
黄石红椿种群Fourier级数分析及动态预测
2016-12-29 06:35:51李新枝
中南林业调查规划 2016年3期
张 梦,尹 茜,刘 仙,李新枝,汪 洋
(湖北生态工程职业技术学院,武汉 430200)
黄石红椿种群Fourier级数分析及动态预测
张 梦,尹 茜,刘 仙,李新枝,汪 洋
(湖北生态工程职业技术学院,武汉 430200)
为研究鄂东地区红椿种群数量动态变化,及其生命周期中生物学特征与种群动态之间的关系,以林木径级代表年龄结构,分析了龄级结构,对种群进行Fourier级数谱分析,并对种群不同龄级动态进行预测。结果表明: 黄石红椿种群为增长型;T1和T2种群波谱分析表明,2个种群均有2次明显小周期,种群天然更新存在着周期性,且有小周期的多谐波叠加;动态预测表明红椿种群龄级数量在未来12年、24年和36年间,均为前期先升,中期降,后期稳定,种群趋于老化。
黄石;红椿;Fourier级数;动态预测
种群动态是一定时间和空间范围内种群的大小和数量变化规律,是植物个体生存能力与环境相互作用的结果,已成为植物生态学研究的热点之一[1]。复杂的周期现象可以由不同振幅和相应的谐波组成,Fourier谱分析是探讨种群天然分布的波动性和年龄更替过程周期性的数学工具,种群天然更新动态可通过种群不同龄级的株数分布波动来表现[2-4]。种群动态研究可以揭示植物种群发展规律,而Fourier谱分析对于发现濒危种群的小周期和其诱发干扰机制,从而制定相应种群的保护措施具有现实意义。
红椿(ToonaciliataRoem.),楝科落叶大乔木,被列为Ⅱ级国家重点保护野生植物[5]。黄石黄荆山红椿天然种群,作为湖北东部红椿种质保存的“岛屿”,是有助于连接湖北东南部和周边省份各天然红椿种群之间的“通道”或“走廊”,该种群的研究对于减少居留种灭亡的风险和物种保护是有着重要意义的。通过对黄荆山红椿种群设立标准样地调查,探讨种群不同龄级的数量和动态变化规律,对于红椿种群生态学研究具有重要意义。
1 研究区概况
研究区位于湖北黄石黄荆山北麓。黄荆山呈东西走向,属低山丘陵地形区,出露地层的岩性主要为石灰岩[7],土壤为山地黄壤;属亚热带气候,四季分明,雨量充沛,年均降雨量1382.6 mm,年最大降雨量2180.1mm,降雨主要集中在3—8月,其降雨量约占全年的81%,年均相对湿度为77%,年均气温19 ℃左右,最高达40 ℃以上,最低达-9 ℃[6],无霜期264 d, 年日照时数1699 h;植被以常绿、落叶阔叶混交林为主。
黄石红椿种群是鄂东至今发现最完整、面积最大的种群,红椿为群落优势种。研究区内主要乔木有梧桐(Firmianasimplex)、枫香(Liquidambarformosana)等;主要灌木及木质藤本有小蜡树(Fraxinusmariesii)、紫藤(Wisteriasinensis)、白花油麻藤(Mucunabirdwoodiana)、胡颓子(Elaeagnuspungens)、扁担杆(Grewiabiloba)等;主要草本和草质藤本植物有风轮菜(Clinopodiumchinense)、庐山石韦(Pyrrosiasheareri)、猪殃殃(Galiumaparine)、九头狮子草(Peristrophejaponica)等。
2 研究方法
2.1 样地设置和调查方法
受环境地形条件限制,在约1 800 m2红椿林设立2个面积为20 m×20 m的标准样地,分别用T1和T2表示,2个样地之间由半荒废的建筑隔开,样地内的红椿全部为成活植株。在每块样地内采用相邻格子法设置5 m×5 m的样方,2样地共设置32个样方,以每个样方为单位调查统计样方内存活的红椿植株,胸径起测点和相关调查模式参照汪洋等[8]采用的方法进行。同时记录环境因子(见表1),对林下乔灌草藤本每木调查统计。

表1 红椿样地环境特征样地海拔/m坡度/(°)坡位坡向/(°)土壤pH值郁闭度T1344~35631中下WN76山地黄壤6 00 8T2342~34510下WS72山地黄壤6 00 7
2.2 龄级划分方法
参照已有研究成果,通过红椿平均胸径与连年生长过程分析,采用“空间替代时间法”,即以立木级结构代替种群年龄结构分析种群动态[9-11],每4年为一个龄级,根据现有种群径级,将该种群划分为12个龄级[12]。湖北全境红椿标准样地天然种群全部按此方法划分龄级,即:幼苗级Ⅰ(DBH<2.5 cm);从第2龄级开始,以胸径5 cm为步长增加一级,幼树级Ⅱ(2.5 cm≤DBH<7.5 cm);Ⅲ级(7.5 cm≤DBH<12.5 cm);Ⅳ(12.5 cm≤DBH<17.5 cm);中树级Ⅴ(17.5≤DBH<22.5 cm);Ⅵ(22.5 cm≤DBH<27.5 cm);大树级Ⅶ(27.5 cm≤DBH<32.5 cm);Ⅷ级(32.5 cm≤DBH<37.5 cm);Ⅸ级(37.5 cm≤DBH<42.5 cm);Ⅹ级(42.5 cm≤DBH<47.5 cm);XI级(47.5 cm≤DBH<52.5 cm);Ⅶ级(DBH≥52.5 cm)。
以龄级为横坐标,不同龄级个体数为纵坐标,绘制红椿种群龄级结构图,并分析不同龄级红椿植株的数量动态特征(见图1)。
2.3 种群Fourier级数分析
国内多位学者利用Fourier级数,来研究不同濒危物种的基波和振幅,以揭示种群发展过程中的小周期和其诱因。Fourier级数正弦波表达和分解中的各个参数[3-4]如下:
Nt=A0+A1sin(ω1t+θ1)+…+Apsin (ωpt+θp)
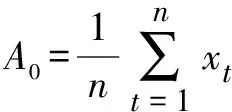
ωk=2πk/T;θk=arctg(ak/bk);
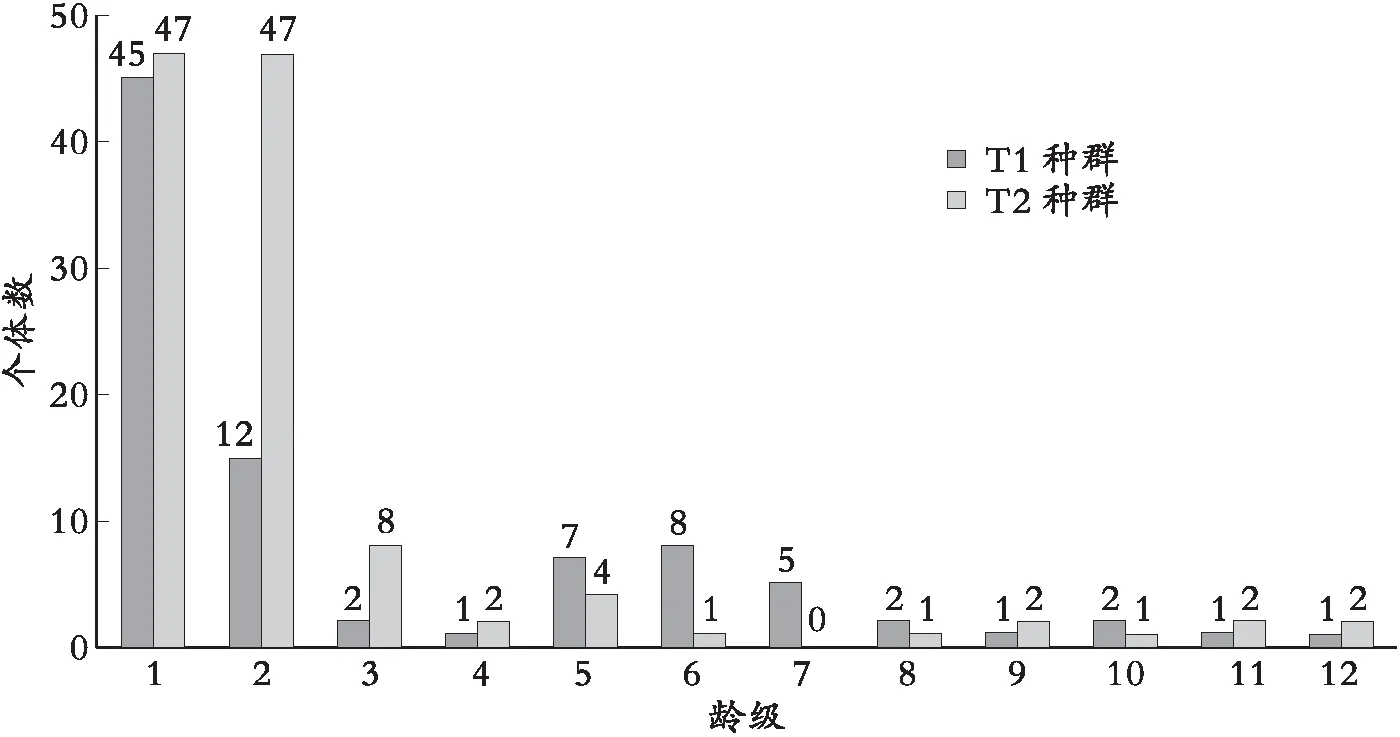
图1 不同红椿种群龄级数量
2.4 种群数量动态时间序列预测

文中数据运用SPSS 22、Excel 2007软件处理。
3 结果与分析
3.1 龄级结构
黄荆山2个红椿样地共16×2个样方,大小存活红椿植株总数分别为T190株,T2117株,共计207株。图1可以看出,T1样地种群第I龄级红椿幼苗级植株为45,占种群全部存活植株的50%,第II龄级为15株,占16.67%。中间经历了Ⅲ和Ⅳ2个龄级数量的锐减,个体数量形成一个谷区,到Ⅴ-Ⅵ龄级时存活数量逐渐上升,其后下降并保持平稳;其中第Ⅳ,Ⅸ,Ⅺ和Ⅻ4个龄级相应数量相同,仅为1株,占比11.11%。T2样地种群第I龄级数量47,占全部存活植株的40.17%,第Ⅱ龄级与第Ⅰ龄级数量相等,第Ⅲ龄级数量仅为8株,占6.84%,第Ⅳ和第Ⅴ2个龄级趋势和T1种群相似,处于逐渐上升,但数量很少,分别为2株和4株,第Ⅴ龄级为0,其后种群趋势与T1相同,保持平稳动态。
由于红椿为强阳性植物,因此从图1龄级分布上可以看出,T1样地种群II龄级损失较大,说明环境条件,尤其是光照条件形成了环境筛,其龄级分布与鄂西南和鄂西北的红椿种群龄级分布十分接近;T2种群处于较为独立的缓坡,周边是一条狭窄山道,光照条件较好,种群第II龄级的数量得以更好保存,其后龄级数量损失极可能与人为干扰或其他环境因素有关。T1和T2种群的数量动态指数分别为55.92%和41.74%[15],2个种群目前都处于稳定和发展阶段。
3.2 红椿种群Fourier级数分析
伍业钢[3]研究红松种群认为,各世代所形成的一定时间间隔的规律性就是红松种群天然更新过程的周期性反映。红椿作为黄石黄荆山红椿群落的优势种,其动态与其他高大乔木一样,形成机制类同。本研究通过红椿存活数量在对应龄级的分布波动来探讨红椿种群的波动特征,以揭示在排除人为干扰后,种群在其生物学因子作用下,如何形成现有的龄级分布。黄石红椿种群的龄级比鄂西北和鄂西南同类种群高[12],因此,种群Fourier谱分析能更准确地表现红椿种群波动特征。
国内大多数学者对不同珍稀植物种群进行过Fourier谱研究,以期解释种群本身固有生物学动因对种群龄级分布的影响[3,14]。在种群的谱分析时,由于研究对象各龄级个体数量相差较大,国内学者在运算时均采用自然对数化处理,即以Xt=ln(Xt+1)代换公式中的Xt[16-17],研究结果与原始龄级分布数量结果间具有较高离散度。因此,本文采用种群固有龄级数量进行Fourier谱分析研究,以期讨论。以龄级结构数量作为波谱分析的级差,共12级,总波序K=N/2=6,基波和不同谐波的振幅Ak值通过Fourier级数公式计算(K=1,2,……,P)(见表2)。Ak值的大小差异反映了各周期作用的大小差异[18]。T1样地种群基波A1=5.521 7,为种群本身所固有,反应种群波动特性。A4=0.585 9,A4的谐波特征表示,红椿种群数量动态除受基波影响外,还显示出明显的小周期波动。第1次小周期反应在12×5/4=15 cm,对应第IV龄级13~16 a(A4),从T1样地种群龄级分布图上,可以看出第IV龄级正是T1样地种群波动谷值,解释了小周期现象的出现;T1样地种群第2次小周期出现在A6,谐波为0.319 3,对应第III龄级,9~12 a(A6),是理论上的最小周期,其形成机制可能来源于光照条件的改变,也可能存在环境条件变化或种内种间竞争。T2样地种群的基波为2.301 1,小于T1的基波,T2小周期波动出现在A3和A5,分别对应17~20 a(A3)和9~12a(A5)。
3.3 种群数量动态序列预测
采用一次移动平均法预测结果如图2和图3。图2可看出,第12 a时,T1样地种群现有存活植株依据种群现有龄级动态,将会由目前的45株(见图1)。
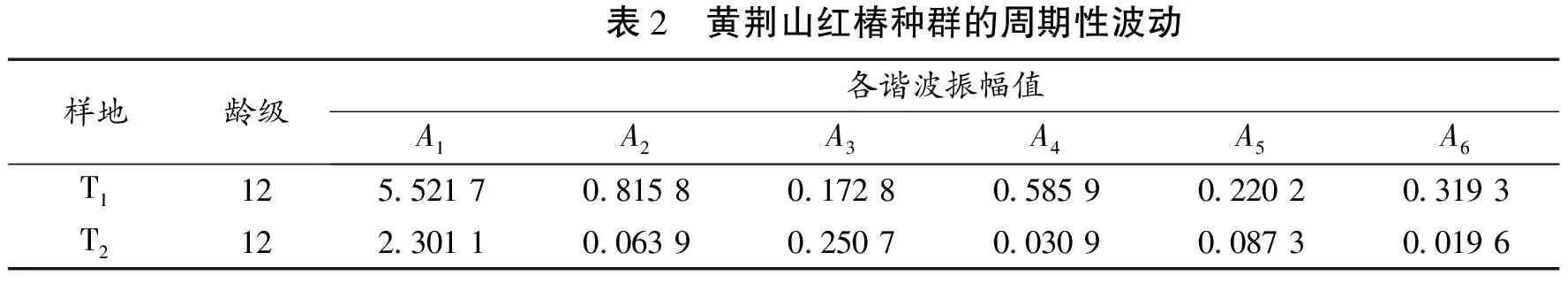
表2 黄荆山红椿种群的周期性波动样地龄级各谐波振幅值A1A2A3A4A5A6T1125 52170 81580 17280 58590 22020 3193T2122 30110 06390 25070 03090 08730 0196
大幅下降到20株左右,种群数量总数在第3龄级会增加,但随年时间推移,到Ⅴ和Ⅵ龄级时数量减少,其后趋于稳定;24 a和36 a时的预测表明,龄级数量动态变化在相应龄级均呈现先增后减。从图3可看出,T2样地种群预测到第12 a时,其相应数量高于T1样地种群同期数量,第V龄级、Ⅵ龄级和Ⅶ龄级的数量高于种群现有水平,其后趋于稳定;T2样地种群第24 a和36 a时动态预测结果表明,种群数量动态发展趋势将与T1样地种群一致。应该注意的是,从Ⅶ和Ⅷ龄级开始,T1样地和T2样地种群均保持平稳态势,成树比例较大,如缺乏更新机制,种群将呈老龄化。
4 结论与讨论
红椿是强阳性植物,除幼苗期外,其生理过程中需要强光照条件。因此,幼苗在发育到幼树的过程中,受到光环境筛的强烈选择。由于缺乏光照,不能维持自身的碳素平衡,植株在进入主层林之前死亡[8],一般情况下,Ⅱ-Ⅴ龄级为红椿种群数量锐减期,这也是湖北红椿天然种群龄级结构的共同特征。而黄荆山T2样地种群幼苗到幼树期的高存活率,可用“林窗机遇”[8,12,17]来解释。由于黄荆山红椿种群具有较为特殊的林缘环境条件,改善的光照条件可以满足其生理需求,解释了T2种群II龄级植株的高存活现象。较高龄级植株进入主林层前,死亡主要来自林冠层下的竞争,植株一旦进入林冠层,即进入稳定发展期。对黄荆山红椿种群研究表明,红椿种群大周期内有小周期的多谐波迭加特征。也有可能同一周期内,个体生长受到前后不同周期波动的影响[8]。小的周期波动可使种群的自我稳定性得以维持与延续[19],小周期波动特征反应了红椿相应龄级的动态特征。

图2 黄荆山T1种群龄数量动态预测
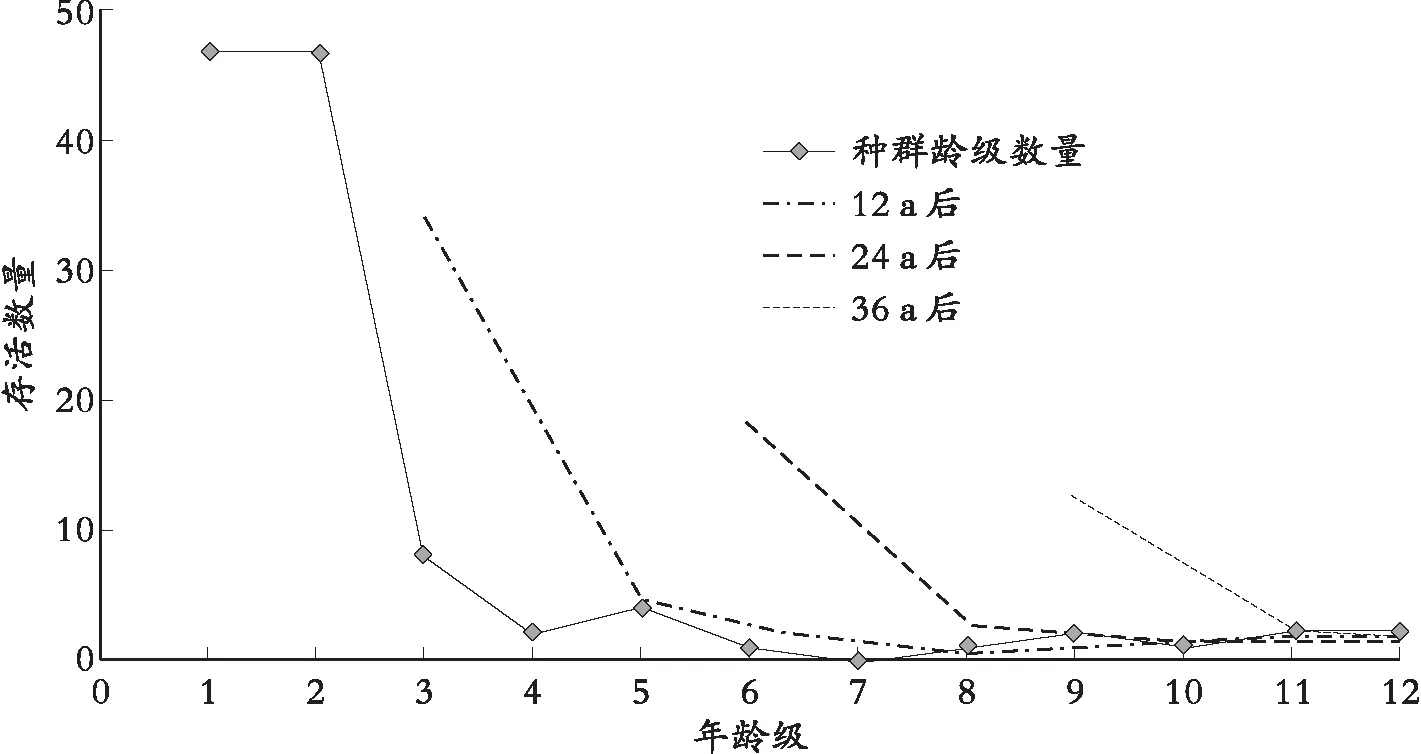
图3 黄荆山T2种群龄数量动态预测
黄荆山红椿幼苗在2个样地中均存在,林缘位置使得红椿幼苗能获得较好的光照。然而,种群自身发展中,环境条件存在巨大的不确定性,如外界环境变化影响光照条件、人为干扰等,会在不定龄级形成新的环境筛,导致种群相应龄级数量结构发生变化。因此,对现存种群保护可以有针对性地进行,如采取适当的“人工正向辅助”措施开辟林窗,以制造有利环境帮助幼树穿过光环境筛,从而促进红椿种群恢复和发展。
[1] 李妍,李登武,韩东辰.贺兰山东坡青海云杉种群动态[J].浙江农林大学学报,2014, 31(1):50-56.
[2] 吴明作,刘玉萃.栓皮栎种群数量动态的谱分析与稳定性[J].生态学杂志,2000,19(4):2-26.
[3] 伍业钢,薛进轩.阔叶红松林红松种群动态的谱分析[J].生态学杂志,1988, 7(1):19-23.
[4] Stewart G H,Rose A B. Thesignificance of history strategies in the developmental history of mixed beech (Nothofagus) forest, New Zealand[J]. Vegetation, 1990,87:101-114.
[5] 法律出版社法规出版中心.国家重点保护野生植物名录:第一批[M].北京:法律出版社,2003.
[6]张起风,石章胜,祝劲,等. 资源枯竭型城市废弃采石场植被恢复与再利用探讨[J]. 林业资源管理,2015(2): 145-149.
[7] 金章利, 刘高鹏, 许文年,等. 黄石黄荆山北麓胡家湾采石场生态修复设计[J]. 中国水土保持,2010(5):61-62.
[8] 汪洋,张敏,陈霞,等. 七姊妹山保护区红椿种群动态与谱分析[J]. 湖北农业科学,2015, 54(22):5632-5636.
[9] 龙汉利,冯毅,向青,等.四川盆周山地红椿生长特性研究[J].四川林业科技,2011,32(3):37-41,68.
[10] 江洪.云杉种群生态学 [M].北京:中国林业出版社,1992:8-26.
[11] 吕海英,王孝安,李进,等.珍稀植物银砂槐中国分布区的种群结构与动态分析[J].西北植物学报,2014, 34(1): 0177-0183.
[12] 汪洋,冷艳芝,苏长江,等.恩施天然红椿种群结构及空间分布格局[J].浙江农林大学学报,2016,33(1): 17-25.
[13] 肖宜安, 何平, 李晓红,等. 濒危植物长柄双花木自然种群数量动态[J]. 植物生态学报,2004,28(2):252-257.
[14] 张文辉, 王延平, 康永祥, 等. 濒危植物太白红杉种群年龄结构及其时间序列预测分析[J]. 生物多样性, 2004, 12(3):361-369.
[15] 周火明,石章胜,何衍明,等. 黄荆山红椿种群动态分析[J]. 湖北林业科技,2016,45(4): 22-26.
[16] 陈远征,马祥庆,冯丽贞,等. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报, 2006, 26(12):4267-4272.
[17] 汪洋,闵水发,江雄波,等.恩施盛家坝红椿天然种群动态与预测[J].福建林业科技,2016,43(2):21-26.
[18] 封磊,洪伟,吴承祯,等.珍稀濒危植物南方铁杉种群动态研究[J]. 武汉植物学研究, 2003,21(5):401-405.
[19] 闫淑君,洪伟,吴承祯,等.丝栗栲种群生命过程及谱分析[J].应用与环境生物学报, 2002, 8(4): 351-355.
FourierSeriesandPredictionAnalysesofToonaciliataPopulationinHuangshi
ZHANG Meng,YIN Xi,LIU Xian,LI Xinzhi,WANG Yang
(Hubei Vocational College of Economical Engineering,Wuhan 430200,Hubei,China)
In order to research the dynamics change, and clarify the association between biological characters and population quality in the life cycle ofToonaciliatain in Huangjingshan Mountain, the diameter classes ofT.ciliataplants were stood for age classes, numeral dynamics of the plants in different age classes were analyzed, spectral analyses and a prediction for future development of the populations were conducted. The research indicated that the populations were in growth form; in the spectral analyses of the 2 populations, the fundamental waves were each overlaid by 2 different harmonic waves with 2 obvious minor cycles, demonstrating there was a periodicity in natural regeneration ofT.ciliatapopulations; in the prediction for upcoming 12, 24, and 36 years, the individual numbers of plants declined first, increased moderately then and stabilized thereafter, the populations was tended to over aging.
Huangshi;Toonaciliata;Fourier series;prediction analysis
2016-09-20
湖北省科技厅公益性研究项目(402012DBA40001);湖北省教育厅科研项目(B20160555)。
张梦(1984-),女,湖北巴东人,主要研究方向:园林技术。
李新枝(1984-),女,讲师,主要研究方向:园林技术。
Q948.15+7
A
1003-6075(2016)03-0044-05
10.16166/j.cnki.cn43-1095.2016.03.010
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
东北林业大学学报(2022年7期)2022-08-04 01:48:02
东坡赤壁诗词(2020年2期)2020-06-04 15:44:10
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
乡村科技(2018年17期)2018-08-16 02:27:50
环球时报(2018-08-03)2018-08-03 08:11:48
湖北林业科技(2016年5期)2016-12-02 10:23:28
儿童故事画报(2016年2期)2016-04-18 03:32:25
剑南文学(2015年2期)2015-02-28 01:15:20
