱼类区系复合体的混合体[6],研究者较早就发现了鱼类入侵引起塔里木盆地鱼类区系剧烈改变的生态学问题[6],而现今有越演越烈的趋势。入侵鱼类中鲤形目鱼类占据了绝大多数,这与入侵种主要起源国内有关(88.6%),我国为东亚鲤科鱼类的演化中心,长江流域鲤形目鱼类占鱼类总数可达71%以上[38],且经济性状良好,生态适应性强,是引种的主要类群。
3.2 入侵鱼类引起了塔里木盆地鱼类区系与其相关区域均一化演变
许多研究指出,入侵鱼类能够导致大尺度具有天然地理阻隔区域鱼类区系的均一化现象[1, 4, 39-40]。例如,在澳大利亚不同流域鱼类区系相似性随着鱼类入侵的深入而显著上升[39];在里海及其邻近海域鱼类区系的相似性也在鱼类入侵日益加剧的情况下明显上升[40]。塔里木盆地内鱼类组成随着入侵种的大量进入,与国内相关区域的鱼类群落相似性急速上升,特别是与长江中下游主要湖泊如鄱阳湖、洞庭湖、太湖的共有种均已经达20种以上,相似系数从0上升为0.157、0.160、0.218。这种相似性的演变与外来种主要来自于长江中下游密切相关,本文分析显示,塔里木盆地44种外来鱼类中,起源于长江中下游区域的有30种,占68.2%之多。而与之有密切关系的伊犁河流域也因有18种共有外来种而使得鱼类群落相似系数从历史上的0.143上升为0.275。不同鱼类区系在进化过程中所形成的特有性随着外来种的入侵而消解,往往伴随着生物多样性的严重损失[1]。虽然目前塔里木盆地尚没有土著鱼类因外来鱼类的进入而完全灭绝,但是绝大部分物种因之而濒危,如扁吻鱼、塔里木裂腹鱼、叶尔羌高原鳅等代表性物种已经相继濒危,其种群的续存完全有赖于近年来投入巨大的人工保护工作[19,41-42]。可以预见,随着土著鱼类种群进一步萎缩乃至灭绝,塔里木盆地鱼类群落与相关区域的相似性仍将进一步提高。
3.3 入侵鱼类在塔里木盆地广泛适应性及扩散趋势
在一般应用上,鱼类个体生态矩阵分析方法可能由于定量分析数据的缺失而损失了大量信息,从而对研究结果产生较大影响,如对稀有种的高估及对优势种的低估等。然而,这种方法避免了传统上利用定量采集标本数据分析鱼类群落时所出现的系列问题,扩展了历史调查数据的应用范畴,因而有着重要的应用价值[32-33]。在鱼类入侵生态学范畴,入侵种往往是常见种或优势种,土著种常成为稀有种,这种显著分化提示应用鱼类个体生态矩阵方法分析特定已经受到入侵物种干扰的生境中鱼类群落问题需谨慎,应当结合其他数据资料共同分析。
从鱼类个体生态矩阵分析显示入侵鱼类在塔里木盆地均具有广泛的适应性,如杂食性、定居性等特征,并且许多入侵种能够充分利用土著种没有利用的环境资源;而另一方面,当前塔里木河水利建设、水文环境的改变十分有利于具有这些特点的鱼类物种生存,提示入侵鱼类具有在塔里木盆地进一步扩散的巨大潜在能力。
首先,在演化过程中,塔里木盆地土著鱼类尚没有充分利用该区所有栖息生境及环境资源,例如对浮游生物的利用、对水生植物的利用等等均无演化出相应物种,这种生态位的空置为入侵种的成功入侵提供了重要机遇。塔里木盆地内几乎所有土著物种均具有与底栖生活及觅食相关的形态结构如口下位等,适应于摄食底栖生物(表3),而缺少对于水体的中上层区域资源的利用能力。另一方面,除扁吻鱼外,没有演化出其他的肉食性鱼类,本区捕食压力相对于长江中下游区域低得多,这也是该区物种特化及分化剧烈的主要因素之一[12],这种情况与我国云南高原湖泊特有种及物种分化明显的原因十分类似[43]。外来种则广泛利用了土著鱼类没有利用的资源,如鲢鱼(Hypophthalmichthysmolitrix)、鳙鱼(Aristichthysnobilis)、池沼公鱼等充分利用了土著鱼类利用较少的浮游动植物;草鱼(Ctenopharyngodonidellus)、鳊鱼(Parabramispekinensis)、团头鲂(Megalobramaamblycephala)等大量摄食该区未被利用的水生植物;大银鱼(Protosalanxhyalocranius)、中华青鳉(Oryziassinensis)则充分利用了湖滨及表层水体,在许多区域成为数量上的优势物种。入侵鱼类主要为口端位、活跃于中上层物种居多,并且有多个物种为肉食性鱼类,如乌鳢(Ophiocephalusargus)、河鲈等为代表性物种。在资源利用角度分析,入侵种很好利用了各种维度食物资源条件。
其次,塔里木盆地水域栖息生境复杂多样、人工干扰显著,为入侵鱼类的生存提供可能性。例如,对繁殖生态要求特别严格的四大家鱼在塔里木流域能够形成自然种群。虽然有调查显示草鱼、鲢鱼和鳙鱼不能在塔里木河支流开都河自然产卵[44],但是草鱼、鲢鱼在塔里木干流够自然繁殖,如肖汉纲和开文[45]在塔里木河的农垦十六团渡口、阿拉尔渡口、沙雅县塔河段、农垦十六团上游水库进库处等4个点调查发现上述各点均获得一定数量的各个发育期的鱼卵和鱼苗,确定是草鱼、鲢鱼在塔里木河中自然产的卵及孵化出的鱼苗,同时确定从沙雅到塔里木河拦河闸塔河段至少有3—7处产卵场,证实了这些鱼类已经能够在塔里木河流域自然繁殖。而对于大多数入侵鱼类而言,其喜于缓流生境、定居性,对繁殖期水文条件要求并不严格,很好适应了当前塔里木盆地水域生境的变化格局,例如塔里木河流域大量兴建的水工建筑改变了河流水文格局,适宜于静水栖息鱼类大量繁衍,如云斑鮰Ictalurusnebulosus在多浪水库成功定殖并成为重要渔业物种之一[46],而鲤鱼(Cyprinuscarpio)、鲫鱼(Carassiusauratus)、麦穗鱼(Pseudorasboraparva)、虾虎鱼类等物种几乎遍布塔里木盆地主要水域。
最后,入侵鱼类生态学上具广泛适应性的特点导致其在塔里木盆地广泛分布及具备进一步扩散潜力,已经及将会引致严重挤压土著鱼类的生存空间、物种濒危的严峻后果。捕食性鱼类如乌鳢、河鲈、云斑鮰、大口鲶(Silurussoldatovimeridionalis)等对土著鱼类具有明显捕食压力,如河鲈引入博斯腾湖对塔里木裂腹鱼、扁吻鱼等的捕食是导致这些土著鱼类在该湖区灭绝的关键性原因[14]。另一方面,土著鱼类多为杂食性鱼类,引入鱼类也以杂食性为主,两者食物竞争剧烈。麦穗鱼、虾虎鱼类等物种对鱼类的卵及仔幼鱼捕食压力较强,能显著降低以产粘性及沉性卵为主的土著鱼类早期补充资源量。池沼公鱼对浮游动物如哲水蚤类的大量摄食显著降低其他鱼类物种以浮游动物为主要食物的早期仔鱼的成活率[14, 47-48]。草鱼在巴楚县附近的小海子水库和永安坝南库导致沉水植被几乎完全消失[49],显著改变了部分土著鱼类繁殖环境,如许多土著的高原鳅类繁殖期间产粘性卵,需粘附在水生植物上发育[50-51]。扁吻鱼现存的唯一自然种群克孜尔种群被发现10年后调查中显示已经无低龄群体,11+—25+龄的个体占种群的81.35%,呈现严重衰退问题[52-53],外来鱼类在该栖息地鱼类群落中已占47.4%[19]。
外来鱼类的引入导致土著鱼类的濒危已经成为事实,针对外来鱼类对塔里木盆地土著鱼类及生态环境的影响及作用机制,需投入大量的人力物力开展系统研究。
[1] Scott M C, Helfman G S. Native invasions, homogenization, and the mismeasure of integrity of fish assemblages. Fisheries, 2001, 26(11): 6-15.
[2] 潘勇, 曹文宣, 徐立蒲, 殷守仁, 白璐. 鱼类入侵的过程、机制及研究方法. 应用生态学报, 2007, 18(3): 687-692.
[3] 陈国柱, 林小涛, 陈佩. 食蚊鱼(Gambusiaspp.)入侵生态学研究进展. 生态学报, 2008, 28(9): 4476-4485.
[5] Melles S J, Chu C, Alofs K M, Jackson D A. Potential spread of Great Lakes fishes given climate change and proposed dams: an approach using circuit theory to evaluate invasion risk. Landscape Ecology, 2015, 30(5): 919-935.
[6] 王德忠. 塔里木河鱼类区系变化及土著鱼类保护问题. 干旱区研究, 1995, 12(3): 54-59.
[7] 陈银瑞, 杨君兴, 李再云. 云南鱼类多样性和面临的危机. 生物多样性, 1998, 6(4): 272-277.
[8] 陈锋, 陈毅峰. 拉萨河鱼类调查及保护. 水生生物学报, 2010, 34(2): 278-285.
[9] 叶学齐. 塔里木盆地. 北京: 商务印书馆, 1959: 1-6.
[10] 汤奇成. 塔里木盆地水资源与绿洲建设. 资源科学, 1989, 11(6): 28-34.
[11] 邓铭江. 塔里木河流域未来的水资源管理. 中国水利, 2004, (17): 20-23.
[12] 谢春刚, 马燕武, 郭焱. 塔里木盆地鱼类生物地理学初步解析. 水产学杂志, 2015, 28(2): 40-46.
[13] 王德忠. 新疆的裂腹鱼亚科鱼类研究. 干旱区研究, 1998, 15(4): 26-32.
[14] 郭焱, 张人铭, 蔡林刚, 田永胜, 吐尔逊·提立瓦尔地, 赖英. 博斯腾湖鱼类资源及渔业. 乌鲁木齐: 新疆科学技术出版社, 2005.
[15] 马燕武, 郭焱, 张人铭, 吐尔逊, 谢春刚, 刘健, 李林. 新疆塔里木河水系土著鱼类区系组成与分布. 水产学报, 2009, 33(6): 949-956.
[16] 程勇, 严光文, 王智超, 姜作发, 梁志春. 塔里木河干流上游及附属水体外来鱼类物种调查及检索表的编制. 塔里木大学学报, 2011, 23(2): 8-13.
[17] 武云飞, 吴翠珍. 喀喇昆仑山-昆仑山地区渔业资源及渔业发展对策的初步研究. 自然资源学报, 1990, 5(4): 354-364.
[18] 马燕武, 郭焱, 陈朋, 谢春刚, 祁峰, 牛建功. 新疆开都河鱼类区系组成与分布. 淡水渔业, 2013, 43(5): 21-26.
[19] 马燕武, 张人铭, 李红, 吐尔逊, 郭焱. 扁吻鱼的栖息地及其群落保护生物学研究. 水生态学杂志, 2010, 3(1): 38-42.
[20] 王智超, 程勇, 武军元. 塔里木河流域发现河川沙塘鳢. 动物学杂志, 2009, 44(4): 150-151.
[21] 蒋志刚, 纪力强. 鸟兽物种多样性测度的G-F指数方法. 生物多样性, 1999, 7(3): 220-225.
[22] 王丹, 赵亚辉, 张春光, 周解. 广西野生淡水鱼类的物种多样性及其资源的可持续利用. 动物分类学报, 2007, 32(1): 160-173.
[23] 任慕莲. 伊犁河鱼类. 水产学杂志, 1998, 11(1): 7-17.
[24] 张春光, 许涛清, 蔡斌, 叶恩琦. 西藏鱼类的组成分布及渔业区划. 西藏科技, 1996, (1): 10-19.
[25] 范丽卿, 土艳丽, 李建川, 方江平. 拉萨市拉鲁湿地鱼类现状与保护. 资源科学, 2011, 33(9): 1742-1749.
[26] 唐文家, 王猛, 李柯懋. 青海省土著鱼类名录. 水产学杂志, 2005, 18(1): 13-17.
[27] 唐文家, 何德奎. 青海省外来鱼类调查(2001-2014年). 湖泊科学, 2015, 27(3): 502-510.
[28] 张堂林, 李钟杰. 鄱阳湖鱼类资源及渔业利用. 湖泊科学, 2007, 19(4): 434-444.
[29] 彭平波, 胡军华, 何木盈. 西洞庭湖鱼类资源调查与研究. 岳阳职业技术学院学报, 2012, 27(2): 27-32.
[30] 朱松泉, 刘正文, 谷孝鸿. 太湖鱼类区系变化和渔获物分析. 湖泊科学, 2007, 19(6): 664-669.
[31] 刘毅, 林小涛, 孙军, 张鹏飞, 陈国柱. 东江下游惠州河段鱼类群落组成变化特征. 动物学杂志, 2011, 46(2): 1-11.
[32] Suen J P, Herricks E E. Investigating the causes of fish community change in the Dahan River (Taiwan) using an autecology matrix. Hydrobiogia, 2006, 568(1): 317-330.
[33] 万安, 曹莉, 潘伟, 陶峰, 李家磊, 张晓可, 余世金, 安树青. 低头坝养鱼对鱼类局域栖息地环境的影响—基于鱼类个体生态矩阵的分析. 安徽农学通报, 2015, 21(7): 122-126.
[34] 李捷, 李新辉, 贾晓平, 李跃飞, 何美峰, 谭细畅, 王超, 蒋万祥. 西江鱼类群落多样性及其演变. 中国水产科学, 2010, 17(2): 298-311.
[35] 王智超, 姜作发, 张长征, 韩传信, 王能志, 白雪. 塔里木河流域土著鱼类名录文献考究及检索表的编制. 渔业科学进展, 2010, 31(1): 15-18.
[36] 赵铁桥. 新疆条鳅属(Nemachilus)鱼类一新种. 动物学研究, 1985, 6(4): 53-56.
[37] 李高岩, 韩松霖, 梁士楚, 宛玉剑, 文泉营, 韩耀全. 漓江光倒刺鲃金线鲃保护区鱼类资源现状调查. 广西师范大学学报: 自然科学版, 2011, 29(1): 66-71.
[38] 于晓东, 罗天宏, 周红章. 长江流域鱼类物种多样性大尺度格局研究. 生物多样性, 2005, 13(6): 473-495.
[39] Olden J D, Kennard M J, Pusey B J. Species invasions and the changing biogeography of Australian freshwater fishes. Global Ecology and Biogeography, 2008, 17(1): 25-37.
[40] Shiganova T. Biotic homogenization of inland seas of the Ponto-Caspian. Annual Review of Ecology, Evolution, and Systematics, 2010, 41(1): 103-125.
[41] 张人铭, 郭焱, 马燕武, 吐尔逊. 塔里木裂腹鱼资源与分布的调查研究. 淡水渔业, 2007, 37(6): 76-78.
[42] 陈生熬, 宋勇, 王智超, 姚娜. 塔里木河流域叶尔羌高原鳅人工繁殖试验. 水产养殖, 2008, (6): 25-26.
[43] 李树深. 云南湖泊鱼类的区系及其类型分化. 动物学报, 1982, 28(2): 169-176.
[44] 张苏江, 周万里, 李风格, 康尔梅. 草鱼、鲢鱼和鳙鱼不能在开都河自然产卵. 新疆农业科学, 1997, (3): 142-142.
[45] 肖汉纲, 开文. 草鱼、白鲢在塔里木河自然繁殖. 淡水渔业, 1980, (6): 25-25.
[46] 陈生熬, 谢从新, 王腾云, 宋勇, 王智超, 刘旭强, 任道全. 塔里木河流域多浪水库云斑鮰生物学特征分析. 塔里木大学学报, 2014, 26(1): 42-45.
[47] 唐富江, 姜作发, 阿达可白克·可尔江, 蔡林钢, 吴青. 新疆乌伦古湖河鲈食性变化的研究. 水产学杂志, 2008, 21(1): 49-52.
[48] 赖英. 博斯腾湖哲水蚤种群消失与池沼公鱼的关系. 水生态学杂志, 2009, 2(4): 136-139.
[49] 陈洪达, 张弘, 龚洛军, 何明钧. 小海子水库和永安坝南库的水生殖物调查. 水利渔业, 1991, (5): 8-12.
[50] 任波, 马燕武, 吐尔逊, 郭焱, 张人铭, 阿布都, 艾则孜, 刘宇. 阿克苏河叶尔羌高原鳅Triplophysa(Hedinichthgs)yarkandensis(Day)生物学研究. 水产学杂志, 2004, 17(1): 46-52.
[51] 郭焱, 蔡林钢, 吐尔逊, 张人铭, 刘昆仑, 张北平. 赛里木湖新疆高原鳅生物学研究. 水产学杂志, 2002, 15(2): 6-11.
[52] 任波, 任慕莲, 郭焱, 张人铭, 马燕武, 刘宇, 艾则孜, 吐尔逊, 阿不都. 扁吻鱼的生物学研究. 水产学杂志, 2006, 19(2): 9-22.
[53] 任波, 任慕莲, 张人铭, 马燕武, 刘宇, 艾则孜, 吐尔逊, 郭焱, 阿不都, 刘晓明. 扁吻鱼成鱼的采捕与运输. 水产学杂志, 2006, 19(1): 47-51.
[54] 中国科学院动物研究所. 新疆鱼类志. 乌鲁木齐: 新疆人民出版社, 1979.
[55] 武云飞, 吴翠珍. 青藏高原鱼类. 成都: 四川科学技术出版社, 1992: 1-599.
[56] 叶尚明, 李兴玖, 李晓春, 刘栓. 新疆塔什库尔干河水库建设对鱼类资源的影响. 水利渔业, 2004, 24(4): 63-65.
[57] 任波, 马燕武, 吐尔逊, 刘宇, 艾则孜, 阿布都. 新疆渭干河土著鱼类. 水产学杂志, 2005, 18(2): 53-58.
[58] 周颖. 喀什兰干水库鱼类寄生虫病病原种类及流行病学调查研究[D]. 乌鲁木齐: 新疆农业大学, 2014.
[59] 高行宜. 新疆脊椎动物种和亚种分类与分布名录. 乌鲁木齐: 新疆科学技术出版社, 2005: 1-25.

附表1 塔里木盆地鱼类名录[6,14-20,36,54-58]
鳡鱼Elopichthysbambusa、红鳍原鲌Cultrichthyserythropterus、黄尾密鲴Xenocyprisdavidi、鲃Spinibarbussp.、六须鲇Silurussoldatovi、胡子鲶Clariasfuscus、罗非鱼Oreochromis spp.等被认为在全新疆分布的但在塔里木盆地内尚无自然种群调查数据物种未列入[59],其他虽有明确的引入记录但无自然种群报道的物种在本文并未列入; △表示土著鱼类; *表示为作者所在单位采集到标本物种(文献亦有记录),其余为文献所记录物种; **物种分布区域仅列出所参考文献记述的主要地点,实际上许多鱼类在塔里木盆地内广泛分布
Fish invasions and changes in the fish fauna of the Tarim Basin
CHEN Guozhu*, QIU Yuping, LI Liping
NationalPlateauWetlandResearchCenter,SouthwestForestryUniversity,Kunming650224,China
Many native fish species in the Tarim Basin are endangered, owing to the invasion of alien fish species over the past decades. To better understand how to protect native fish and control the spread of alien fish, the number of alien fish species in the Tarim Basin and their potential ecological effects on native fish species should be assessed. In the present study, we used Jaccard′s Index of similarity to investigate the homogenization of fish fauna between the Tarim Basin and associated regions. Furthermore, we adopted the fish autecology matrix to test whether invasive fish would be able to spread widely in the Tarim Basin in the near future. These data were collected from the literature and from field investigations performed in recent years. Our results showed that a total of 63 fish species have been recorded in the Tarim Basin, including 19 native and 44 alien species. The introduction of alien fish species increased species, genus, and family diversity. However, theG-Fdiversity index (the ratio of genus diversity index and family diversity index) has decreased. The introduction of alien fish to the Tarim Basin is suggested to have changed the species composition from simple and highly differentiated to complicated and poorly differentiated. Moreover, among 44 alien species, 39 (68.2%) were originally from the middle and lower Yangtze River, 7 (15.9%) were originally from the Ertix River, and 7 (15.9%) were originally from America, Europe, and other regions of Asia. We also found that the index of similarity of fish fauna between the Tarim Basin and associated regions significantly increased before and after the introduction of alien species (P<0.01), indicating that alien species caused homogenization of the fish fauna within the studied regions. Furthermore, the fish autecology matrix revealed that alien fish species can be well adapted to the habitats of the Tarim Basin, and can cause the decline of native fish species through predation and competition. Therefore, control measures against alien fish should be implemented and/or intensified in order to protect the native fish species of the Tarim Basin.
Tarim Basin; biological invasions; fish; endangered species
国家自然科学基金青年基金资助项目(31400477)
2015-08-10;
日期:2016-06-13
10.5846/stxb201508101685
* 通讯作者Corresponding author.E-mail: chenguozhu79@163.com
陈国柱, 仇玉萍, 李丽萍.塔里木盆地鱼类入侵及区系演变趋势.生态学报,2017,37(2):700-714.
Chen G Z, Qiu Y P, Li L P.Fish invasions and changes in the fish fauna of the Tarim Basin.Acta Ecologica Sinica,2017,37(2):700-714.
猜你喜欢
热带作物学报(2022年9期)2022-10-17
矿产勘查(2020年11期)2020-12-25
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
环球时报(2017-10-10)2017-10-10
新疆地质(2015年3期)2015-12-10
植物营养与肥料学报(2014年1期)2014-03-11
石油与天然气地质(2014年6期)2014-03-11
断块油气田(2013年2期)2013-03-11
中国烟草学报(2012年1期)2012-04-09
