
跑台运动对高脂饮食诱导的肥胖大鼠骨骼肌肌联素表达的影响
2016-12-09 08:05张靓董泽源马谨陈雪飞
中国运动医学杂志 2016年6期
张靓 董泽源 马谨 陈雪飞
北京师范大学体育与运动学院(北京 100875)
跑台运动对高脂饮食诱导的肥胖大鼠骨骼肌肌联素表达的影响
张靓 董泽源 马谨 陈雪飞
北京师范大学体育与运动学院(北京 100875)
目的:观察跑台运动对高脂饮食诱导的肥胖大鼠骨骼肌肌联素(myonectin)表达的影响。方法:5周龄健康雄性SD大鼠随机分成对照组和高脂喂养组,分别进行普通饲料和高脂饲料喂养,12周后筛选肥胖大鼠16只,随机分为高脂安静组和高脂运动组。跑台训练持续10周,每周5次,每次60 min,跑速26 m/min。测定大鼠血脂及血浆自由脂肪酸水平,采用real-time PCR方法测定比目鱼肌、腓肠肌肌联素mRNA表达,比目鱼肌脂肪酸转位酶(fatty acid translocase,FAT/CD36)的mRNA表达。结果:与对照组相比,高脂饮食喂养大鼠体重和体脂显著增加(P<0.01),总甘油三酯、总胆固醇和低密度脂蛋白水平显著升高,高密度脂蛋白水平显著降低(P<0.05),血自由脂肪酸(free fatty acid,FFA)水平显著升高(P<0.05)。比目鱼肌和腓肠肌肌联素mRNA表达水平显著升高(P<0.05),比目鱼肌CD36表达显著上调(P<0.05)。与高脂饮食组大鼠相比,10周跑台运动大鼠体重显著降低(P<0.01),血脂改善,血FFA水平显著降低(P<0. 05)。此外,比目鱼肌和腓肠肌肌联素mRNA表达显著下调(P<0.05),比目鱼肌CD36表达显著降低(P<0. 05)。相关性分析发现,骨骼肌肌联素水平与血脂和血糖水平呈显著正相关(P<0.05)。结论:骨骼肌肌联素的表达与血脂、血糖水平显著相关,运动促进了血脂和血糖水平的降低,进而间接下调了骨骼肌肌联素的表达,提示肌联素可能不参与运动引起的自由脂肪酸水平的降低。
跑台运动;肌联素;自由脂肪酸;肥胖;骨骼肌
肌联素(myonectin)是新发现的肌肉因子[1],为补体C1q/肿瘤坏死因子相关蛋白(C1q/tumor necrosis factorrelated protein,CTRP)家族成员,即CTRP15。肌联素特异性地在骨骼肌高表达,并以内分泌的方式作用于外周,促进多种组织对自由脂肪酸(free fatty acid,FFA)的摄入,降低血清FFA水平。以每克体重5 μg外源性给予重组肌联素,发现注射后2小时,小鼠血清FFA显著降低,3小时后血清FFA下降30%,达到最低点[1]。此外,在离体培养的3T3-L1脂肪细胞及H4IIE肝细胞,5 μg/mL的肌联素处理显著促进了长链脂肪酸棕榈酸酯的摄入[1]。在体和离体实验均提示肌联素可通过促进脂肪酸摄入降低血清脂肪酸水平。根据以往文献,有氧运动可以有效地降低血浆FFA水平[2,3],促进外周组织对FFA的摄入及氧化,改善多种代谢性疾病的症状,但运动发挥上述效应的作用机制尚不明晰。肥胖时,有氧运动是否是通过肌联素来介导其降低血清FFA的作用,目前尚不清楚。本研究以高脂饮食喂养诱导的肥胖大鼠为模型,观察10周的跑台运动后,血清脂肪酸及骨骼肌肌联素mRNA表达的变化,探讨肌联素在有氧运动降低血清脂肪酸中的作用。
1 材料与方法
1.1 实验动物与材料
本实验所有动物饲养和实验程序按中国动物管理条例(中华人民共和国卫生部2001年第55号文件)批准的动物实验标准进行。5周龄雄性SD大鼠由北京大学医学部动物中心提供,实验动物生产许可证号:SCXK(京)2011-0012。动物实验于北京大学医学部动物中心完成,动物使用许可证号:SYXK(京)2011-0039。常规条件下饲养,室内温度为22±2℃,湿度为40%~60%,自由进食和饮水。血脂、血葡萄糖检测试剂盒由中生北控股份有限公司提供,血胰岛素检测放免试剂盒及血清FFA测定试剂盒由北京华英生物技术研究所提供。高脂饲料购自北京科澳协力饲料有限公司,其中每1000 g高脂饲料由800 g基础饲料加200 g脂肪(170 g猪油+30 g玉米油)与1%胆固醇混合而成,脂肪供能占40%[4]。实验中所用其他试剂均为市售分析纯试剂。
1.2 动物分组与运动模型建立
5周龄健康雄性SD大鼠48只,随机分成对照组(n=16)和高脂喂养(high fat diet,HFD)组(n=32),分别进行普通饲料和高脂饲料喂养,12周后,以体重≥对照组体重均值加1.4倍标准差为标准筛选肥胖大鼠16只。将对照组大鼠随机分为对照安静组(Con,n=8)和对照运动组(Con+Exe,n=8);高脂喂养肥胖大鼠随机分为高脂安静组(HFD,n=8)和高脂运动组(HFD+Exe,n=8),继续高脂饮食饲养。运动组采用跑台运动,经1周适应,跑台速度逐渐加快至26米/分钟[5],跑台坡度为0,运动时间逐渐延长至60 min。正式跑台训练持续10周,每周5次,每次60 min,跑速26米/分钟。每次训练在下午3点至6点间进行,不使用声、光、电等刺激手段。
1.3 取材及组织准备
大鼠末次运动结束后,禁食过夜,自由饮水,腹腔注射乌拉坦(1 g/kg)麻醉。经腹主动脉取血,用于各生化指标的测定。动物处死后,分离比目鱼肌和腓肠肌,用锡箔纸包裹、标记后于-80℃保存,进行下述测定。
1.4 Real-time PCR测定比目鱼肌、腓肠肌肌联素及脂肪酸转位酶(fatty acid translocase,FAT/CD36)的mRNA水平
采用Trizol一步法提取骨骼肌总RNA,Promega逆转录系统(Promega,Madison,WI,USA)进行逆转录。real-time PCR反应体积共20 μL:SuperRealPreMix Plus(SYBR Green)体系(TianGen Biotech,Beijing)16.8 μl,10 μmol/L的上下游引物各0.6 μl,cDNA模板2 μl。β-actin的上游引物:5’-GAG ACC TTC AAC ACC CCA GCC-3’;下游引物为:5’-TCG GGG CAT CGG AACCGCTCA-3’;肌联素的上游引物:GGCACAGTTTCTCCTTCTGG;下游引物为:AGCCATCTTCATCACCAAGC;FAT/CD36的上游引物:ATTTGTTCTTCCAGCCAACG;下游引物为:CCCAGTCTCATTTAGCCACA。所有引物均由生工生物工程(上海)股份有限公司合成。经95℃7 min变性后,肌联素进行95℃30 s,64℃30 s,72℃40 s,热循环40次。FAT/CD36进行95℃30 s,60℃30 s,72℃40 s,热循环40次。Real-time PCR于Mx3000P多通道实时定量PCR仪(Stratagene,La Jolla,CA,USA)上进行,以β-actin作为内参。
1.5 统计方法
实验结果以均数±标准差(means±SD)表示。数据采用单因素方差分析处理,P<0.05为有显著性差异。
2 结果
2.1 高脂饮食及跑台运动对大鼠体重、血糖、血胰岛素、血脂和血FFA的影响
与正常饮食组大鼠相比,高脂饮食诱导大鼠出现显著的肥胖,代谢紊乱,表现为体重显著增加(P<0.01)(图1),血糖(P<0.05)、血胰岛素水平显著上调(P<0. 01),血总胆固醇(P<0.05)、总甘油三酯(P<0.01)、低密度脂蛋白胆固醇(P<0.01)显著升高,高密度脂蛋白胆固醇显著降低(P<0.01)(图2)。此外,血浆FFA水平在高脂组大鼠也显著升高(P<0.05)(图2)。10周的跑台运动显著逆转了高脂饮食诱导的上述变化,体重显著降低(P<0.01)(图1),血糖和血胰岛素水平显著下降,脂代谢各参数也都回归到正常水平(图2),提示运动改善了肥胖大鼠的糖、脂代谢。同时,运动还显著降低肥胖大鼠血FFA水平(图2)。

图1 高脂饮食喂养及运动对大鼠体重的影响
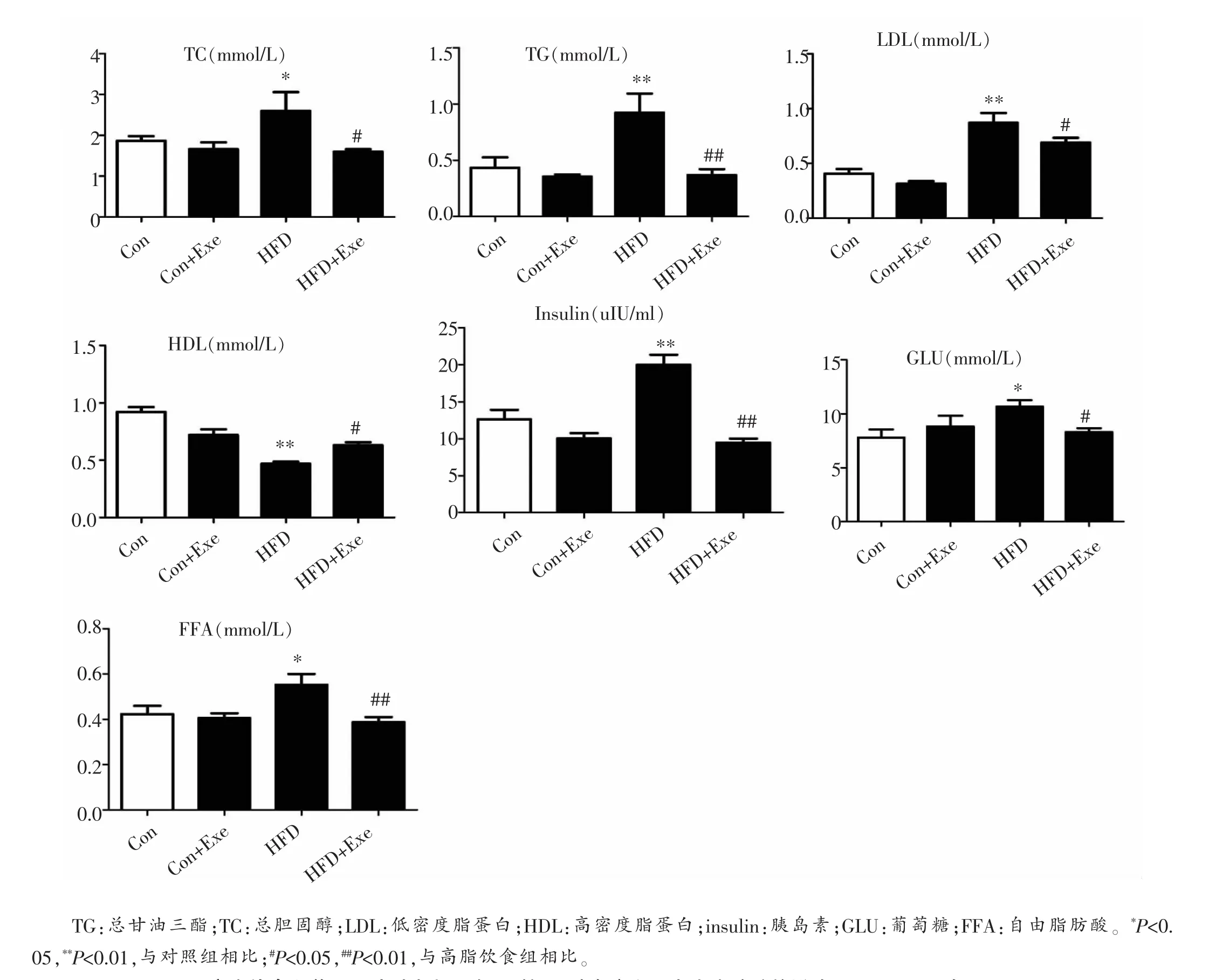
图2 高脂饮食喂养及运动对大鼠血脂、血糖、血胰岛素和血自由脂肪酸的影响(means±SD,每组n=8)。
2.2 高脂饮食及跑台运动对大鼠比目鱼肌和腓肠肌肌联素mRNA表达的影响
与对照组相比,单纯高脂饮食组大鼠比目鱼肌和腓肠肌的肌联素mRNA表达水平显著升高(P<0.05)(图4)。跑台训练显著降低了肥胖大鼠比目鱼肌和腓肠肌肌联素mRNA的表达(P<0.05)(图4)。正常饮食运动组骨骼肌肌联素的表达水平无显著变化。我们的结果与Seldin等[1]的报道不一致,在其研究中发现运动可以显著上调小鼠骨骼肌中肌联素的表达,而高脂喂养则对其有显著的抑制作用。而Peterson等[6]在肥胖的Zucker大鼠观察到膈肌中肌联素的表达显著升高,而运动则显著下调其肌联素的表达,支持了本文的研究结果。我们将在讨论部分对结果的差异进行分析。
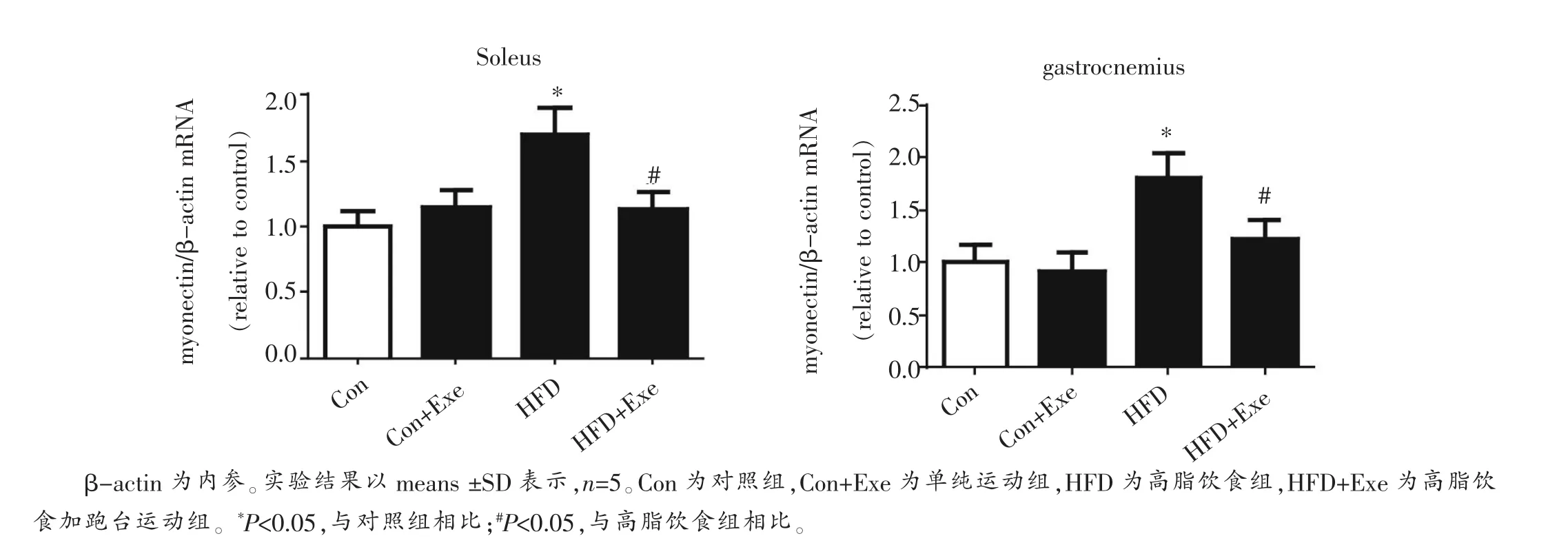
图3 高脂饮食喂养及运动对大鼠比目鱼肌和腓肠肌肌联素mRNA表达的影响
2.3 高脂饮食及跑台运动对大鼠比目鱼肌脂肪酸转位酶(fatty acid translocase,FAT/CD36)mRNA表达的影响
FAT/CD36是骨骼肌主要的脂肪酸转运体之一,在FAT/CD36基因敲除的小鼠[7],脂肪酸摄入显著减少,而FAT/CD36基因过表达时[8],骨骼肌对脂肪酸的利用显著增加,表明FAT/CD36在骨骼肌脂肪酸摄入中的关键作用。有研究表明,肌联素通过刺激FAT/CD36的表达来促进脂肪酸的摄入。在本实验中,长期的高脂饮食大鼠比目鱼肌FAT/CD36的表达显著升高(P<0.05)(图5),而跑台训练则显著下调了肥胖大鼠FAT/CD36的表达(P<0.05)(图5),该结果与以往文献报道一致[9]。 2.4血甘油三酯、血糖及血游离脂肪酸与骨骼肌肌联素mRNA表达的相关性分析

图4 高脂饮食喂养及运动对大鼠比目鱼肌脂肪酸转位酶CD36 mRNA表达的影响
血甘油三酯、血糖及血游离脂肪酸与比目鱼肌、腓肠肌中肌联素mRNA表达的相关性分析结果表明(表1),血甘油三酯、血糖及血游离脂肪酸与比目鱼肌、腓肠肌中肌联素mRNA表达呈显著正相关(P<0.05)。
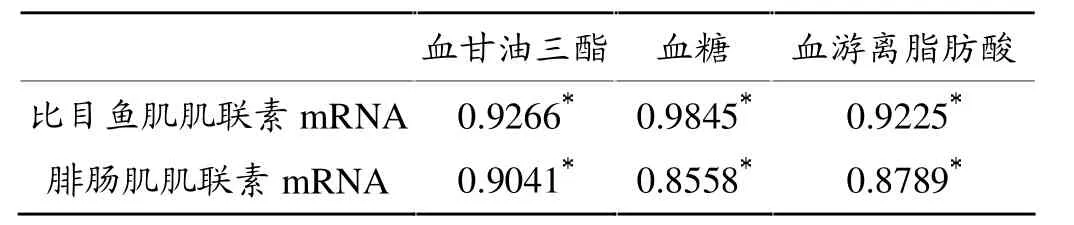
表1 血甘油三酯、血糖及血游离脂肪酸与比目鱼肌、腓肠肌中肌联素mRNA表达水平的相关系数
3 讨论
3.1 运动对血游离脂肪酸的调节作用
自由脂肪酸(free fatty acid,FFA),又称为非酯化脂肪酸,脂肪组织中的甘油三酯被动员后水解产生FFA,被肌肉、肝脏等外周组织摄取利用,是人体重要的能量底物。FFA不溶于水,在血液中以血浆白蛋白为载体进行运输,只有不到0.1%的FFA是真正“游离”的。血浆白蛋白有7个与FFA结合的位点,其中3个位点的亲和力高,而其余四个位点的亲和力较低。在正常生理状况下,血清FFA与白蛋白的比值大概在0.1~2,但在某些时候,如运动、肥胖、禁食、胰岛素抵抗等特殊状
态下,FFA与白蛋白的比值显著升高,其比值甚至会高达6左右[10-11]。
运动和禁食等生理状况下,血FFA水平的升高通常是一过性的,随着运动或禁食的结束,FFA水平随即恢复。但在肥胖、胰岛素抵抗等病理状态下,血浆FFA水平持续升高,是正常体重人群的两倍左右[12]。升高的血浆FFA,远远超过外周组织的氧化能力[13],给肝脏、肌肉等器官带来极大的代谢压力。此外,循环中超载的FFA可与多种膜受体和核受体结合,如模式识别受体(pattern recognition receptors,PRRs)、Toll样受体(Tolllike receptors,TLRs)、NOD样受体(nucleotide-binding and oligomerization domain-like receptors,NLRs)及自由脂肪酸受体(FFA receptors,FFARs)等[14],诱导活性氧生成、内质网应激、细胞凋亡及炎症发生[15],导致胰岛素抵抗、脂代谢紊乱、II型糖尿病、心血管疾病及多种代谢性疾病的发生[16]。本文结果发现,22周的高脂饮食喂养,大鼠出现显著的肥胖和糖脂代谢紊乱,血浆FFA水平显著升高。
大量文献证实,运动能有效地降低血FFA水平。本文中,10周的跑台运动后,肥胖大鼠血浆FFA水平显著降低,此外体重、糖脂代谢指标均显著改善。运动降低血FFA的机制主要与FFA的摄取及氧化增加有关。以骨骼肌为例,不管是在体还是离体研究,均表明肌肉收缩能显著增加脂肪酸的摄入水平。骨骼肌细胞膜存在三类脂肪酸转运体,分别是脂肪酸转位酶(FAT/ CD36)、胞膜脂肪酸结合蛋白(plasma membrane fatty acid-binding protein,FABPpm)和脂肪酸转运蛋白(fatty acid transport protein,FATP),它们负责骨骼肌脂肪酸的摄入[17]。研究发现,运动可促进它们在骨骼肌的表达和/或膜转位,从而加速脂肪酸的摄入。但运动调节脂肪酸转运体的表达和膜转位的机制尚不清楚。
3.2 肌联素在运动降低脂肪酸中的作用
蛋白质组学研究发现,人及小鼠的骨骼肌细胞可合成大量的可分泌蛋白,人骨骼肌产生大约250种,小鼠骨骼肌产生的则更多,约600种,它们通过自分泌、旁分泌及内分泌的形式发挥功能[18,19]。已将骨骼肌合成、分泌的细胞因子和活性多肽命名为肌肉因子(myokines)[20]。运动时肌肉收缩诱导肌肉因子的合成和分泌改变,已成为运动调节代谢的重要机制之一。
肌联素是2012年利用脂联素的cDNA序列询问人及小鼠表达序列标签(expressed sequence tags,ESTs)和基因组数据库所得到的一个高度保守的蛋白家族成员之一[1]。由于对脂联素生物学功能的熟知,肌联素的发现也让研究者们充满了期待。Seldin等[1]研究表明,肌联素是骨骼肌特异表达的肌肉因子,且在慢肌中的表达量显著高于快肌。肌联素对FFA代谢有显著的调节作用,外源性给予小鼠肌联素,显著降低了血清FFA的水平。离体实验证实,肌联素处理能显著促进肝脏和脂肪细胞对脂肪酸的摄取,但对脂肪酸的氧化无明显作用。进一步研究发现,肌联素通过上调肝脏和脂肪细胞脂肪酸转运蛋白CD36、FATP1、Fabp4的转录水平,促进脂肪酸摄入。据此,我们推测运动可能通过上调骨骼肌肌联素的表达来促进骨骼肌脂肪酸摄入。
但本实验的结果却不支持我们的假设。在正常饮食运动组,比目鱼肌和腓肠肌肌联素mRNA的表达与不运动组相比无显著差异,提示运动不影响正常大鼠骨骼肌肌联素的基因表达水平。然而,长期的高脂饮食却显著上调了骨骼肌中肌联素的基因表达水平,而运动显著下调了肥胖大鼠骨骼肌肌联素的基因表达。结果表明肌联素可能不参与运动对血清FFA的调节作用。在测定肌联素的下游分子CD36的mRNA表达后发现,骨骼肌CD36表达的变化趋势与肌联素相吻合,即肥胖大鼠CD36表达显著升高,运动则回调肥胖大鼠CD36的表达。但高脂和运动对CD36膜转位的调节作用还需进一步研究。
3.3 运动和肥胖对肌联素表达的调节
本实验的上述结果与Seldin等[1]的报道不一致,为此我们对肌联素的表达调控因素进行了分析,发现肌联素对机体的营养状况尤其敏感,在禁食和再饲2小时后,比目鱼肌肌联素的表达上调近百倍。为了辨别是碳水化合物还是脂肪增加了再饲小鼠肌联素水平,再饲时分别选择葡萄糖和脂肪乳灌胃,发现两者均显著增加血浆中肌联素的水平,没有显著差异[1]。在小鼠C2C12细胞分别进行25 mM的葡萄糖和1μM的棕榈酸酯培养,发现肌联素的表达均显著上调[1],提示机体的血糖和血脂可直接诱导骨骼肌肌联素的表达上调。本实验也发现,骨骼肌肌联素的表达与大鼠的血脂和血糖水平成显著正相关,跑台训练降低了肥胖大鼠的血脂和血糖水平,间接下调了骨骼肌肌联素的表达。本研究结果再次证实,机体的营养状况对肌联素的表达起着决定性作用。此外,本实验所有动物在处死之前均空腹过夜,因而禁食对肌联素强大的抑制作用有可能掩盖了运动的效应。
Peterson等[6]在肥胖的Zucker大鼠上同样观察到膈肌肌联素的表达显著上调,而运动则明显下调了肌联素的表达,与本文结果一致。由于瘦素对肌联素的表达有显著的刺激作用[21],因而肥胖时循环中瘦素水平的升高也可能是诱导骨骼肌肌联素表达升高的原因之一。
4 总结
10周的跑台运动显著降低的肥胖大鼠血清FFA
水平,下调了肥胖诱导的骨骼肌肌联素的表达升高,提示肌联素可能不参与运动降低血FFA水平的调节。
[1]SeldinMM,PetersonJM,ByerlyMS,etal.Myonectin(CTRP15),a novel myokine that links skeletal muscle to systemic lipid homeostasis.J Biol Chem,2012,287(15):11968-11980.
[2]Katsanos CS.Lipid-induced insulin resistance in the liver:role of exercise.Sports Med,2004,34(14):955-965.
[3]Jensen MD.Fate of fatty acids at rest and during exercise:regulatory mechanisms.Acta Physiol Scand,2003,178(4):385-390.
[4]Kim YJ,Park T.Genes are differentially expressed in the epididymal fat of rats rendered obese by a high-fat diet.Nutr Res,2008,28(6):414-422.
[5]Gauthier MS,Couturier K,Charbonneau A,et al.Effects of introducing physical training in the course of a 16-week high-fat diet regimen on hepatic steatosis,adipose tissue fat accumulation,and plasma lipid profile.Int J Obes Relat Metab Disord,2004,28(8):1064-1071.
[6]Peterson JM,Mart R,Bond CE.Effect of obesity and exercise on the expression of the novel myokines,Myonectin and Fibronectin type III domain containing 5.Peer J,2014,30,2:e605.
[7]Febbraio M,Abumrad NA,Hajjar DP,et al.A null mutation in murine CD36 reveals an important role in fatty acid and lipoprotein metabolism.J Biol Chem,1999,274(27):19055-19062.
[8]IbrahimiA,BonenA,BlinnWD,etal.Muscle-specific overexpression of FAT/CD36 enhances fatty acid oxidation by contracting muscle,reduces plasma triglycerides and fatty acids,and increases plasma glucose and insulin.J Biol Chem,1999,274(38):26761-26766.
[9]SmithAC,MullenKL,JunkinKA,etal.Metforminand exercise reduce muscle FAT/CD36 and lipid accumulation andblunttheprogressionofhigh-fatdiet-induced hyperglycemia.Am J Physiol Endocrinol Metab,2007,293(1):E172-181.
[10]Fujiwara S,Amisaki T.Fatty acid binding to serum albumin: molecular simulation approaches.Biochim Biophys Acta,2013,1830(12):5427-5434.
[11]Simard JR,Zunszain PA,Ha CE,et al.Locating high-affinity fatty acid-binding sites on albumin by x-ray crystallography and NMR spectroscopy.Proc Natl Acad Sci U S A,2005,102(50):17958-17963.
[12]EbbertJO,JensenMD.Fatdepots,freefattyacids,and dyslipidemia.Nutrients,2013,5(2):498-508.
[13]Mittendorfer B.Origins of metabolic complications in obesity:adipose tissue and free fatty acid trafficking.Curr Opin Clin Nutr Metab Care,2011,14(6):535-541.
[14]Ringseis R,Eder K,Mooren FC,et al.Metabolic signals and innate immune activation in obesity and exercise.Exerc Immunol Rev,2015,21:58-68.
[15]Mizuno Y,Harada E,Katoh D,et al.Cardiac production of B-type natriuretic peptide is inversely related to the plasma level of free fatty acids in obese individuals-possible involvement of the insulin resistance.Endocr J,2013,60(1):87-95.
[16]Boden G.Obesity,insulin resistance and free fatty acids.Curr Opin Endocrinol Diabetes Obes,2011,18(2):139-143.
[17]PelsersMM,StellingwerffT,vanLoonLJ.Theroleof membranefatty-acidtransportersinregulatingskeletal muscle substrate use during exercise.Sports Med,2008,38(5):387-399.
[18]Henningsen J,Rigbolt KT,Blagoev B,et al.Dynamics of the skeletal muscle secretome during myoblast differentiation.Mol Cell Proteomics,2010,9(11):2482-2496.
[19]NorheimF,RaastadT,ThiedeB,etal.Proteomic identification of secreted proteins from human skeletal muscle cells and expression in response to strength training.Am J Physiol Endocrinol Metab,2011,301(5):E1013-E1021.
[20]Pedersen BK,Febbraio MA.Muscles,exercise and obesity:skeletal muscle as a secretory organ.Nat Rev Endocrinol,2012,8(8):457-465.
[21]Rodríguez A,Becerril S,Méndez-Giménez L,et al.Leptin administration activates irisin-induced myogenesis via nitric oxide-dependentmechanisms,butreducesitseffecton subcutaneous fat browning in mice.Int J Obes(Lond),2015,39(3):397-407.
Effects of Treadmill Running on the Myonectin in Skeletal Muscle of Obese Rats
Zhang Jing,Dong Zeyuan,Ma Jin,Chen Xuefei
School of P.E.and Sports Science,Beijing Normal University,Beijing,China 100875 Corresponding Author:Zhang Jing,Email:zhangjing@bnu.edu.cn
Objective To observe the alteration of the myonectin expression in skeletal muscle of obese rats induced by high-fat diet.Methods 5-week-old male SD rats were randomly divided into control group and high-fat diet group.16 obese rats after 12-week feeding with high-fat diet were recruited and then randomly divided into sedentary group and treadmill running group.The rats in treadmill running group underwent 60-minute treadmill running once per day at the intensity of 26 m/min,5 days per week for 10 weeks.The mRNA expression of myonectin and fatty acid translocase(FAT/CD36)level were detected by real-time PCR.Results The body weight and body fat increased(P<0.01),levels of triglyceride,cholesterol and low density lipoprotein elevated(P<0.05),high density lipoprotein decreased(P<0.05),level of free fatty acid(FFA)increased(P<0. 05),and the mRNA levels of myonectin and CD36 in soleus and gastrocnemius increased(P<0.05)in obese rats as compared with the control group.The body weight decreased(P<0.01),lipid profile improved,serum FFA content decreased(P<0.05)and the mRNA level of myonectin and CD36 in muscle decreased(P<0.05)in treadmill running group after 10-week exercise as compared with sedentary obese group.There was positive correlation between the expression of myonectin mRNA and serum triglyceride,fasting blood sugar and FFA(P<0.05).Conclusion Results showed that exercise improved blood sugar and lipid metabolism,and thus
treadmill running,myonectin,free fatty acid,skeletal muscle,obesity
2015.12.18
国家自然科学基金(31471137)
张靓,Email:zhangjing@bnu.edu.cn
downregulated the myonectin mRNA level in muscle of obese rats indirectly,suggesting that myonectin might not involve in the exercise-induced decreased level of FFA.
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
医学综述(2021年16期)2021-12-01
中华养生保健(2020年5期)2020-11-16
中国生殖健康(2018年1期)2018-11-06
中成药(2018年10期)2018-10-26
天然产物研究与开发(2018年2期)2018-04-04
心脑血管病防治(2015年3期)2016-01-15
医学研究杂志(2015年12期)2015-06-10
军事体育学报(2015年2期)2015-02-27
江苏农业科学(2014年4期)2014-07-11
