
粗柄独尾草不同部位种子特性对不同生境的响应
2016-12-04 03:06石河子大学生命科学学院新疆石河子832000
种子 2016年9期
, , , , (石河子大学生命科学学院, 新疆 石河子 832000)
粗柄独尾草不同部位种子特性对不同生境的响应
王文洁,王丹丹,段呈,张霞,王绍明
(石河子大学生命科学学院, 新疆 石河子 832000)
以新疆准噶尔古尔班通古特沙漠南缘的粗柄独尾草(Eremurusinderiensis)为研究对象,通过划分不同破碎化程度斑块(强、中、弱),并根据植株花序轴高度将其分为长度相等的上部、中部和下部3个部位,研究不同斑块之间植株不同部位粗柄独尾草种子大小以及萌发特性。结果显示: 1) 随着生境破碎化程度的逐渐加强,各部位的种翅宽没有显著变化,而种子千粒重及种子长和宽均呈减小趋势; 2) 种子数量在整个植株、植株上部和中部均随着生境破碎的增强显著降低,而植株下部的种子数量在各斑块之间差异不显著; 3) 生境破碎化弱的植株不同部位种子萌发率均最高,破碎化中的植株上部种子萌发率显著高于破碎化强的,而在植株下部萌发率与之相反。生境破碎化对植物有较大影响,为适应变化生境破碎化和最大程度地繁衍后代,植株不同部位的种子特性会产生相应变化。
生境破碎; 粗柄独尾草; 不同部位; 萌发率
同一物种种群间或种群内,植物个体间或个体内的种子大小差异,是对复杂环境适应的结果。种子大小作为植物生活史中一个重要的性状,影响种子的散布类型、种子数量、种子萌发、幼苗的存活、土壤种子库寿命等[1]。研究表明,植物为适应变化的环境,植株个体的种子大小会产生相应的生物学或生态学特征[2-3]。然而不同环境梯度下,种群间种子大小的差异,对深入了解植物如何响应环境的变化有重要意义。
生境破碎化是指在人为活动或自然干扰下,大块连续分布的自然生境,被其它非适宜生境分隔成许多面积较小、分散且孤立的生境斑块(岛屿)的过程,是目前普遍存在的一种现象[4-6]。生境破碎化可改变种群的扩散和迁入模式、种群遗传和变异等,影响物种的繁殖和迁移能力[7];增大物种灭绝的几率[8];导致生物多样性的下降[9]等。对于不同生境破碎化强度,种群间种子大小和数量会产生相应变化[10],然而同一植株,不同部位的种子有不同的分工[11-12]。种群间植株不同部位的种子大小、数量和萌发率如何变化还少有研究,这对深入剖析植物为适应环境的变化有重要意义。
表1 粗柄独尾草生境破碎化强弱分类

样地编号123456破碎化程度强强中中弱弱样地面积(m2)793.9687.47909.63324.615612.513458.8经纬度44°54′N,85°34′E44°44′N,85°56′E44°53′N,85°36′E44°55′N,85°29′E44°54′N,85°33′E44°46′N,85°52′E周围环境评价南为农田西临道路东北为防护林带四周为农田东为荒地南接道路东北为防护林带西接道路东北为酒庄沙漠斑块沙漠斑块
粗柄独尾草(Eremurusinderiensis)属百合科(Liliaceae)独尾草属(Eremurus)植物,生长在干旱、炎热、多风的沙漠环境中,主要以种子繁殖进行繁衍后代,同时具翅,主要靠风媒传播。在我国仅分布于新疆的北疆地区(青河、阜康、沙湾、裕民),是新疆沙漠地区典型的抗旱抗风沙的早春类短命植物[13-14]。近年来,农田的开垦、道路工程的建设以及防护林工程的建设,造成古尔班通古特沙漠南缘的原有生境破碎化。现粗柄独尾草不同部位种子特性及生态适应性已有研究[11],但没有资料显示不同部位种子在不同的生境中会如何响应破碎化的环境。本研究对不同生境破碎化强度下,粗柄独尾草花序上部、中部和下部的种子大小、数量和萌发率进行比较分析,探讨规律,以期为干旱荒漠地区的生态保护和植被恢复提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于古尔班通古特沙漠南缘(44°44′N~44°56′N,85°28′E~85°56′E)。古尔班通古特沙漠属于温带荒漠气候类型,春季湿润,夏季干燥,冬季寒冷;年平均气温为6~10 ℃,最高气温在40 ℃以上,最热时平均气温为24~27 ℃;年降水量为70~150 mm,年蒸发量为2 000~2 800 mm;冬季的积雪深度一般为10~30 cm。
1.2 生境破碎化评价
根据斑块周围环境、干扰因素和斑块面积大小等[15],选择破碎化程度不同的6个样地,分别为:强、中、弱3个等级,划分结果见表1。
1.3 种子收集及处理
在2013年6月种子成熟期,对研究所需种子进行采集。根据先前调查研究,采集样地设置在不同生境破碎化强度的6个样地,分别为两强、两中和两弱(表1)。不同生境破碎化强度各随机采集20株样本,即每个样地随机选择10株,植株间距不小于5 m。根据每株粗柄独尾草的花序轴高度,将其分为长度相等的上部、中部和下部(近地端)3个部位,分开采收,装入种子袋中,带回实验室进一步处理。
实验测量数据包括种子数量;外形指标包括种翅宽、种子长度和宽度,均由游标卡尺(0.01 mm)测得;种子千粒重,由万分之一天平(0.000 1 g)测得。
1.4 种子萌发率测定
粗柄独尾草种子具有休眠性,因此在萌发实验前4个月,将收集的种子放置于冰箱0 ℃冷冻层保存。1%次氯酸钠消毒3~5 min,经蒸馏水冲洗进行萌发试验。每个不同强度生境破碎化处理设置30个重复。首先将50粒饱满的种子放在装有细沙的培养皿中萌发,培养皿基质为种子生长原生境沙土,每天喷洒一定的蒸馏水,培养皿置于GHP-400智能光照培养箱(温度10 ℃,光照强度4 000 lx,12 h光/12 h暗)中进行培养[16]。胚根突破种皮视为萌发,每24 h统计1次发芽种子数,50 d后结束,计算萌发率。
1.5 统计分析
首先将数据输入Excel表格中。采用SPSS 19.0分析不同生境破碎化强度的种群间,整个植株、植株顶部、中部、基部的种子大小、外形和萌发率的差异性。用非参数检验(Nonparametric Test)中1-Sample K-S 检验数据是否为正态分布,对于正态分布的数据进行方差同质性检验,用单因素方差分析(One-wayANOVA)检验数据的差异性(p=0.05),对不服从正态分布的用K Independent Samples (Kruska-l Wallis H)进行比较。采用Origin 8.5软件做图。

图1 不同破碎化种群粗柄独尾草不同部位种子外形特征
2 结果与分析
2.1 不同破碎化强度下不同部位种子大小与外形比较
由图1可知,不同破碎化程度间,只有不同部位的种翅宽没有显著差异,而种子千粒重、长与宽在不同部位均有不同的显著差异。整个植物、植株中部和植株下部的种子千粒重在破碎化强的生境下显著低于破碎化中与破碎化弱的,且破碎化中和弱生境的种子千粒重没有显著差异。整个植物、植物上部和植株中部的种子宽在破碎化强的生境下显著小于破碎化中与破碎化弱的,且破碎化中和弱生境的种子宽没有显著差异,植株下部在生境破碎化强、中、弱均有显著性差异。整个植物、植物中部和植株下部的种子长在破碎化强、中与弱的生境下均有显著差异,且植株上部的种子长只有在破碎化强和弱的生境下有显著差异。
2.2 不同破碎化强度不同部位种子数差异
由图2可知,粗柄独尾草不同部位的种子数有一定差异。除植株下部种子数在不同生境破碎化梯度下没有显著性差异,整个植株、植株上部和植株中部的种子数在不同程度破碎化间均有显著性差异,整个变化规律为生境破碎化弱的生境种子数显著多于生境破碎化中显著多于生境破碎化强。

图2 不同破碎化强度间不同部位种子数量差异
2.3 不同破碎化强度不同部位萌发率的比较
由表2和图3可知,粗柄独尾草整个植株和植株中部的种子,在生境破碎化弱的生境下萌发率最高;生境破碎化强和中的生境下,种子的萌发率没有显著差异,但生境破碎化强的植株种子萌发率要高一些。生境破碎化中的植株,在植株上部的种子萌发率最高,显著高于破碎化强的植株,与生境破碎化弱的植株没有显著差异。生境破碎化强的植株,在植株下部的种子萌发率最高,达54%,显著高于破碎化中的生境植株,与生境破碎化弱的植株没有显著差异。
表2 粗柄独尾草不同破碎化强度下萌发率的比较(平均值±标准误)

破碎化强破碎化中破碎化弱植株上部44.2±4.498b58.0±3.947ab61.8±6.279a植株中部51.2±6.252b43.2±2.074b66.0±4.486a植株下部54.0±5.303a35.0±2.125b57.2±4.120a植株49.8±3.110b45.4±2.038b61.6±2.904a
注:a和b表示每行的显著性比较,p=0.05。
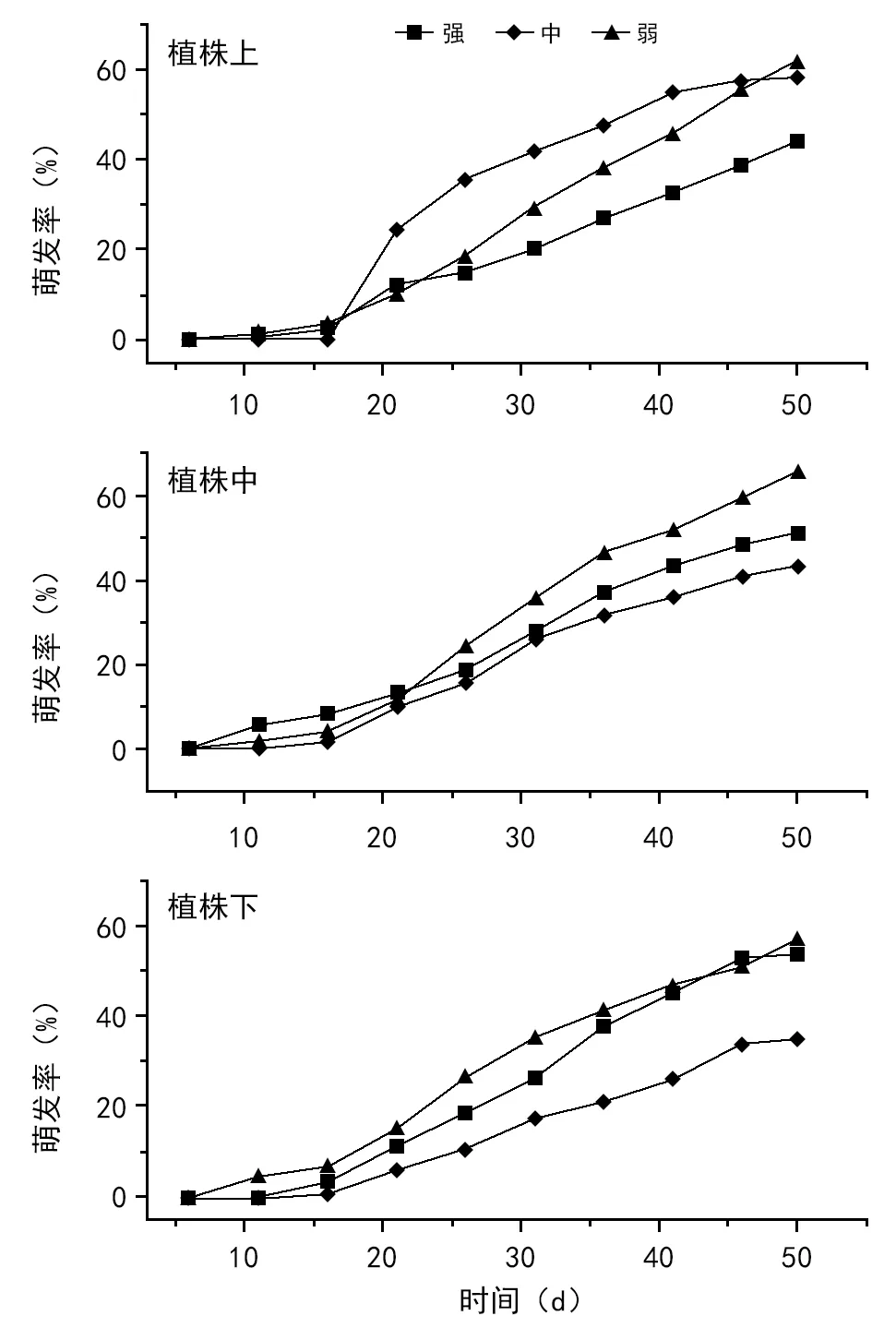
图3 粗柄独尾草不同破碎化强度不同部位种子萌发率比较
3 讨 论
本研究结果表明,生境破碎化对植物不同部位(植株上中下)种子大小和萌发率均有较大影响。种子的特性(大小和萌发率等)是植物生活史的重要特征,然而在生境破碎化选择压力下,植物的种子特性会产生相应变异。随着生境破碎化强度的增加,种子体积和重量会相应变小[10]。本研究结果表明,生境破碎化强、中与弱的种群中植株不同部位种子千粒重相比,生境破碎化中等种群种子的重量没有显著变化,生境破碎化强的重量显著降低,这可能是由于个植物种的种子重量一般被看作具有一个相对稳定的数值和性状[17],影响种子萌发、幼苗建植、存活、个体适合度以及植物生活史特征的表达[18],然后随着生境破碎化的加强,为适应变化的环境,产生了相应变化。生境破碎化强、中与弱种群的植株不同部位种子宽和长相比,总体上是随着生境破碎化增强,植株种子的体积变小,植株产生的种子数量也显著降低,这是植物在不同生境破碎化的适应对策。植株不同部位的种子特性变化不同,如植株下部的种子数量随生境破碎化的增加,没有显著降低,这是因为粗柄独尾草为总状花序,以向顶的顺序形成花和果实,早开放的花比晚开放的花更接近营养,更易夺取资源[19-20],有充分的时间保证种子成熟而不需要对环境做出响应,说明植株不同部位的种子具有一定的分工作用,使种群最大可能的延续。
生境破碎加大了植株间的近亲交配,降低了种群遗传多样性。种子和种翅的大小均与其散布能力有关,轻小和翅大的种子具有较大的散布能力,更容易被生物和非生物等传播媒介所携带和传播,让破碎化程度较强的斑块存活下去[18,21]。本研究结果表明,在破碎化较强的种群中,种子变轻,种翅大小没有显著变化,且随生境破碎的增强,种翅大小有一定的增大,这更有利于小种子在恶劣环境中更多更远地把后代散布出去,在离母体更远的地方建植的可能性较大,避免了近交衰退。
Leishman和Westoby发现,在干旱的条件下,大种子比小种子具有更高的萌发比率[22-23]。然而本研究结果表明,整株植株在破碎化强的生境中种子萌发率低于破碎化中,原因可能是破碎化弱的种群1和种群2周围有防护林和农田,会有灌溉带,所以植物有充足的水分。虽然总体上随着生境破碎化的增强,种子的萌发率降低,但是生境破碎化强的植株下部萌发率显著高于生境破碎化中的种子萌发率,与破碎化弱的无显著差异。这说明顶端花后开放没有充足的时间异花授粉,也可能是随着生境破碎化程度的增加,小斑块导致授粉质量低,种子质量下降,种子的萌发率也随之降低;但生境破碎化增加大种子的灭绝风险,所以粗柄独尾草在生境破碎化强的植株下部的萌发率没有显著降低,这也是对破碎化环境的适应和后代延续的一种对策。
[1]Stanton M L.Seed variation in wild radish:effect of seed size on components of seedling and adult fitness[J].Ecology,1984:1 105-1 112.
[2]武高林,杜国祯.植物种子大小与幼苗生长策略研究进展[J].应用生态学报,2008,19(1):191-197.
[3]柯文山,钟章成,席红安,等.四川大头茶地理种群种子大小变异及对萌发、幼苗特征的影响[J].生态学报,2000,20(7):697-702.
[4]Wilcox D S,Mclellan C H and Dobson A P.Habitat fragmentation in the temperate zone.In:Soul M E ed.Con-servation Biology:The Science of Scarcity and Diversity[M].Sunderland Massachuretts:Sinauer Associates Inc.,1986:257-285.
[5]Lovejoy T E,Bierregaard R O,Rylands A B,et al.Edge effects and other effects of isolation on Amazon forestfragments.In:Soul M E ed.Conservation Biology:The Science of Scarcity and Diversity.Sunderland Mas-sachuretts:Sinauer Associates Inc.,1986:257-285.
[6]Fahrig L.Effects of habitat fragmentation on biodiversity[J].Annual Review of Ecology,Evolution,and Systematics,2003,34:487-515.
[7]Dos Santos K,Kinoshita LS,Santos FAM.Tree species composition and similarity in semi deciduous forest frag-ments of southeastern Brazil[J].Biological Conservatio,2007,135:268-277.
[8]Arroyo-Rodríguez V,Pineda E,Escobar F,et al.Value of small patches in the conservation of plant-species diversi-ty in highly fragmented rainforest[J].Conservation Biology,.2008,23:729-739.
[9]武正军,李义明.生境破碎化对动物种群存活的影响[J].生态学报,2003.23(11):2 424-2 435.
[10]郭丽,吴玲,王绍明,等.破碎化生境中伊犁郁金香种群种子形态特征和萌发特性的适应性演化[J].种子,2015,34(9):1-5,10.
[11]王丹丹,张驰,王绍明,等.粗柄独尾草种子特性及生态适应性研究[J].种子,2015,34(7):16-19.
[12]李孟良.油菜不同部位粒重的差异研究[J].种子,2006,25(6):31-33.
[13]崔乃然,毛祖美,李学禹,等.新疆植物志(6)[M].乌鲁木齐:新疆科技卫生出版,1996:480-482.
[14]王雪芹,蒋进,雷加强,等.古尔班通古特沙漠短命植物分布及其沙面稳定意义[J].地理学报,2003,58(4):598-605.
[15]Fahrig L.Effects of habitat fragmentation on biodiversity[J].Annual Review of Ecology and Systematics,2003,34:487-515.
[16]吴玲,张霞,王绍明.粗柄独尾草种子萌发特性的研究[J].种子,2005,24(7):1-4.
[17]Harper JL,Lovell PH,Moore KG.The shapes and sizes ofseeds[J].Annual ReviewofEcology and Systematics,1970(1):327-356.
[18]于顺利,陈宏伟,种子重量的生态学研究进展[J].植物生态学报,2007,31(6):989-997.
[19]Stephenson AG.Flower and fruit abortion:proximate causes and ultimate functions[J].Annual Review of Ecology and Systematics,1981,12:253-279.
[20]Wyatt R.The reproductive biology of Asclepias tuberose:Ⅰ.Flower number,arrangement,and fruit-set[J].New Phytologist.1980,85(1):119-131.
[21]Venable DL,Brown JS.The selective interactions of disper-sal,dormancy,and seed size as adaptations for reducing risk invariable environments[J].AmericanNaturalist,1988,131:360-384.
[22]Leishman MR,Westoby M.Hypotheses on seed size:tests using the sem-i arid flora of western NewSouth Wales,Australia[J].American Naturalist,1994,143,890-906.
[23]Leishman MR,Westoby M.The role of seed size in seedling establishment in dry soil conditions-experimental evidence from sem-iarid species[J].Journal of Ecology,1994 b,82:249-258.
Different Parts of the Seed Characteristics in Response to the Different Habitats Fragmentation ofEremurusinderiensis
WANGWenjie,WANGDandan,DUANCheng,ZHANGXia,WANGShaoming
(College of Bio-Engineering Shihezi University,Shihezi Xinjiang 832000,China)
habitat fragmentation;Eremurusinderiensis; different position; seed germination rate
2016-04-10
兵团科技攻关项目(编号:2015 AD 023);国家青年科学基金项目(编号:31300406)。
王文洁(1991—),女,硕士研究生,研究方向:植物遗传学。
张 霞,教授,硕士生导师,E-mail:xiazh@shzu.edu.cn。
10.16590/j.cnki.1001-4705.2016.09.015
Q 948.12
A
1001-4705(2016)09-0015-05
Abstrct:This article takeEremurusinderiensisin south of Gurbantunggut Desert as the research object,divided into different degree of fragmentation by patches (strong, medium and weak),and according to the length of inflorescence axis, divide it into equal to the top,middle and bottom(near), to study the characteristics of seeds located on different position in the inflorescence and germination characteristics in different patches. The results showes that:1)As the habitat fragmentation degree gradually strengthen, each part of seed wing width had not change significantly, while seed weight and length and width of seed showed a decreasing trend.2) With enhancement of habitat fragmentation the number of seeds were significantly reduced in whole plants, the top and middle, while the number of seeds in bottom of the plant between the patches were not significant.3) Seed germination rate was the highest in the weaker fragmentation degree, the top of plant seed germination rate in middle fragmentation was significantly higher than stronger fragmentation,and the seed germination rate in the bottom of plant contrast. Habitat fragmentation had a greater impact on the plant, to adapt to changes in habitat fragmentation and the greatest degree of offspring, seed characteristics in different parts of the plant will produce a corresponding change.
猜你喜欢
今日农业(2022年15期)2022-09-20
中国农业科学(2022年13期)2022-07-26
落叶果树(2021年6期)2021-02-12
江苏农业科学(2020年21期)2020-03-08
红土地(2018年7期)2018-09-26
考试周刊(2016年11期)2016-03-17
体育科技(2016年2期)2016-02-28
安徽农业科学(2015年25期)2015-12-22
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
