
石刁柏Ty1—copia反转座子逆转录酶序列克隆及异质性分析
2016-12-08 18:58石晓卫李书粉邓传良卢龙斗
湖北农业科学 2016年18期
石晓卫++李书粉++邓传良++卢龙斗++高武军

摘要:对石刁柏(Asparagus officinalis)品种UC309及TD818的两个反转座子逆转录酶序列进行了PCR扩增并对其在基因组中的异质性进行了分析。结果表明,s101366和s107518两个转座子序列均属于Ty1-copia反转座子,是Ty1-copia反转座酶的部分保守区序列,其长度分别为636和653 bp。对随机获得的两个反转座子测序表明两个反转座子存在一定的异质性,其突变主要是由于发生了碱基置换,其中碱基转换分别占到80.4%和80.0%,T?圮C和G?圮A转换的比例差异不显著。通过对两个石刁柏品种 (TD818和UC309) 中获得的序列聚类分析发现,s101366和s107518均未表现出性别间、品种间及近缘种间的保守性,甚至表现出了种内异质性大于种间的异质性特征。这说明了两个反转座子处于高度变异的状态,是产生染色体不稳定的重要因素之一。
关键词:石刁柏(Asparagus officinalis);Ty1-copia反转座子;逆转录酶;序列分析;异质性
中图分类号:Q754 文献标识码:A 文章编号:0439-8114(2016)18-4819-05
DOI:10.14088/j.cnki.issn0439-8114.2016.18.048
显花植物中,包括6个双子叶和5个单子叶亚纲在内的75%的科,都存在雌雄单性或者雌雄异株种[1,2],大约38%(167个)的科和7.1%(959个)的属中有6%(14 620种)的植物是雌雄异株植物[2],如石刁柏(Asparagus officianalis)、啤酒花(Humulus lupulus)、番木瓜(Carica papaya)、菠菜(Spinacia oleracea)等[3]。研究表明,雌雄异株植物的性别决定方式多种多样,其中性染色体决定是最为常见的性别决定方式之一[2]。截至2011年,在陆生开花植物中已经发现有15个科21个属40种植物具有同型或者异型性染色体(多为XY或者ZW系统),这些性染色体(XY或ZW)最初起源于一对常染色体[3,4]。但是对于早期性染色体如何从常染色体逐步演化为异型或者同型性染色体的机制仍不清楚。
反转座子是一类广泛存在于真核细胞中可移动的DNA元件,具有高拷贝数、高异质性和插入位点专一性等特点[5],又称反转录转座子、逆元件。研究表明,反转座子在植物的基因组中广泛存在且占有较大的比例,如拟南芥和水稻基因组中反转座子分别占了基因组的10%[6]和22%[7],小麦基因组中反转座子成分甚至达到49.07%[8]。Ty1-copia类反转座子是广泛存在于高等植物基因组中的一类反式元件[9],且能够通过整合过程中的重组和染色体重排来影响基因组和染色体组的稳定[10,11]。因此,Ty1-copia类反转座子的这种诱发染色体组的不稳定作用可能是导致性染色体起源的因素之一[12]。研究表明,番木瓜Yh染色体雄性特异区的转座子主要为Ty1-copia和Ty3-gypsy,且X染色体上的Ty3-gypsy反转座子是常染色体上的两倍[13],进一步暗示了Ty1-copia类反转座子在植物性染色体演化中可能具有重要的作用。
石刁柏是一种较为严格的雌雄异株植物,体细胞染色体数为2n=20,具有一对同型的性染色体[14]。研究发现石刁柏染色体组存在异型的常染色体对、B染色体数目不固定等特征[15,16],说明石刁柏的染色体组处于不稳定的状态,但是尚未有报道证明这种不稳定状态产生的原因。因此,本研究在利用高通量测序技术对石刁柏基因组重复序列测序的基础上分析了两个Ty1-copia类反转座子的序列特征,并对其异质性进行了研究,为进一步分析反转座子和石刁柏染色体组的稳定性直接关系及了解性染色体进化早期阶段的相关信息提供参考依据。
1 材料与方法
1.1 材料
1.1.1 植物材料 石刁柏(品种UC309和TD818) 种植于河南师范大学生物实验园地。采用无激素、化肥栽培方式管理,通过花的结构鉴定性别。石刁柏同属雌雄同株植物文竹(Asparagus setaceus)购自于河南省新乡市农贸花卉市场。
1.1.2 载体与引物 克隆载体为pMD?誖19-T,购自于宝生物工程(大连)有限公司。所用两个反转座子序列引物根据新乡医学院石刁柏基因组高通量测序结果[17]设计,反转座子s101366上游引物序列为5′-GCAATCGTGTTGGTCTAAGT-3′,下游引物序列为5′-CGCCTCTGTAGTTGATAATG-3′,预期长度为635 bp。s107518上游引物序列为5′-TCATTATC- GCTCTGCTCCT-3′,下游引物序列为5′-CTACACA- CCTTCACATTCTC-3′,预期长度为658 bp。引物由北京华大基因研究中心合成。
1.2 方法
1.2.1 石刁柏基因组DNA提取 石刁柏基因组DNA的提取参照文献[18]方法进行改良,分别提取石刁柏2个品种UC309和TD818的雌雄各3个单株基因组DNA。
1.2.2 石刁柏基因组中Ty1-copia反转座子的扩增及分析 PCR的扩增体系均为:25 μL的反应体系中:H2O2 18.9 μL,10×Buffer 2.5 μL,dNTP Mixture 1.5 μL (各2.5 mmol/L),上、下游引物各 0.5 μL(10 μmol/L),Taq酶 0.1 μL(5 U/μL),1 μL的基因组DNA(约200 ng)。反应程序:94 ℃预变性2 min;94 ℃变性50 s,52 ℃复性 1 min,72 ℃延伸 1 min,共 35 个循环;72 ℃再延伸 10 min;4 ℃保存。取扩增产物在1.5%的琼脂糖凝胶进行电泳,电泳结果用OLYMPUS数码相机在紫外灯下进行观察拍照后参照SanPrep柱式DNA胶回收试剂盒(上海生工生物工程技术服务有限公司)的说明书进行扩增产物回收。感受态细胞的制备、转化、阳性克隆的筛选鉴定参照Sambrook等[19];样品送至生工生物工程(上海)有限公司进行测序。
1.2.3 反转座子序列异质性及聚类分析 利用DNAStar软件进行开放阅读框(Open reading frame,ORF)的查找,分析获得序列的反转座子序列特征;利用DNAMAN软件将Ty1-copia反转录转座子序列进行比对,统计并分析序列碱基置换结果;利用MEGA软件构建系统进化树并进行分析。
2 结果与分析
2.1 反转座子s101366和s107518的克隆与测序分析
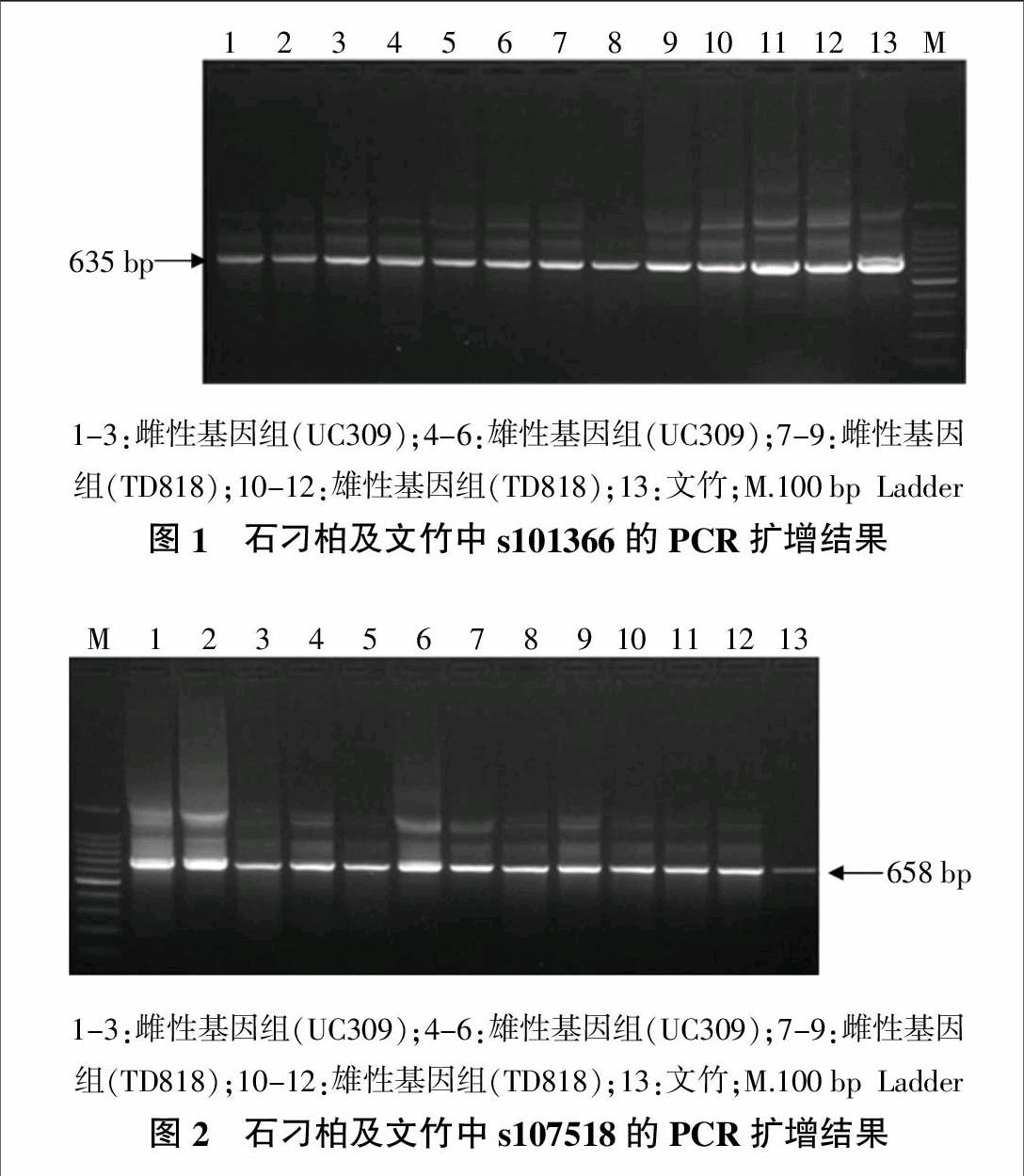
利用已设计的两对特异引物分别对提取的石刁柏(UC309、TD818)的雌、雄各3个单株及文竹中的基因组DNA共13个模板进行PCR扩增,分别扩增出大小约为635和658 bp的产物,其大小均分别与预期条带大小一致(图1,图2)。
将目的条带分别进行回收测序结果表明,对石刁柏及文竹中的s101366进行扩增并克隆测序,分别获得了52条和3条序列,其中雌株序列27条,雄株序列25条,序列长度范围为630~636 bp;而分别对石刁柏及文竹中的s107518进行扩增并克隆测序,各获得了57条和2条序列,其中雌株序列27条,雄株序列30条,序列长度范围为653~661 bp。
通过NCBI中Blast对所获得的石刁柏的109条序列进行搜索,结果表明,s101366的52条序列中有42条序列包含Ty1-copia反转座子逆转录酶保守区,有10条序列包含RNA酶保守区序列;s107518的57条序列中有51条序列包含Ty1-copia反转座子逆转录酶保守区。因此,进一步证明所获得的两个序列属于Ty1-copia反转座子序列。
2.2 反转座子s101366和s107518异质性分析
利用DNAMAN软件将克隆出的石刁柏52条s101366序列进行多序列比对并统计分析碱基变异(图3),结果表明,序列的一致性为92.44%,该序列中有326个突变位点,有67处发生了单碱基置换,32处发生了双碱基置换,43处发生了多碱基置换,其中共发生了2 082次碱基转换、508次碱基颠换、25个T缺失、86个G缺失和46个A缺失,有80%的碱基置换为转换,20%的碱基置换为颠换。碱基转换中T?圮C占51%,G?圮A占49%。
利用DNAMAN软件将克隆出的石刁柏57条s107518序列进行多序列比对并统计分析碱基变异,结果(图4)发现序列的一致性为92.13%,该序列中有227个突变位点,单碱基置换的有80处,双碱基置换的有35处,多碱基置换的有23处,其中共发生了3 039次碱基转换、760次碱基颠换、6个T缺失、2个G缺失、17个A缺失和23个C缺失,有80%的碱基置换为转换,20%的碱基置换为颠换。碱基转换中T?圮C占51%,G?圮A占49%。结果表明,两个石刁柏反转录转座子序列异质性主要是由于碱基转换(T?圮C)造成的。
2.3 反转座子s101366和s107518序列聚类分析
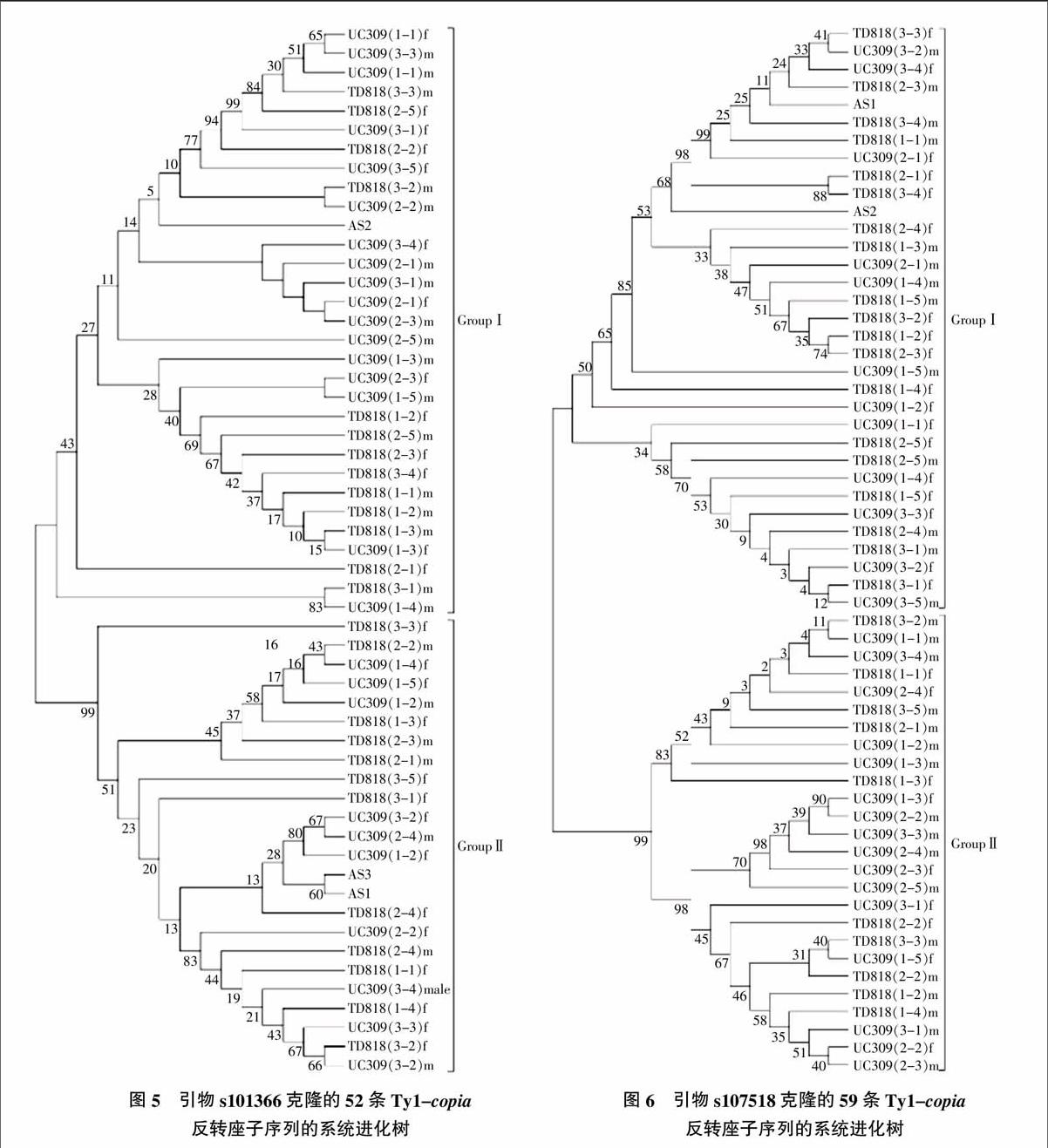
通过DNAMAN软件对s101366和s107518的序列分别进行多序列比对,其序列一致性分别为92.12%和92.08%。利用MEGA软件对s101366和s107518序列构建进化树(图5、图6),从系统进化树中可以看出,两个反转座子序列各聚类为两个大组。据序列的分布情况可知,反转座子s101366和s107518在石刁柏的品种及的雌、雄株性别之间并未表现出明显的规律性。石刁柏两个品种中的s101366和s107518序列分别与其近缘同属的文竹中的s101366和s107518序列也没有显示出明显差异;文竹的一个单克隆As2和另外两个单克隆As1和As3分别分在了GroupⅠ和GroupⅡ,说明即使同一植物中相同类型的反转录转座子序列也存在一定的差异,且有时种内的异质性差异会大于种属之间。
3 小结与讨论
反转座子具有高度异质性的特征,其异质性主要来源于碱基的缺失、突变、插入和置换,且碱基置换是主要的异质性来源。本研究分析的两个反转座子的异质性也主要是来源于碱基置换,其中,碱基转换的频率要明显高于碱基颠换的频率。早期的研究也表明,不论是核内还是核外的基因,基因突变中转换的频率都要远远超出颠换的频率。而且,一般情况下,转换与颠换的概率比为2∶1,这主要是由于密码子的3个位点所受到的选择压力不同,从而造成突变偏向于密码子的第三位点[20,21]。另外,认为甲基化CG序列是基因突变的一个热点,因为甲基化的C可经过脱氨基的作用转换为T,也提高了碱基转换的频率[22,23]。
本研究对序列的聚类分析结果表明,两个Ty1-copia反转座子在品种、性别、单克隆及近缘种之间的序列均未发现明显的特异性,甚至雌雄同株的近缘种文竹的3个克隆序列也未聚为一组,说明所分析的两个反转录转座子的保守性较强,无论是从雌雄同株向雌雄异株的垂直演化还是不同品种间的横向传递过程中,产生了大量的碱基置换、插入和缺失的变异。但是其变异呈现随机性而均未表现出有规律的变异,这也反映了相同植物类型的反转录转座子保守性较强,同一植物中的相同类型的反转录转座子序列变异比较大,甚至种内异质性差异会大于种间[24],这种规律是否在石刁柏整个基因组的反转座子中呈现普遍性还需要进一步分析。
参考文献:
[1] YAMPOLSKY C,YAMPOLSKY H. Distribution of sex forms in the phanerogamic flora[J].Bibliotbeca Genet,1966,3:1-62.
[2] RENNER S S,RICKLEFS R E. Dioecy and its correlates in the flowering plants[J].Am J Bot,1995,82:596-606.
[3] KUMAR S,KUMARI R,SHARMA V. Genetics of dioecy and causal sex chromosomes in plants[J].J Genet,2014,93(1):241-77.
[4] BULL J J.Evolution of Sex Determining Mechanism[M].Menlo Park:Benjamin-Cummings,1983.
[5] 徐 玲,杨 静,刘 林,等.植物反转录转座子的研究进展[J].江西农业学报,2012,24(6):42-46.
[6] NOMA K,OHTSUBO E,OHTSUBO H. Non-LTR retrotransposons(LINEs) as ubiquitous components of plant genomes[J].Molecular and General Genetics,1999,261(1):71-79.
[7] YU J,HU S,WANG J,et al. A draft sequence of the rice genome(Oryza sativa L.ssp. indica)[J].Science,2002,296(5565):79-92.
[8] LING H Q,ZHAO S,LIU D,et al. Draft genome of the wheat A-genome progenitor Triticumurartu[J].Nature,2013,496(7443): 87-90.
[9] FESCHOTTE C,JIANG N,WESSLER S. Plant transposableelements:Where genetics meets genomics[J].Nat Rev Genet,2002, 3(5):329-341.
[10] UMEZU K,HIRAOKA M,MORI M,et al. Structural analysis of aberrant chromosomesthat occur spontaneously in diploid Saccharomyces cerevisiae:Retro-transposon Ty1 plays a crucial role in chromosomal rearrangements[J].Genetics,2002,160:97-110.
[11] MORRISH T A,GARCIA-PEREZ J L,STAMATO T D,et al. Endonuclease-independent LINE-1 retrotranspositionatmammaliantelomeres[J].Nature,2007,446:208-212.
[12] 李书粉,李 莎,邓传良,等.转座子在植物XY性染色体起源与演化过程中的作用[J].遗传,2015,37(2):157-164.
[13] WANG J,NA J K,YU Q,et al. Sequencing papaya X and Yhchromosomes reveals molecular basis of incipient sex chromosome evolution[J].Proc Natl Acad Sci USA,2012,109(34):13710-13715.
[14] LPTIEN H. Identification of the sex chromosome pair in asparagus(Asparagus officinalis L.)[J].Zeitschriftfür Pflanzenzü- chtung,1979,82:162-173.
[15] DENG C L,QIN R Y,WANG N N,et al. Karyotype of asparagus by physical mapping of 45S and 5S rDNA by FISH[J].J Genet,2012,91(2):209.
[16] 高武军,张小莉,尹为治.芦笋性染色体的细胞学研究[J].河南农业科学,2008(10):116-120.
[17] LI S F,GAO W J,ZHAO X P,et al. Analysis of transposable elements in the genome of Asparagus officinalis from high coverage sequence data[J].PLoS One,2014,9(5):e97189.
[18] 邹喻苹,葛 颂,王晓东.系统与进化植物学中的分子标记[M].北京:科学出版社,2001.
[19] SAMBROOK J,FRITSCH EF,MANIATIS T.Molecular Cloning:A Laboratory Manual[J].NY:Cold Spring Harbor Laboratory Press,1989.
[20] 丁 波,张亚平.人与大猩猩,黑猩猩和猩猩亲缘关系的探讨[J].遗传学报,1999,26(6):604-609.
[21] DASSANAYAKE M,OH D H,HAAS J S,et al. The genome of the extremophile crucifer Thellungiella parvula[J].Nature Genetics,2011,43(9):913-918.
[22] GREENBERG B D,NEWBOLD J E,SUGINO A. Intraspecific nucleotide sequence variability surrounding the origin of replication in human mitochondrial DNA[J].Gene,1983,21(1):33-49.
[23] HORAI S,HAYASAKA K. Intraspecific nucleotide sequence differences in the major non-coding region of human mitochondrial DNA[J].Am J Human Genetics,1990,46(4):828-834.
[24] STEINHAUER D A,HOLLAND J J. Direct method for quantitation of extreme polymerase error frequencies at selected single base sites in viral RNA[J].J Virology,1986,57(1):219-228.
猜你喜欢
林业科学(2022年1期)2022-03-23
中国蜂业(2021年5期)2021-05-22
教学考试(高考生物)(2020年6期)2020-11-23
食品与生物技术学报(2020年8期)2020-01-06
科学24小时(2019年5期)2019-06-11
科学之谜(2019年3期)2019-03-28
发明与创新(2019年9期)2019-03-26
科学之谜(2018年8期)2018-09-29
浙江农林大学学报(2016年6期)2016-12-12
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
