
深海微生物硝化作用驱动的化能自养固碳过程与机制研究进展*
2017-01-20 03:44:07王燕吴佳鹏洪义国
自然杂志 2016年2期
王燕,吴佳鹏,洪义国†
①广东省科技图书馆,广州 510070;②中国科学院南海海洋研究所热带海洋环境国家重点实验室,广州510301
深海微生物硝化作用驱动的化能自养固碳过程与机制研究进展*
王燕①②,吴佳鹏②,洪义国②†
①广东省科技图书馆,广州 510070;②中国科学院南海海洋研究所热带海洋环境国家重点实验室,广州510301
深海微生物硝化作用通过化能自养固定无机碳,是深海生态系统中重要的能量来源途径,直接影响深海生态系统的食物网结构和深海的碳储库。近年来氨氧化古菌的发现,对这一生物地球化学过程的传统认识提出了新的挑战,同时也带来了新的科学问题,为认识硝化作用化能自养的固碳机制提供了新的研究方向。通过系统评述深海硝化作用驱动的自养固碳过程和机制以及由此带来的生态环境效应,为相关领域的深入研究提供参考。
微生物硝化作用;氨氧化古菌;化能自养固碳;深海生态系统
海洋生产力占到全球生产力的一半以上,同时海洋也是生物圈中最大的碳储库。传统的观点认为,海洋中的生产力过程主要发生在真光层,依靠光能自养的形式固定CO2,而海洋内部(真光层以下)的能量需求主要源于真光层输入的颗粒有机物(POM)——这些有机物主要在海洋中层被矿化[1]且释放出能量(图1)。然而,海洋学家对北大西洋和藻海(Sargasso Sea)在区域尺度上的研究发现,海洋内部生物能量的需求超过了POM的能量输入(以碳计算),即以POM中有机碳计算输入的能量小于微生物消耗的能量(微生物总耗能=微生物生产耗能+呼吸耗能)。由此带来了深海内部能量收支不平衡的问题,而且随着深度的增加这种不平衡越发显著(图1)[2-4]。为了平衡海洋内部的能量收支,有学者提出在深海内部应当存在其他“新”的能源途径,以补充上层海洋能量输入的“不足”。
近年来科学家们逐渐认识到,深海是一个活跃的食物网圈层,在整个海洋的物质循环和能量流动过程中发挥了重要的作用[5-6]。上层海洋产生的POM会通过沉降和物理混合作用进入海洋内部,在微生物的作用下释放出NH4+。如果按照生物平均碳氮比(C/N=106/16),向下沉降的POM释放出的NH4+的通量应该是非常巨大的。这些NH4+都会通过微生物的硝化过程转变成稳定的NO3-储存在海洋内部,同时收获储存在NH4
+中的化学能用于固定CO2,合成自身需要的有机物。从全球海洋来看,海洋内部水体绝大部分是有氧环境(以O2的浓度≤20 μmol/L计算的低氧层只占全球海洋水体体积的1%以下)[7],这为有氧或者低氧条件下进行的微生物硝化过程提供了必要的环境条件,同时也限制了微生物无氧呼吸过程。因此,从通量估算和环境条件两个方面的理论分析来看,微生物硝化过程应该是海洋内部生态系统中重要的能量来源途径。硝化过程分为两步,分别为氨氧化过程和亚硝酸盐氧化过程。有关氨氧化过程自养固碳的研究较多,具有较为深入的认识;而对亚硝酸盐氧化过程还没有太多的研究,认识较少。本文主要以氨氧化偶联自养固碳的过程为重点,详细论述其作用过程、机制及其产生的生态效应。
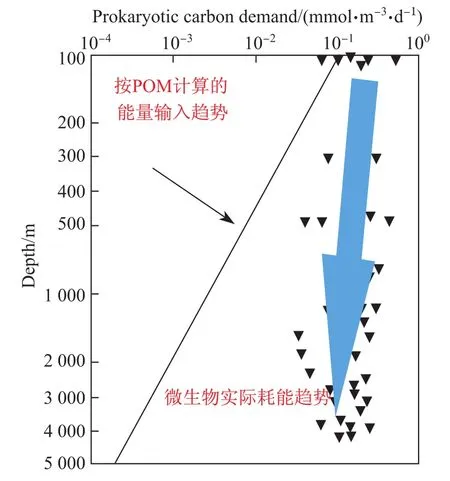
图1 深海能量的不平衡过程
1 深海微生物硝化过程驱动的自养固碳过程
早在20世纪80年代,微生物海洋学(microbial oceanography)研究的开拓者Karl等[3]通过分析向深海底部输运的有机碳(OC)、有机氮(ON)、ATP以及RNA的通量,发现在700~900 m水深的太平洋中层水体中有原位 “新”的颗粒有机碳合成,提出了深海微生物原位合成有机碳的假说。在接下来的研究中,Karl等在距墨西哥曼萨尼约330 km的东热带太平洋上定点进行了细菌化能自养生产力假说的验证。结果显示,在不同水体深度上的微生物都具有化能自养的活性[3]。维也纳大学和荷兰皇家海洋研究所的研究人员联合测定了大西洋中层和深层水体中溶解无机碳(DIC)的固定。[14C]HCO3-固定率测定结果表明,中层水团的微生物具有显著的固碳量(0.1~56.7 μmol·m3·d-1)。深海化能自养微生物的固碳过程为海洋内部提供了“新”的有机物,占到从真光层有机碳输入的15%~53%。催化报告免疫沉淀原位杂交技术(CARD-FISH)与放射自显影技术相结合的单细胞分析进一步证明DIC的固定主要发生在海洋中层,而深层主要为微生物的DIC同化[8]。
Karl的进一步研究从多个层面为氨氧化过程偶联固碳的假说提供了生物学和地球化学的证据。研究表明,在150 m以下的水层中,颗粒物吸附的NH4+的向下输运浓度和通量都普遍下降。也就是在海洋深部有NH4+的消耗,同时有NO2-生成,而且NH4+的消耗与NO2-的增加密切相关,表明存在活跃的氨氧化过程。利用免疫荧光技术在所有沉积物捕捉器收集的沉积物中都检测到了丰富的氨氧化细菌(AOB)。在预先装有化能自养底物的沉积物收集器中,原位培养后具有更高的细胞密度,表明了氨氧化微生物的活性。研究还发现,在墨西哥曼萨尼约沿岸海域,当消耗的同时,微生物的ATP和总碳量都有所升高,而最大升高发生在550 m处。在550 m和750 m深度可以检测到显著的化能自养生产力,且其在550 m处占总微生物碳生产力的90%[3]。这些研究结果表明,在有氧海洋中层水体中,基于微生物氨氧化过程的化能自养DIC固定为深海微生物食物网提供显著的有机碳源和能源,并证明从海洋真光层向深部输送的能量比原来认为的保存在颗粒有机碎屑中的能量要多。海洋深部的微生物利用有机颗粒分解后释放的还原性物质(主要为)作为电子供体通过电子传递链获得能量,进而固定无机碳成为自身需要的有机碳,部分解释了海洋内部能量收支不平衡的问题。
综合目前的研究结果,在深海内部水体中微生物氨氧化代谢过程是重要的化能自养固碳驱动力。根据从海洋表层输入到中层和深层的有机物通量,荷兰海洋学家对全球海洋的化能自养固碳通量进行了估算:全球海洋化能自养的固碳总量为0.77 Pg/a,相当于通过河流由陆地输入到海洋的总有机碳量,其中海洋水体中为0.40 Pg/a(真光层0.29 Pg/a,相当于37%;无光层0.11 Pg/a,相当于15%),沉积物中为0.37 Pg/a (主要发生在近岸海域的沉积物中)。化能自养固碳的能量主要来自氨氧化过程[9]。13C标记的稳定同位素探针实验也表明,硝化微生物是吸收沉积物无机碳的主要微生物群体[10]。然而,这些数据基本上来自于估算。对于微生物硝化作用在化能自养固碳过程中有多大的贡献,目前还没有确切的结论,仍然需要对海洋微生物氨氧化自养固碳过程进行更深入的定量研究。
2 海洋微生物硝化过程及自养固碳的分子机制
海洋环境中发现的AOB主要为β和γ变形菌(Proteobacteria):β-AOB主要有亚硝化螺旋菌属(Nitrosospira)和亚硝化单胞菌属(Nitrosomonas)两个属,而亚硝化螺旋菌一般是海洋环境中AOB的主导群体;γ-AOB目前仅发现亚硝化球菌(Nitrosococcus)一个属,包括海洋亚硝化球菌(N. Oceani)和嗜盐亚硝化球菌(N. Halophilus)两个种,目前只有在海洋和高盐的环境中发现γ-AOB[11]。早期的研究表明海洋水体具有活跃的氨氧化过程,但发现氨氧化的通量与微生物活性并不匹配,具有氨氧化能力的细菌只有很少的种类,而且在所有原核微生物类群中也只占有很小的数量比例[12-13]。抑制实验也表明,AOB不是硝化作用的主要群体[14]。因此有人提出,可能存在其他的氨氧化微生物类群在海洋的硝化作用过程中发挥重要作用。直到2005年,Könneke等[15]首次从海洋中分离和鉴定了一株具有氨氧化活性的古菌“Nitrosopumilus marinimus”,开辟了微生物硝化作用研究的新领域。随后,海洋氨氧化古菌(AOA)成了研究热点。大量实验研究结果表明,AOA是海洋环境中氨氧化作用的主导群体[16-18]。基因组学和生理生态特征表明,AOA与中温泉古菌(Crenarchaeota)有很大的不同。Brochier-Armanet等[19]在2008年建议将其划分为一个新门——奇古菌门(Thaumarchaeota),作为古菌域的第三个主要类群。生理学实验证明AOA比AOB对低浓度的氨具有更高的亲和力,进一步支持AOA在海洋环境中的主导地位[20]。
基因组分析显示,AOA具有与AOB不同的氨氧化电子传递途径(图2)。与AOB相比,AOA的氨氧化酶(AMO)缺乏羟胺-醌氧化还原酶模块,这导致可能有两种电子传递机制:一种可能是通过特异性含铜的羟胺氧化酶(CuHAO)进行羟胺氧化的反应;另外一种机制可能是AOA的AMO不生成羟胺,生成硝酰基氢化物(HNO)。然后电子通过醌池传给醌氧化还原酶(QRED),进一步传给NADH和细胞色素,与氧气和H+结合生成水。通过一系列的电子传递,把储存在NH4+的化学能转化成H+的势能,这种势能进一步通过ATP酶的作用把能量储存到能量货币ATP中,用于生命活动的需要[21]。
在纯培养水平上的研究表明,AOB都是依赖卡尔文循环固定CO2作为它们的碳源,其中1,5-二磷酸核酮糖羧化酶(RubisCO)是关键酶,CO2固定估计占到能量消耗的80%[22-23]。通过对纯培养氨氧化古菌(Nitrosopumilus maritimus)的基因组和生理学分析表明,氨氧化古菌(AOA)具有与AOB完全不同的固碳途径,它通过3-羟基丙酸/4-羟基丁酸(3-HP/4-HB)固定CO[21,24]2,其中3-羟基丙酰-CoA脱氢酶和4-羟基丁酰-CoA合成酶是AOA固碳的关键酶(图2)。
真光层向深部水体输入的有机物被异养微生物分解,释放出NH4+;化能自养的氨氧化微生物依靠氧化NH4+到NO2-获得能量,用来吸收并还原CO2成为有机物。与光合成相比较,微生物氨氧化作用的化能合成的产能效率是很低的,大约需要35 mol的NH4+才能支持1 mol CO2的固定[25]。利用获得的有限能量来固定自身需要的碳,即使在最优化的实验条件下,其生长速率也较低。Nitrosomonas和Nitrobacter的最小代时为7~13天[26];N. maritimus最小代时为21天[15]。如果假定氨氧化过程是化能自养的主要途径,就能计算出在中层海洋水体中维持细菌生产力所需要的最小能量。氧化NH4+到NO2-释放的自由能⊿G为-66 kcal/mol,每摩尔NH4+的氧化支持的细胞产量为0.03~0.06 mol的碳。这样,在550 m处化能自养微生物的生产力(425 μg·m-2·d-1)需要的NH4+通量估计为787 μmol·m-2· d-1。
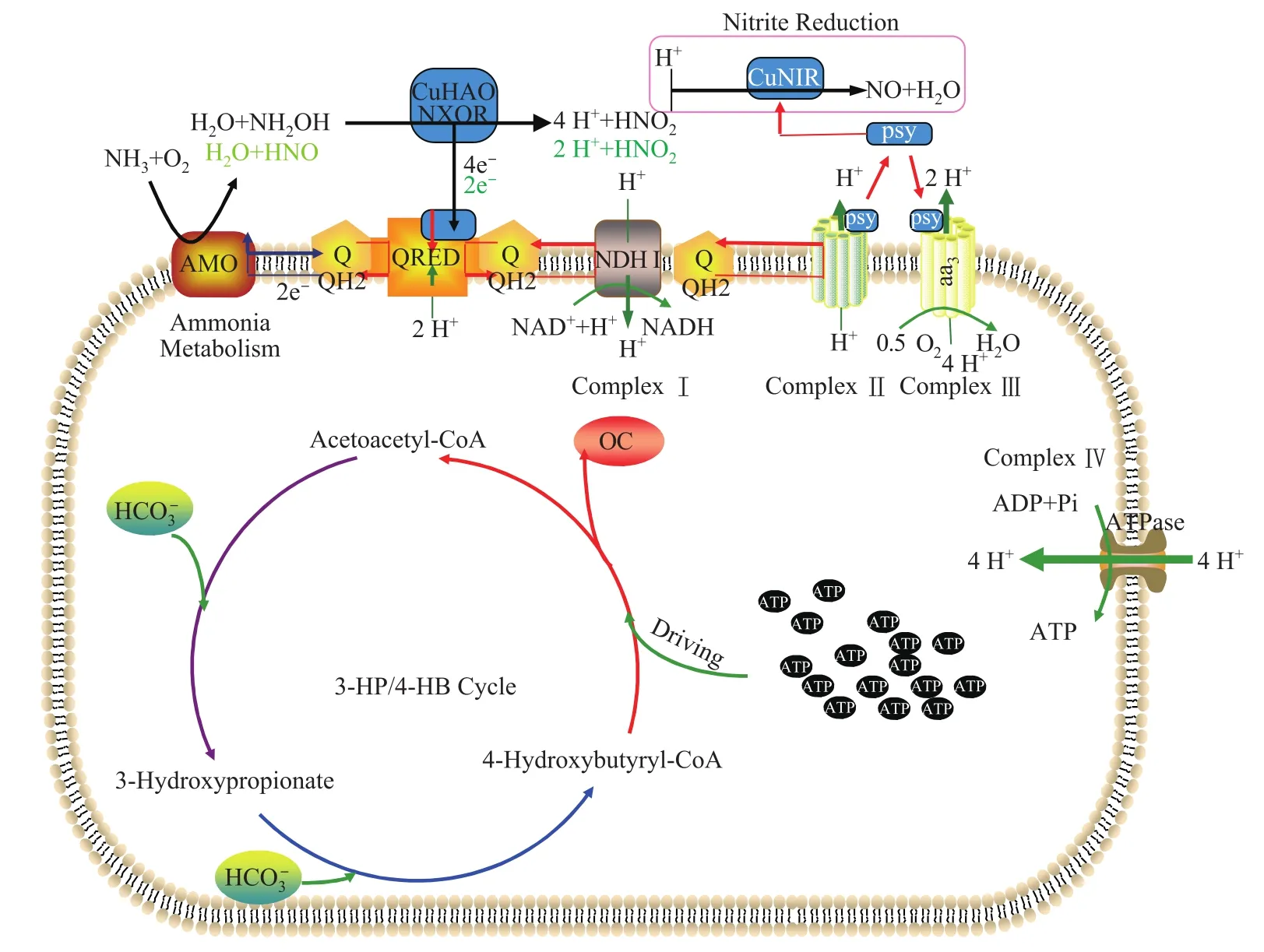
图2 氨氧化古菌的氨氧化代谢产能过程示意图
由于受到技术方法以及认识的限制,早期的氨氧化驱动固碳的研究基本没有微生物分子生态学方面的内容。对于微生物氨氧化过程,早期的研究都是基于氨氧化细菌(AOB)开展的,没有涉及到氨氧化古菌(AOA)是否发挥自养固碳的作用。利用同位素示踪结合分子生态学的方法,Bergauer等[27]对伊特鲁里亚海(Tyrrhenian sea)的化能自养过程进行了深入研究,结果表明氨氧化古菌是这一海域深部水体初级生产力的重要贡献者。其中检测到的与氨氧化古菌碳固定相关的基因乙酰-CoA羧化酶基因(accA)以及4-羟基丁酰-CoA脱氢酶基因(4-hbd)分成深海和浅海两种类型。在利用[14C]HCO3培养后,使用鸟枪蛋白质组的方法检测具有放射性标记的蛋白,发现有100多个寡肽段分属于40多个海洋泉古菌特异的蛋白序列,而且这些蛋白参与多个不同的代谢过程,包括化能自养过程。本研究表明了在数量上占主导的深海泉古菌是全球海洋无机碳固定的重要贡献者[27-28]。同时,古菌脂的稳定同位素分析也证明,在太平洋一个600 m深的站点,自养泉古菌的固碳占到所有古菌碳吸收的80%[29]。但是,这些研究还没有从地球化学和分子生态学相结合的角度阐明AOA和AOB各自功能的大小,也没有澄清微生物的氨氧化自养固碳与环境因素之间的关系,因而需要进行更深入的研究来阐释这些科学问题。
对南海的研究也表明,AOA也是硝化过程的主要功能微生物[30-32]。借助黑潮海洋专项航次,我们对南海深海站位H1进行了初步研究,并发现随着深度的增加,海水中DIC的浓度在逐渐增加。AOA的氨氧化功能基因amoA以及与固碳相关的accA基因都呈现明显的浅层和深层两个不同的群体,而且accA/amoA丰度的比率在表层和深层也具有明显的差异,其中表层在0.5~2.0之间,深层达到25~35(图3)。这表明深层的AOA具有更强的固碳需求,而表层的AOA并不是完全自养。这些初步的研究结果说明了氨氧化微生物的自养固碳具有典型的空间分布特征。
3 深海微生物自养固碳的生态环境意义
深海是地球上最大的生物圈层,共有1.3×1018m3。由于缺乏充分的光照,深海中没有光合作用。与真光层相比较,深海水体环境具有高压、低温的特点。绝大部分从真光层输入到深海内部的有机碳在海洋中层被原核生物呼吸分解,以CO2形式重新释放到大气中[33-35]。越来越多的证据表明,海洋中层栖息着多样且活跃的细菌和古菌,这些微生物的代谢是深海物质和能量循环的重要驱动力[5-6]。对微生物氨氧化驱动的自养固碳过程的研究表明,在海洋真光层的颗粒有机碎屑输入到海洋深部的能量比通常认识的要多[36-37]。在富含氧气的海洋水体中,化能自养的微生物可以利用储存在沉积颗粒有机物中的NH4+获得足够的能量生长(图4)。微生物硝化过程驱动的化能自养固碳研究,为解决海洋中层和深层的能量供应与消耗的不平衡问题提供了重要导向。
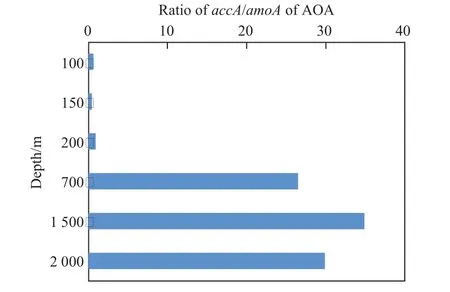
图3 氨氧化古菌固碳的关键功能基因的比值变化

图4 海洋硝化过程偶联自养固碳生产力过程示意图
与海洋表层的光能自养固碳相比,虽然化能自养过程的固碳通量要小的多,却给海洋深部输入了“新”的有机碳,是海洋碳循环收支的重要环节,对整个海洋深部的食物网结构有重要的影响。另外,微生物的自养固碳过程可能使有机物降解过程中形成的相对惰性的碳转化成为更加难降解的形式,并长期储存在海洋内部。这样,在千年的时间尺度上,海洋内部的CO2就不必再循环返回到海洋表层,这部分碳可能在较长的时间尺度上被“隔离”,因此微生物的氨氧化过程自养固碳可能是海洋微生物碳泵(MCP)[38]机制的重要贡献者。
4 本领域需要进一步解决的科学问题
虽然科学家在这一领域进行了开创性研究,并认识到海洋微生物的硝化过程是有氧海洋环境中化能自养生产力的重要驱动因素,但是,先前的研究认为氧化NH4+的过程是由氨氧化细菌(AOB)来完成的,而近年来有关海洋中氨氧化古菌(AOA)的发现对上述认识提出了挑战。研究表明AOA是海洋环境中氨氧化过程的主导微生物功能群[32,39-40]。新的科学发现催生了新的科学问题:海洋系统中的氨氧化偶联固碳的过程是否为AOA和AOB共同参与的结果?AOA和AOB各自的自养固碳贡献有多大,AOA是否为这一过程的主导者?AOA和AOB自养固碳对环境条件变化的响应有何不同?这些问题表明,我们目前对于海洋微生物氨氧化驱动的自养固碳的生物地球化学过程和分子生态学机制仍然是不清楚的,需要更广泛深入的研究对上述科学问题做出科学的解释。
基于上述科学问题,我们拟以南海深海为目标研究区域,系统地研究南海深海系统微生物氨氧化的自养固碳过程和机制,探讨影响氨氧化微生物自养固碳的关键环境因素。南海是西太平洋最大的热带边缘海,地处热带亚热带。各种理化参数显示南海的地球化学特征跟大洋类似,可以把南海看作一个“微型大洋”,它既有与大洋相似的特征,又有其本身的陆架边缘海特征。因此,南海是研究深海生物地球化学循环过程的一个理想海域,对认识整个海洋具有重要的科学价值。另外,对南海的研究已经表明,南海的水体整体溶氧水平较高,基本高于50 μmol/L[41],不具备微生物厌氧呼吸代谢的环境条件,同时为有氧条件下进行的微生物氨氧化过程提供了有利的环境条件。我们的研究将会进一步提升对深海内部微生物碳-氮生物地球化学过程相互作用的认识,深刻理解微生物氨氧化过程驱动的化能自养固碳的生物地球化学过程和微生物分子生态学机制,为解决海洋内部能量收支不平衡的问题提供更直接的科学证据,为更准确估算南海有机碳储库提供科学依据。
(2015年10月30日收稿)■
[1] DEL GIORGIO P A, DUARTE C M. Respiration in the open ocean [J]. Nature, 2002, 420: 379-384.
[2] BALTAR F, ARISTEGUI J, GASOL J M, et al. Evidence of prokaryotic metabolism on suspended particulate organic matter in the dark waters of the subtropical North Atlantic [J]. Limnology and Oceanography, 2009, 54: 182-193.
[3] KARL D M, KNAUER G A, MARTIN J H, et al. Bacterial chemolithotrophy in the ocean is associated with sinking particles [J]. Nature, 1984, 309: 54-56.
[4] BRANDON K S, MARTINEZ-GARCIA M, PRESTON C M, et al. Potential for chemolithoautotrophy among ubiquitous bacteria lineages in the dark ocean [J]. Science, 2011, 333: 1296-1300.
[5] KARNER M B, DELONG E F, KARL D M. Archaeal dominance in the mesopelagic zone of the Pacific Ocean [J]. Nature, 2001, 409: 507-510.
[6] KIRCHMAN D L, ELIFANTZ H, DITTEL A I, et al. Standing stock and activity of Archaea and Bacteria in the western Arctic Ocean [J]. Limnology and Oceanography, 2007, 52: 495-507.
[7] LAM P, KUYPERS M M M. Microbial nitrogen cycling processes in oxygen minimum zones [J]. Annual Review of Marine Science, 2011, 3: 317-345.
[8] REINTHALER T, VAN AKEN A, VETH C, et al. Prokaryotic respiration and production in the meso- and bathypelagic realm of the eastern and western North Atlantic basin [J]. Limnology and Oceanography, 2006, 51: 1262-1273.
[9] MIDDELBURG J J. Chemoautotrophy in the ocean [J]. Geophysical Research Letters, 2011, 38: L24604.
[10] GUILINI K, VAN OEVELEN D, SOETAERT K, et al. Nutritional importance of benthic bacteria for deep-sea nematodes from the Arctic ice margin: Results of an isotope tracer experiment [J]. Limnology and Oceanography, 2010, 55: 1977-1989.
[11] CAPONE D G, BRONK D A, MULHOLLAND M R, et al. Nitrogen in the marine environment [M]. 2nd ed. Amsterdam: Elsevier, 2008: 199-261.
[12] BOTHE H, FERGUSON S J, NEWTON W E. Biology of the nitrogen cycle [M]. Amsterdam: Elsevier, 2007: 209-222.
[13] KOWALCHUK G A, STEPHEN J R. Ammonia-oxidizing bacteria: a model for molecular microbial ecology [J]. Annual Review Microbiology, 2001, 55: 485-529.
[14] STARKENBURG S R, CHAIN P S G, SAYAVEDRA-SOTO L A, et al. Genome sequence of the chemolithoautotrophic nitriteoxidizing bacteriumNitrobacter winogradskyiNb-255 [J]. Applied and Environmental Microbiology, 2006, 72: 2050-2063.
[15] KÖNNEKE M, BERNHARD A E, DE LA TORRE J R, et al. Isolation of an autotrophic ammonia-oxidizing marine archaeon [J]. Nature, 2005, 437: 543-546.
[16] FRANCIS C A, ROBERTS K J, BEMEN J M, et al. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean [J]. Proceedings of the National Academy of Sciences of USA, 2005, 102: 14683-14688.
[17] WUCHTER C, ABBAS B, COOLEN M J L, et al. Archaeal nitrification in the ocean [J]. Proceedings of the National Academy of Sciences of USA, 2006, 103: 12317-12322.
[18] LAM P, JENSEN M M, LAVIK G, et al. Linking crenarchaeal and bacterial nitrification to anammox in the Black Sea [J]. Proceedings of the National Academy of Sciences of USA, 2007, 104: 7104-7109.
[19] BROCHIER-ARMANET C, BOUSSAU B, GRIBALDO S, et al. Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota [J]. Nature Reviews Microbiology, 2008, 6(3): 245-252.
[20] MARTENS-HABBENA W, BERUBE P M, URAKAWA H. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria [J]. Nature, 2009, 461: 976-979.
[21] WALKER C B, DE LA TORRE J R, KLOTZ M G, et al.Nitrosopumilus maritimusgenome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea [J]. Proceedings of the National Academy of Sciences of USA, 2010, 107: 8818-8823.
[22] FORREST W W, WALKER D J. The generation and utilization of energy during growth [J]. Advances in Microbial Physiology, 1971, 5: 213-274.
[23] KELLY D P. Bioenergetics of chemolithotrophic bacteria [M]//BULL A T, MEADOW P M (eds). Companion to microbiology. London: Longman, 1978: 363-386.
[24] SASHA G T, BREAKWEL K, ELBOURNE L D H, et al. Life in the dark: metagenomic evidence that a microbial slime community is driven by inorganic nitrogen metabolism [J]. International Society for Microbial Ecology Journal, 2013, 7: 1227-1236.
[25] BAAS BECKING L G M, PARKS G S. Energy relations in themetabolism of autotrophic bacteria [J]. Physiological Reviews, 1927, 7: 85-106.
[26] BOCK E, KOOPS H P, HARMS H. Nitrifying bacteria [M]// SCHELGEL H G, BOWIEN B (eds). Autotrophic bacteria. Berlin: Springer-Verlag, 1989: 81-96.
[27] BERGAUER K, SINTES E, VAN BLEIJSWIJK J, et al. Abundance and distribution of archaeal acetyl-CoA/propionyl-CoA carboxylase genes indicative for putatively chemoautotrophic Archaea in the tropical Atlantic's interior [J]. FEMS Microbiology Ecology, 2013, 84: 461-473.
[28] BENNER R. Chemical composition and reactivity [M]//HANSELL D A, CARLSON C A (eds). Biogeochemistry of marine dissolved organic matter. Amsterdam: Elsevier Science, 2002: 59-90.
[29] INGALLS A E, SHAH S R, HANSMAN R L, et al. Quantifying archaeal community autotrophy in the mesopelagic ocean using natural radiocarbon [J]. Proceedings of the National Academy of Sciences of USA, 2006, 103: 6442-6447.
[30] 胡安谊, 焦念志. 氨氧化古菌——环境微生物生态学研究的一个前沿热点[J]. 自然科学进展, 2009, 19: 370-379.
[31] HU A Y, JIAO N Z, ZHANG C L. Community structure and function of Planktonic Crenarchaeota: Changes with depth in the South China Sea [J]. Microbial Ecology, 2011, 62: 549-563.
[32] CAO H, HONG Y, LI M, et al. Community shift of ammonia-oxidizing bacteria along an anthropogenic pollution gradient from the Pearl River Delta to the South China Sea [J]. Applied and Environmental Microbiology, 2012, 94: 247-259.
[33] ARISTEGUI J, DUARTE S C M, GASOL J M, et al. Active mesopelagic prokaryotes support high respiration in the subtropical northeast Atlantic Ocean [J]. Geophysical Research Letters, 2005, 32: L03608.
[34] HANSELL D A, VARLSON C A. Deep-ocean gradients of dissolved organic carbon [J]. Nature, 1998, 395: 263-266.
[35] CARLSON C A, DUCKLOW H W, MICHAELS A F. Annual flux of dissolved organic carbon from the euphotic zone in the northwestern Sargasso Sea [J]. Nature, 1994, 371: 405-408.
[36] MICHAEL H, STEFAN M S. Beyond the Calvin cycle: autotrophic carbon fixation in the ocean [J]. Annual Review of Marine Science, 2011, 3: 261-289.
[37] THOMAS R, VAN AKEN H M, HERNDL G J. Major contribution of autotrophy to microbial carbon cycling in the deep North Atlantic's interior [J]. Deep Sea Research Part II, 2010, 57: 1572-1580.
[38] JIAO N Z, HERNDL G J, HANSELL D A, et al. Microbial production of recalcitrant dissolved organic matter: long-term carbon storage in the global ocean [J]. Nature Reviews Microbiology, 2010, 8: 593-599.
[39] HANSMAN R L, GRIFFIN S, WATSON J T, et al. The radiocarbon signature of microorganisms in the mesopelagic ocean [J]. Proceedings of the National Academy of Sciences of USA, 2009, 106: 6513-6518.
[40] HERNDL G J, REINTHALER T, TEIRA E, et al. Contribution of Archaea to total prokaryotic production in the deep Atlantic Ocean [J]. Applied and Environmental Microbiology, 2005, 71: 2303-2309.
[41] 刘洋, 鲍献文, 吴德星. 南海溶解氧垂直结构的季节变化分析[J]. 中国海洋大学学报, 2011, 41(1/2): 25-32.
(编辑:沈美芳)
Microbial nitrification coupled to the hemoautotrophic carbon fixation in the deep ocean
WANG Yan①②, WU Jiapeng②, HONG Yiguo②
①The Science and Technology of Library of Guangdong, Guangzhou 510070, China; ②State Key Laboratory of Tropical Oceanography, South China Sea Institute of Oceanography, Chinese Academy of Sciences, Guangzhou 510301, China
Nitrification, the two-step oxidation of ammonia to nitrate via nitrite, plays a critical role in the global carbon cycle by fixing CO2with the form of chemolithoautotrophy, and consequently influencing the organic carbon pool and food web structure in the dark ocean. It has long been assumed that the first and rate-limiting step of nitrification, ammonia oxidation, was restricted to a few groups within the domain Bacteria. However, the recent discovery of ammonia-oxidizing archaea (AOA) has seriously challenged our understanding of the microbial ecology and biogeochemistry of nitrification in the ocean. Therefore, the biogeochemical mechanism of CO2fixation by microbial nitrification is largely unknown. The multi-disciplinary studies in this field have expanded and strengthened the existing oceanographic carbon-nitrogen network, and increased our understanding on microbial and biogeochemical process of autotrophic production by marine nitrification. In this paper we reviewed the progress of microbial nitrification and chemolithpautotrophy carbon fixation in the deep sea, which would offer a reference for the researchers in this field.
microbial nitrification, ammonia oxidation archaea, hemoautotrophic carbon fixation, deep sea ecosystem
10.3969/j.issn.0253-9608.2016.02.007
*国家自然科学基金(41576123、31270163),国家重点基础研究发展计划(973计划)(2015CB452902),广东省自然科学基金重点项目(S2013020012823),广州市科学研究专项重点项目(201504010005)和中国科学院青年促进会项目资助
†通信作者,E-mail:yghong@scsio.ac.cn
猜你喜欢
土壤学报(2022年3期)2022-08-26 12:15:26
大自然探索(2022年5期)2022-07-11 03:10:33
知识就是力量(2022年6期)2022-06-16 20:19:36
少儿科学周刊·少年版(2022年19期)2022-05-30 10:48:04
小哥白尼(趣味科学)(2019年7期)2019-11-16 09:04:14
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
学苑创造·A版(2017年4期)2017-05-13 22:56:42
中国环境科学(2016年3期)2016-02-08 15:07:14
应用海洋学学报(2014年2期)2014-11-26 01:20:46
应用海洋学学报(2014年4期)2014-11-22 07:43:56
