
水产动物维生素营养研究进展
2017-01-08 07:22冷向军
饲料工业 2017年16期
■冷向军
(水产科学国家级实验教学示范中心(上海海洋大学)农业部鱼类营养与环境生态研究中心(上海海洋大学)上海水产养殖工程技术研究中心(上海海洋大学),上海201306)
维生素是维持动物机体正常生长、发育和繁殖所必需的微量小分子有机化合物。不同于蛋白质、脂肪和碳水化合物,动物对维生素需要量很少,每日所需量仅以毫克或微克计。
1 维生素概述
维生素的主要作用是作为辅酶参与物质代谢和能量代谢的调控、作为生理活性物质直接参与生理活动、作为生物体内的抗氧化剂保护细胞和器官组织的正常结构和生理功能,还有部分维生素作为细胞和组织的结构成分。维生素种类多,化学组成、性质各异,一般按其溶解性分为脂溶性维生素和水溶性维生素两大类。脂溶性维生素包括维生素A、D、E、K,水溶性维生素包括B族维生素和维生素C、肌醇、胆碱。B族维生素共8种(B1、B2、B6、B12、烟酸、叶酸、泛酸、生物素),需要量相对较低,主要功能是作为辅酶参与物质代谢,而维生素C、肌醇、胆碱的需要量相对较高。
在哺乳动物,维生素的缺乏,通常会导致特定的缺乏症状出现,但对水产动物而言,维生素缺乏症状的特异性并不强。传统的维生素营养主要研究维生素的需要量、缺乏症及其预防。目前,有关水产动物维生素营养的研究,主要集中在维生素调控机体免疫功能,改善肌肉品质,提高繁殖性能等方面。
2 维生素与免疫功能
维生素的生理功能十分广泛,维生素的缺乏将导致物质代谢、能量代谢以及其它生理生化活动异常,细胞和器官组织受到损害。因此,维生素缺乏将直接或间接导致动物免疫功能和抗病力下降。目前有关维生素对水产动物的免疫功能影响的研究,主要集中在维生素A、维生素E、维生素C等。
维生素A在细胞分化方面具有重要的生理作用,视黄酸在细胞核内与其受体结合后通过促进基因转录来调节机体的新陈代谢,从而影响细胞的增殖和分化。当饲料中缺乏VA时,罗非鱼体表出血,鳃盖骨畸形,鳍糜烂,体色变深,红细胞在低渗溶液中的抗溶血性及血清蛋白、溶菌酶活性和超氧阴离子的产生随饲料VA添加量的增加而增加(Guimarães等,2014);在团头鲂的研究表明,血清补体C3、补体C4、白细胞、红细胞和血红蛋白含量随饲料VA水平的增加而增加,并在饲料VA为1 934 IU/kg或3 885 IU/kg时到达最大值;以噬水气单胞菌攻毒后,VA充足组的成活率显著高于VA缺乏组(Liu等,2016);饲料中VA的缺乏,也降低了草鱼肠道先天性体液免疫反应,加剧了肠道的炎症(Zhang等,2017)。
维生素E是生物膜的重要组成部分,是生物膜中保护脂质的主要抗氧化剂,用以清除脂质氧化带来的过氧化自由基,这对于保持生物膜的完整性和正常功能具有重要意义。Niu等(2014)在饲料中添加0~960 mg/kg VE,饲养大菱鲆15周,血清溶菌酶和SOD活性、头肾NBT活性白细胞和吞噬指数随VE添加量的增加而增加,并在VE添加量480 mg/kg时达到最大值,480 mg/kg VE组的脾、头肾白细胞介素1β(IL-1β)、肿瘤坏死因子α(TNF-α)及肝脏补体C3的mRNA表达量较对照组显著上调。Wang等(2006)的研究表明,饲料中添加维生素E和n-3 HUFA可促进牙鲆的非特异性免疫反应和对爱德华氏菌的抵抗力,并且维生素E和n-3 HUFA具有明显的协同效应。
维生素C通过促进胶原蛋白的形成而参与表皮、黏液、鳞片的形成,在防病抗病过程中发挥着第一道屏障的作用,维生素C也是一种重要的抗氧化剂,可清除机体内的超氧离子、羟自由基和氢过氧化物等,保护生物膜免遭脂质过氧化的破坏。Liang等(2017)在饲料中添加0~320 mg/kg VC饲养黄颡鱼12周,发现随饲料VC添加量增加,血浆SOD、过氧化氢酶和过氧化物酶活性增加,溶菌酶活性、补体活性、头肾细胞吞噬指数和呼吸爆发活性在VC添加量为160 mg/kg组最高,饲料中添加20、40 mg/kg VC,显著提高了噬水气单胞菌攻毒后的鱼体成活率。日本鳗鲡摄食添加200~1 600 mg/kg VC的饲料16周后,肝、肾、肌肉和精巢VC含量与饲料VC含量呈线性相关,高剂量VC添加组的血液白细胞数量、红细胞压积、ALT、AST和SOD活性显著升高(Shahkar等,2015);此外,维生素C还显著提高了鲈鱼(Ai等,2004)、大黄鱼(Ai等,2006)的吞噬指数、呼吸爆发活性、溶菌酶活性和替代途径补体活力,显著降低了哈维氏弧菌感染后的大黄鱼死亡率。
此外,叶酸、肌醇等维生素也与机体免疫功能密切相关。当中华绒螯蟹饲料中叶酸添加量达2 mg/kg后,SOD、谷胱甘肽S-转移酶、酚氧化酶活性和血细胞数量显著升高,在叶酸添加量为2 mg/kg时,酸性磷酸酶、碱性磷酸酶、溶菌酶活性最高,而蟹体死亡率最低(Wei等,2016);在建鲤注射噬水气单胞菌后,白细胞吞噬活性、血凝滴度、溶菌酶活性、抗嗜水气单胞菌抗体滴度和免疫球蛋白M均随饲料肌醇水平增加而增加(Jiang等,2010)。
3 维生素与肌肉品质
许多维生素参与了蛋白质和脂肪的代谢过程,因而对肌肉品质有着直接的影响。与肌肉品质密切相关的维生素主要有维生素C和维生素E。
维生素C对于许多物质的羟化反应都有重要作用,其中最为重要的是胶原蛋白合成过程中的脯氨酸和赖氨酸残基被羟化成为羟脯氨酸和羟赖氨酸的反应,维生素C是此种羟化反应必需的辅助因子之一。胶原蛋白是动物体内最为丰富和重要的蛋白质之一,胶原蛋白作为肌肉主要连接组织,影响其功能和质构特性,也影响肌肉的黏连程度(Aidos等,1990;Gordon等,2010)。高含量的胶原蛋白能够增强鱼类肌肉硬度(Hatae等,1986;Sato等,1986),野生大西洋鲑的肌肉硬度高于养殖个体,主要是高含量的胶原蛋白所致(Johnston等,2006)。由于维生素C对于胶原蛋白的形成起着重要作用,故显著影响着肌肉品质。任泽林等(1998)在饲料中补充VC 500 mg/kg,显著提高了中国对虾肌肉中的胶原蛋白含量,提高了肌肉耐折断力,降低了肌肉失水率;随VC添加量从50 mg/kg增加到300 mg/kg,草鱼鱼种肌肉失水率、肌纤维直径和肌原纤维耐折力呈增加趋势,而肌肉胶原蛋白含量和肝脏VC含量显著增加(胡斌等,2008);饲料中添加VC 100 mg/kg或200 mg/kg,均显著提高了草鱼成鱼肌肉胶原蛋白含量和肌纤维直径,肝脏VC含量随VC添加量的增加而增加(李小勤等,2010)。
维生素E的主要生理功能之一是清除细胞内自由基,防止自由基、氧化剂对生物膜中多不饱和脂肪酸、富含巯基的蛋白质成分以及细胞核和骨架的损伤,保持细胞、细胞膜的完整性和正常功能。对于肌肉而言,细胞膜的完整性、肌肉的保水性能等,是肌肉品质的重要内容。李小勤等(2009)在实用饲料中添加25~200 mg/kg VE,饲喂平均体重为614.9 g的草鱼60 d,发现随饲料VE添加量的增加,肌肉、肝脏VE含量增加,丙二醛含量显著降低,鲜肉滴水损失和冷冻肉渗出损失也随VE添加量的增加而降低;在海鲈鱼的试验中,鱼体脂肪VE含量与饲料VE含量呈线性相关,饲料中添加100 mg/kg VE显著降低了肌肉MDA含量(Gatta等,2006);在大西洋白姑鱼上也有类似报道(Rodríguez等,2017);鲤鱼肌肉的滴水损失、渗出损失、挥发性盐基氮和硫代巴比妥酸值(TBARS)也随饲料维生素E添加量的增加而显著降低(王文娟等,2013)。
此外,肌醇、胆碱等维生素,与脂肪代谢密切相关。在实用饲料中添加0.4%~0.6%氯化胆碱,显著降低了草鱼肌肉、肝胰脏和全鱼脂肪含量(朱瑞俊等,2010);团头鲂幼鱼摄食添加400、800 mg/kg肌醇的饲料后,肝脏和肌肉脂肪含量显著降低,而肌肉蛋白质含量显著升高(崔红红等,2013)。在目前养殖鱼类普遍较肥的情况下(个别高脂肪含量鱼类除外),降低肌肉脂肪含量,有利于提高肌肉品质。
4 维生素与繁殖性能
维生素广泛参与机体的物质代谢,与亲鱼的繁殖性能密切相关,其中较为重要的有维生素A、维生素E、维生素C等。
维生素A影响着细胞的增殖和分化,参与精子生成和免疫反应等,因而与繁殖性能密切相关。野生凡纳滨亲虾的组织维生素A含量显著高于养殖个体,特别是当卵巢发育到后期时表现更加明显,故维生素A能促进养殖亲虾性腺的成熟并提高繁殖成功率(Linancabello等,2003);野生褐牙鲆亲鱼卵中维生素A含量也显著高于养殖亲鱼,但二者在肌肉和肝脏维生素A含量上无显著差异(王际英等,2010);在日本牙鲆(Furuita等,2001)和虹鳟(Fontagné-Dicharry 等,2010)饲料中强化补充维生素A,可以显著提高亲鱼的产卵量和鱼卵中维生素A的沉积量,但过高的维生素A也会导致胚胎和仔鱼死亡率的增加。此外,维生素A还显著影响仔稚鱼的骨骼形成(Fernández等,2008)和免疫机能调节(Manicassamy等,2009)。
维生素E又名生育酚,具有抗不育功用,作为抗氧化剂,可避免细胞膜上的不饱和脂肪酸被氧化,从而保持细胞膜的完整性和正常的生理功能,这对胚胎的正常发育尤为重要。在饲料中添加1 200 mg/kg维生素E,促进了半滑舌鳎亲鱼性腺的发育,提高了亲鱼产卵量、受精率、孵化率以及初孵仔鱼长度和仔鱼成活率,而添加200 mg/kg的维生素E只对亲鱼性腺的发育和初孵仔鱼长度有一定的促进作用(肖登元等,2015);饲料中添加200 mg/kg维生素E,提高了克氏原螯虾产卵雌虾的数量比例和雌虾的产卵量(李铭等,2007)。对日本鳗鲡亲鱼在繁殖前注射维生素C和维生素E,提高了亲鱼和卵中维生素E的沉积量,孵化率、成活率,仔鱼质量也得到提高(Furuita等,2009);在斜带石斑鱼(肖伟平等,2003)、黄金鲈(Lee等,2004)、黄鳝(张国辉,2005)的研究均表明,亲鱼饲料中适宜的维生素E水平能提高产卵量、孵化率、成活率和仔鱼质量。
维生素C在体内的功能十分广泛,直接影响着亲鱼的繁殖能力。硬骨鱼类在生殖细胞发生过程中,维生素C的抗氧化作用对精子和卵子的受精能力,以及保护生殖细胞的遗传完整性都很重要。野生褐牙鲆亲鱼肝脏和卵中维生素C含量显著高于养殖亲鱼,但肌肉中维生素C含量两者无显著差异,维生素C在亲鱼体内的沉积顺序依次为卵、肝脏、肌肉(王际英等,2010);维生素C对精子DNA具有保护作用,高浓度维生素C可保护精子内重要成分免遭氧自由基损伤(Terova等,1998);肖登元等(2014)发现,饲料中添加维生素C能不同程度地提高半滑舌鳎亲鱼肝脏、卵巢、精巢和卵中维生素C含量,随饲料维生素C添加量增加,上浮卵率和孵化率上升,初孵仔鱼畸形率下降,高剂量维生素C(0.525%)有利于促进亲鱼性激素的合成,改善亲鱼的繁殖性能。金鱼摄食添加200 mg/kg VC的饲料后,性腺重、性腺指数、产卵量显著增加,卵的质量、孵化率和仔鱼体重、体长也显著提高(James等,2011)。
其他维生素对水产动物繁殖性能影响的研究较少,仅见于维生素B1、B6等少数几种维生素。对怀卵大西洋鲑雌鱼注射维生素B1,可降低子代死亡率(Ketola等,1998)。在其他维生素方面的研究少,并不意味着其他维生素不重要。在亲本养殖和苗种培育中,应保证饲料中充足的维生素供应。
5 维生素的需要量与添加量
水产动物对维生素的需要包括对维生素种类的定性需要和定量需要。由于测定维生素需要量时采用的评定标准及试验条件不同,所得结果也有差异。通常评判维生素需要量的标准有:不出现缺乏症时的需要量,最适生长时的需要量,与维生素相关的酶类最适活性或相关生化指标最适数值时的需要量,在肝(胰)脏或肌肉或其他组织最大积蓄量时的需要量。一般而言,以酶活性作为评定指标所得的结果与以生长曲线测定的结果相似,而二者一般都低于以肝(胰)脏最大积蓄量为评定指标所测定的结果。
在实际生产中,一般是将养殖动物对维生素的需要量直接作为饲料中维生素的添加需要量,即将维生素需要量作为维生素预混料的供给量进行配方设计,忽略饲料原料中的维生素含量以及肠道微生物可能的维生素合成量。实际上,在饲料生产和使用中,由于加工工艺、贮存条件、养殖模式、环境应激等情况,应适当提高某些维生素的添加量,或增加保险系数。对于加工温度较高,或后熟化时间较长的饲料品种,宜选用稳定性好的维生素剂型。
表1列出了NRC(2011)推荐的部分水产动物的维生素需要量,这些需要量,大都是以幼体动物为试验对象所求得的在试验条件下最佳生长时的最低需要量。
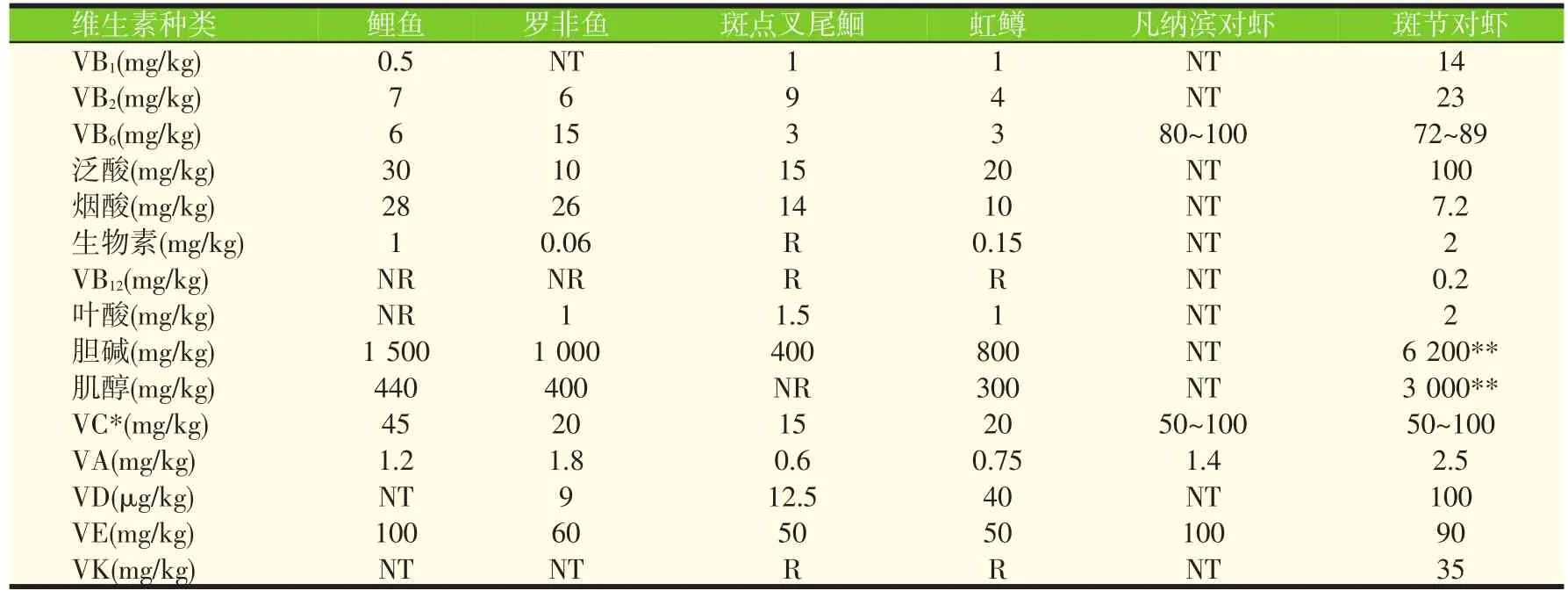
表1 水产动物的维生素需要量(NRC,2011)
目前我国已制订实施了30余项水产饲料行业标准或国家标准,但仅有少部分标准对饲料中的维生素添加量进行了推荐,表2列出了草鱼、青鱼、鲤鱼、鲫鱼、罗非鱼、长吻鮠配合饲料标准对于维生素的推荐量。
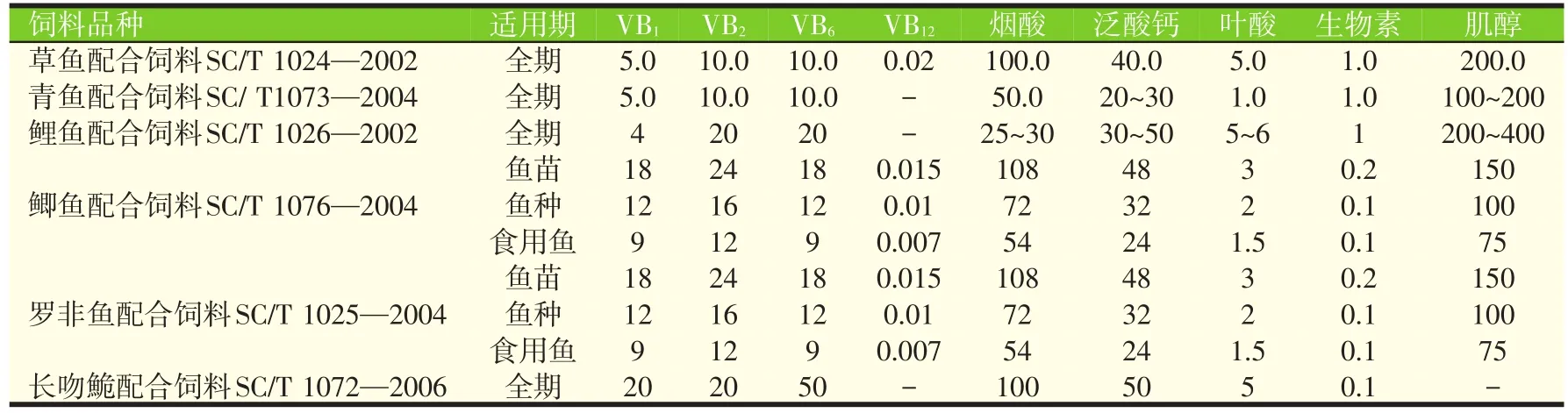
表2 我国水产配合饲料标准对维生素的推荐量(mg/kg)
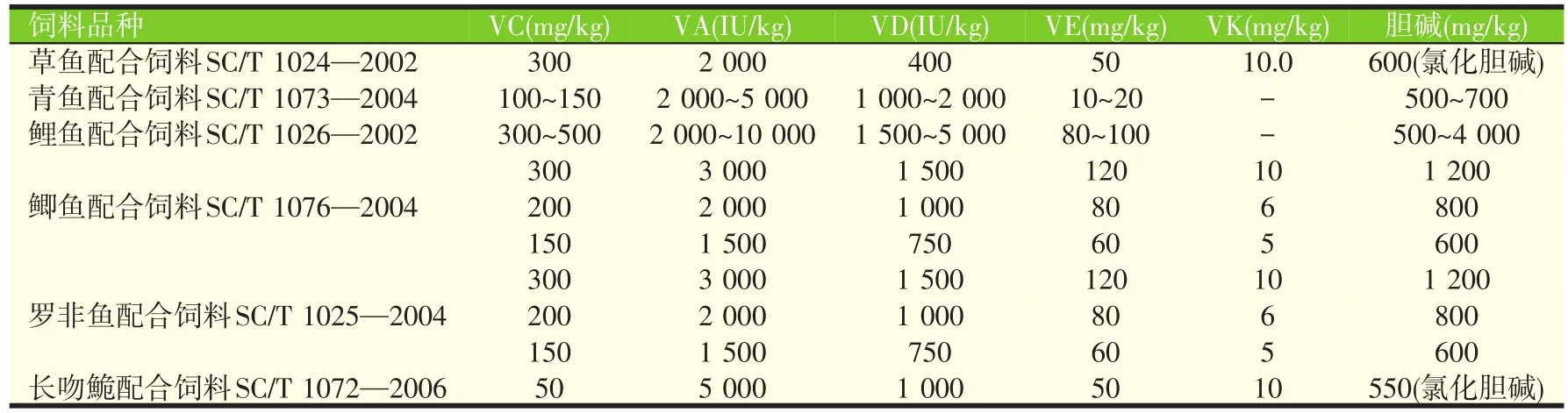
表2 (续) 我国水产配合饲料标准对维生素的推荐量
6 展望
总体来看,维生素的生理功能十分广泛,在动物营养和动物养殖中具有重要的地位和作用。尽管有关维生素营养的研究已进行了多年,但仍然存在许多有待完善和加强的地方。
6.1 维生素的需要量
水产养殖动物的品种繁多,目前已确定维生素需要量的品种十分有限,且所研究的这些品种,大多为幼龄阶段,而其他阶段的维生素需要量并不清楚,今后应加强不同阶段的维生素需要量研究。
6.2 环境对维生素需要量的影响
目前所确定的维生素需要量,多是在实验室条件下,在适宜的密度、温度及良好水质条件下确定的需要量,而在养殖生产中,养殖密度大,水质条件较差,各种环境因子处于较大的波动状态,这些都会对养殖动物产生应激,并影响维生素的需要量。
6.3 加工对维生素保留率的影响
饲料加工中的温度、湿度、压力,特别是膨化加工中的高温高压,会对维生素产生较大的破坏,另外,贮存过程中,也会产生维生素的损失。因此,饲料产品,以及最终为动物所摄食的维生素含量,与配方设计值和生产时的添加值存在较大的差异。有必要对各种加工、贮存条件下维生素的损失进行测定研究,以确保养殖动物能摄入充足的维生素。
6.4 不同生产目的下的需要量
传统养殖,以动物的快速生长为目的,但随着养殖业和社会需求的发展,快速生长并不是唯一目的和评判标准,最佳健康状态、最佳形体、最佳繁殖性能和最佳肉质时对不同维生素的需要量,与快速生长时的需要量并不一致,这些都有待于今后的进一步研究。
猜你喜欢
中国饲料(2021年17期)2021-11-02
中老年保健(2021年5期)2021-08-24
科学养鱼(2020年7期)2020-12-20
保健医苑(2020年1期)2020-07-27
童话世界(2020年8期)2020-06-15
新世纪智能(英语备考)(2019年11期)2020-01-18
湖南饲料(2019年4期)2019-10-17
当代水产(2018年8期)2018-11-02
渔业致富指南(2017年8期)2017-02-02
渔业致富指南(2016年7期)2016-03-28
