
过氧化氢和一氧化氮在小球藻抗阿特拉津胁迫中的作用
2016-12-02 05:41陈思范晓季金瑜剑李星星宋昊孙立伟钱海丰
生态毒理学报 2016年4期
陈思,范晓季,金瑜剑,李星星,宋昊,孙立伟,钱海丰,,*
1. 浙江工业大学 海洋学院,杭州 310032 2. 浙江工业大学 生物工程学院,杭州 310032 3. 浙江工业大学 环境学院,杭州 310032
过氧化氢和一氧化氮在小球藻抗阿特拉津胁迫中的作用
陈思1,范晓季2,金瑜剑2,李星星1,宋昊1,孙立伟3,钱海丰1,3,*
1. 浙江工业大学 海洋学院,杭州 310032 2. 浙江工业大学 生物工程学院,杭州 310032 3. 浙江工业大学 环境学院,杭州 310032
过氧化氢(H2O2)和一氧化氮(NO)作为信号分子,可调节植物生长、发育以及应对外源性胁迫。利用过氧化氢酶(CAT)以及NO清除剂(PTIO),研究了除草剂阿特拉津(atrazine,100 μg·L-1)影响小球藻生长的机理,并分析内源性H2O2和NO在小球藻抗除草剂胁迫中的作用。研究结果表明,阿特拉津在诱发小球藻细胞死亡的过程中,不同程度促发了H2O2和NO生成;外源CAT可通过清除H2O2和诱导NO来缓解阿特拉津对小球藻的生长抑制;PTIO与阿特拉津的联合实验进一步证实,小球藻体内的NO诱导与H2O2的爆发无关,它们之间的合成没有相关性。因此,除草剂阿特拉津主要通过诱导小球藻体内的H2O2爆发来破坏藻细胞,抑制其生长,与NO的信号传递无关。
阿特拉津;小球藻;CAT;一氧化氮;过氧化氢;PTIO
Received 14 April 2016 accepted 5 May 2016
过氧化氢(H2O2)是一种常见活性氧物质,主要由线粒体、叶绿体、过氧化物酶体等产生[1-3]。植物的外界胁迫,如病原体、金属化合物和除草剂暴露等,常诱导H2O2的产生,造成DNA损伤甚至细胞死亡[4-8]。不仅如此,大量研究表明,H2O2作为重要的信号分子传递胁迫信号[9-10],并利用水调蛋白将胁迫信号传递至邻近细胞[11-12]。另一个小分子一氧化氮(NO)由一氧化氮合成酶(nitric oxide synthase, NOS)合成。在哺乳动物体内,NO作为重要信号分子参与神经调节、心血管功能和免疫系统调节,但在植物中的功能尚不清楚[13-14]。近年来,NO在植物中的功能也备受关注,被认为是植物生长发育过程中传递应激反应的信号分子[15-16]。研究证实,NO外源性供体硝普钠(sodium nitroprusside, SNP)促使植物具有抗外界胁迫的功能,如盐胁迫[17]、干旱[18]、病原体[19]、金属化合物[20]、温度[21]、涝灾[22]和除草剂胁迫[23],但过量NO也会抑制植物光合作用[23-24]。这些研究表明,NO是植物体内重要的信号分子[25]。
NO和H2O2在植物应激反应中的关系存在很多争议。Lum等[26]发现,H2O2快速诱导绿豆细胞内NO的积累,认为氧化应激反应可诱发NO的大量产生。同样,Lin等[27]证实,H2O2在水稻体内可作为诱导NO的上游信号分子,并且认为NO与H2O2是水稻叶片细胞致死的重要介质。然而,Neill等[21]发现,病原体入侵植物时,植物体内的NO和H2O2同时产生。这些研究说明NO和H2O2在环境胁迫中都起到信号传递的作用,但一些不一致的结论暗示两者之间的复杂关系尚需进一步解析。
阿特拉津作为全球使用最广泛的除草剂之一,在水土中常被大量检出[28]。阿特拉津通过阻碍光合系统II的电子受体,破坏植物光合作用电子传递;并以脂溶性方式进入藻类叶绿体,导致活性氧ROS (reactive oxygen species, ROS)的爆发,对细胞造成氧化性损伤,进而扰乱细胞内氧化还原平衡[29-31]。过多的ROS破坏细胞膜,产生脂质过氧化产物MDA (malondialdehyde, 丙二醛),从而对细胞产生严重损伤[32]。本实验以小球藻为模式生物,研究除草剂胁迫下NO与H2O2随时间的累积情况;清除NO后,H2O2随时间的累积情况;以及清除H2O2后,NO随时间的累积变化,从而阐明NO和H2O2参与植物对抗除草剂胁迫中的相互关系。
1 材料与方法 (Materials and methods)
1.1 实验材料
供试藻类为普通小球藻,购自中国科学院水生物研究所,水生四号培养液,250 mL锥形瓶中培养,培养条件为(25±0.5) °C,光照强度为4 500 lux左右,光暗比为14 h:10 h。接种浓度为1% (细胞浓度约为6.5×105个·mL-1),直至培养到藻液在680 nm下吸光度(OD)值为0.08进行实验。阿特拉津(Atr)(长兴化工公司,浙江,中国)加入培养液中,单独或与1 kU·mL-1过氧化氢酶(CAT;生物技术研究院,海门,中国)或20 μmol·L-1一氧化氮清除剂(2-phenyl-4,4,5,5-tetramethylimidazoline-3-oxide-1-oxyl,2-苯基-4,4,5,5-四甲基咪唑啉-3-氧代-1-氧,PTIO,西格玛奥德里奇,圣路易斯,美国)联合使用最终浓度设置为100 μg·L-1[23];每组设置3个平行组。
1.2 细胞生长以及细胞内NO、H2O2含量的检测
胞内NO含量的测定根据碧云天生物技术公司的试剂盒提供的方法:取10 mL藻液离心得沉淀,取少量石英砂研磨,使用450 mL HEPE (pH 7.2)缓冲液洗涤研磨液,11 000 r·min-1离心5 min,取200 mL上清液,具体根据试剂盒方法测定。胞内过氧化氢含量的测定根据碧云天生物技术公司的试剂盒提供的方法。使用分光光度计测定680 nm下吸收光值,通过标准曲线换算出藻细胞密度。藻密度线性回归方程为:
Y=162.1X+1.3463 (r2=99.34%)
公式中X为OD685值;Y为105细胞数·mL-1。
2 结果(Results)
阿特拉津处理6~48 h后小球藻细胞生长变化见图1A,对照组的小球藻细胞个数在各个时间段都明显多于Atr处理组,说明Atr对小球藻的生长有明显的抑制作用,且随着处理时间延续,抑制率增加,6,12,24,48 h的抑制率分别为11.8%,7.2%,20.4%和29.6%。类似现象在淡水蓝藻、铜绿微囊藻、海洋硅藻以及三角褐指藻中也被证明[33-34]。但不同品种的藻类对阿特拉津敏感度不同,相对灵敏度依次为铜绿微囊藻>小球藻>三角褐指藻。
阿特拉津作为光合作用抑制剂,阻止电子向质体醌的转移,从而阻止吸收的光能转化为电化学能[35]。多余的电子和分子氧结合,产生ROS来破坏细胞结构,抑制藻的生长[36]。为确认ROS在阿特拉津胁迫下的作用,我们进一步检测了H2O2和NO的表达水平。结果表明,阿特拉津明显刺激了H2O2和NO的产生(图1B和C)。其中H2O2的表达量在6 h达到峰值(3.78倍),且在48 h内持续保持高水平(图1B);而NO的表达水平直到12 h处理后,才达到峰值(1.3倍),且在24 h内回落至正常水平(图1C)。在阿特拉津抑制小球藻生长过程中,均发现H2O2和NO含量在0~12 h处理期间出现峰值,推测这2种物质都参与除草剂诱导小球藻细胞死亡过程。另外,通过观察NO和H2O2到达峰值的时间,结合相关报道推测,NO应该位于H2O2的下游[26,37-38]。
过氧化氢酶(CAT)可将H2O2分解成氧和水,为了确认NO的产生是否依赖于H2O2,我们检测了阿特拉津与CAT联合暴露下小球藻的生长状况。结果显示,1 kU·mL-1的CAT可以完全降解细胞内的H2O2。如图2A所示,CAT不仅能缓解阿特拉津对小球藻的抑制作用,甚至略有促进。在阿特拉津和CAT共同作用6,12,24,36和48 h后,细胞数量为对照组的1.4,1.86,1.73,1.82和1.59倍。有意思的是,CAT处理下,NO仍显著增加,在12,24,36和48 h处理后NO表达量分别为对照组的1.75,2.07,3.22和1.6倍(图2B)。
原研究表明,由于NO的抗氧化特性,能够簇灭细胞内ROS,对细胞起到保护作用[39],使植物受到环境胁迫(如金属化合物、盐、低温胁迫、病毒侵入)时产生耐受性[40-41,17]。本实验证实,相比于阿特拉津单独处理,外源CAT在完全降解H2O2的同时,同时促进NO的大量合成。该结果表明,H2O2的生成是阿特拉津抑制小球藻生长的关键因子,而过多的H2O2可抑制NO的产生。尽管NO也是生物活性分子,但它在植物体内的正效应与其浓度相关,而过高浓度的NO对植物起到抑制效应[22,42]。这也解释了为什么适当的外源性NO可缓解除草剂的胁迫压力[23],而高浓度的外源性NO也会抑制藻细胞的生长。

图1 阿特拉津对小球藻生长(A),H2O2含量(B)以及NO含量(C)的影响Fig. 1 Effect of atrazine on algal growth (A), H2O2(B) and NO content (C)
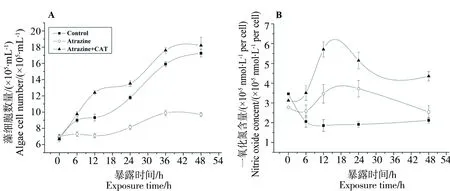
图2 阿特拉津和CAT共同暴露对小球藻生长(A),H2O2含量(B)以及NO含量(C)的影响Fig. 2 Effect of atrazine and CAT on algal growth (A), H2O2(B) and NO content (C)
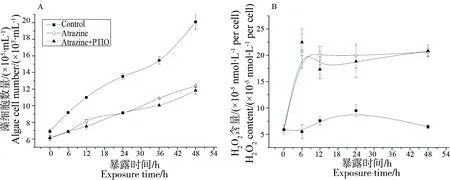
图3 阿特拉津和一氧化氮清除剂(PTIO)共同暴露对小球藻生长(A),H2O2含量(B)以及NO含量(C)的影响Fig. 3 Effect of atrazine and the NO scavenger (PTIO) on algal growth (A), H2O2(B) and NO (C) content
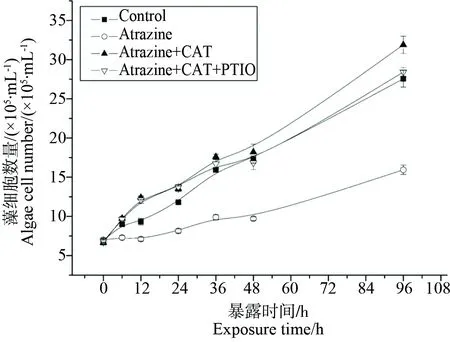
图4 阿特拉津、CAT和PTIO共同暴露对小球藻生长(A),H2O2含量(B)以及NO含量(C)的影响Fig. 4 Effect of combined atrazine, CAT and PTIO treatment on (A) algal growth, the content of (B) H2O2 and (C) NO
利用PTIO能清除NO的特性,我们选取PTIO与阿特拉津共同处理,进一步确定NO在除草剂胁迫中的作用。如图3A所示,PTIO虽能清除NO,但并不能缓解小球藻生长受抑制的现象。当清除NO之后,PTIO组与阿特拉津处理组小球藻生长都受到抑制,说明NO并不直接参与阿特拉津对小球藻生物的抑制效应。我们同时检测0~48 h中藻细胞内H2O2含量变化,发现在清除NO后,H2O2的含量明显高于对照组。在小球藻生长受抑制的情况下,H2O2的含量显著增加,说明H2O2是直接参与阿特拉津对小球藻的致死效应。而在清除了NO之后,藻内的H2O2量并没有明显降低,说明NO并不指导H2O2的合成,且NO位于H2O2信号传导的下游途径。
对于阿特拉津抑制小球藻生长的2种机理推测,将H2O2清除剂(CAT)和NO特异性清除剂(PTIO)同时加入阿特拉津处理的小球藻中,我们发现加入CAT、PTIO和共同暴露组的细胞个数明显多于阿特拉津单独处理组, 说明在清除H2O2和NO的情况下,对小球藻生长具显著缓解作用。因此,我们认为H2O2的有无,对阿特拉津的毒性有至关重要的作用,进一步证明阿特拉津的毒性是通过H2O2的增加来实现的。而NO并没有参于此胁迫过程,内源的NO也不能缓解除草剂的胁迫。在其他外界胁迫下,H2O2和NO的关系还需要更多实验的验证。
[1] Jabs T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals [J]. Biochemical Pharmacology, 1999, 57(3): 231-245
[2] Niyogi K K. Photoprotection revisited: Genetic and molecular approaches [J]. Annual Review of Plant Biology, 1999, 50(1): 333-359
[3] Xu H, Zhang J, Zeng J, et al. Inducible antisense suppression of glycolate oxidase reveals its strong regulation over photosynthesis in rice [J]. Journal of Experimental Botany, 2009, 60(6): 1799-1809
[4] Apel K, Hirt H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction [J]. Annual Review of Plant Biology, 2004, 55: 373-399
[5] Qian H F, Lu T, Peng X F, et al. Enantioselective phytotoxicity of the herbicide imazethapyr on the response of the antioxidant system and starch metabolism in Arabidopsis thaliana [J]. PLoS One, 2011, 6(5): e19451
[6] Qian H, Peng X, Han X, et al. Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana [J]. Journal of Environmental Sciences, 2013, 25(9): 1947-1956
[7] Hieno A, Naznin H A, Sawaki K, et al. Analysis of environmental stress in plants with the aid of marker genes for H2O2responses [J]. Methods in Enzymology, 2013, 527: 221-237
[8] Rejeb K B, Benzarti M, Debez A, et al. NADPH oxidase-dependent H2O2production is required for salt-induced antioxidant defense in Arabidopsis thaliana [J]. Journal of plant Physiology, 2015, 174: 5-15
[9] Dat J, Vandenabeele S, Vranová E, et al. Dual action of the active oxygen species during plant stress responses [J]. Cellular and Molecular Life Sciences, 2000, 57(5): 779-795
[10] Schippers J H M, Nguyen H M, Lu D, et al. ROS homeostasis during development: An evolutionary conserved strategy [J]. Cellular and Molecular Life Sciences, 2012, 69(19): 3245-3257
[11] Allan A C, Fluhr R. Two distinct sources of elicited reactive oxygen species in tobacco epidermal cells [J]. The Plant Cell, 1997, 9(9): 1559-1572
[12] Schmidt S R, Caldana C, Mueller-Roeber B, et al. The contribution of SERF1 to root-to-shoot signaling during salinity stress in rice [J]. Plant Signaling and Behavior, 2014, 9(1): e27540
[13] Mayer B, Hemmens B. Biosynthesis and action of nitric oxide in mammalian cells [J]. Trends in Biochemical Sciences, 1997, 22(12): 477-481
[14] Ignarro L J. Nitric Oxide: Biology and Pathobiology [M]. San Diego, CA, 2000: 23-40
[15] Besson-Bard A, Courtois C, Gauthier A, et al. Nitric oxide in plants:Production and cross-talk with Ca2+signaling [J]. Molecular Plant, 2008, 1(2): 218-228
[16] Fröhlich A, Durner J. The hunt for plant nitric oxide synthase (NOS):Is one really needed? [J]. Plant Science, 2011, 181(4): 401-404
[17] Uchida A, Jagendorf A T, Hibino T, et al. Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice [J]. Plant Science, 2002, 163(3): 515-523
[18] García-Mata C, Lamattina L. Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress [J]. Plant Physiology, 2001, 126(3): 1196-1204
[19] Alvarez M E, Pennell R I, Meijer P J, et al. Reactive oxygen intermediates mediate a systemic signal network in the establishment of plant immunity [J]. Cell, 1998, 92(6): 773-784
[20] Singh H P, Kaur S, Batish D R, et al. Nitric oxide alleviates arsenic toxicity by reducing oxidative damage in the roots of Oryza sativa (rice) [J]. Nitric Oxide, 2009, 20(4): 289-297
[21] Neill S J, Desikan R, Clarke A, et al. Hydrogen peroxide and nitric oxide as signalling molecules in plants [J]. Journal of Experimental Botany, 2002, 53(372): 1237-1247
[23] Qian H, Chen W, Li J, et al. The effect of exogenous nitric oxide on alleviating herbicide damage in Chlorella vulgaris [J]. Aquatic Toxicology, 2009, 92(4): 250-257
[24] Wodala B, Ördög A, Horváth F. The cost and risk of using sodium nitroprusside as a NO donor in chlorophyll fluorescence experiments [J]. Journal of Plant Physiology, 2010, 167(13): 1109-1111
[25] Beligni M V, Lamattina L. Nitric oxide interferes with plant photo-oxidative stress by detoxifying reactive oxygen species [J]. Plant, Cell and Environment, 2002, 25(6): 737-748
[26] Lum H K, Butt Y K C, Lo S C L. Hydrogen peroxide induces a rapid production of nitric oxide in mung bean (Phaseolus aureus) [J]. Nitric Oxide, 2002, 6(2): 205-213
[27] Lin A, Wang Y, Tang J, et al. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice [J]. Plant Physiology, 2012, 158(1): 451-464
[28] Flynn K, Spellman T. Environmental levels of atrazine decrease spatial aggregation in the freshwater mussel, Elliptio complanata [J]. Ecotoxicology and Environmental Safety, 2009, 72(4): 1228-1233
[29] Qian H, Sheng G D, Liu W, et al. Inhibitory effects of atrazine on Chlorella vulgaris as assessed by real-time polymerase chain reaction [J]. Environmental Toxicology and Chemistry, 2008, 27(1): 182-187
[30] Qian H, Pan X, Chen J, et al. Analyses of gene expression and physiological changes in Microcystis aeruginosa reveal the phytotoxicities of three environmental pollutants [J]. Ecotoxicology, 2012, 21(3): 847-859
[31] Qian H, Tsuji T, Endo T, et al. PGR5 and NDH pathways in photosynthetic cyclic electron transfer respond differently to sublethal treatment with photosystem-interfering herbicides [J]. Journal of Agricultural and Food Chemistry, 2014, 62(18): 4083-4089
[32] Bai X, Sun C, Xie J, et al. Effects of atrazine on photosynthesis and defense response and the underlying mechanisms in Phaeodactylum tricornutum [J]. Environmental Science and Pollution Research, 2015, 22(22): 17499-17507
[33] Qian H F, Wei Y, Bao G J, et al. Atrazine affects the circadian rhythm of Microcystis aeruginosa [J]. Chronobiology International, 2014, 31(1): 17-26
[34] RebeccaJ W, Simon M M, Ben J K. Determining the relative sensitivity of benthic diatoms to atrazine using rapid toxicity testing: A novel method [J]. Science of the Total Environment, 2014, 485: 421-427
[35] Rutherford A W, Krieger-Liszkay A. Herbicide-induced oxidative stress in photosystem II [J]. Trends in Biochemical Sciences, 2001, 26(11): 648-653
[36] Miller G, Shulaev V, Mittler R. Reactive oxygen signaling and abiotic stress [J]. Physiologia Plantarum, 2008, 133(3): 481-489
[37] She X P, Song X G, He J M. Role and relationship of nitric oxide and hydrogen peroxide in light/dark-regulated stomatal movement inVicia faba [J]. Acta Botabica Sinica-English Edition, 2004, 46(11): 1292-1300
[38] Bright J, Desikan R, Hancock J T, et al. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2synthesis [J]. The Plant Journal, 2006, 45(1): 113-122
[39] Wang Y S, Yang Z M. Nitric oxide reduces aluminum toxicity by preventing oxidative stress in the roots of Cassia tora L[J]. Plant and Cell Physiology, 2005, 46(12): 1915-1923
[40] Prochazkova D, Sumaira J, Wilhelmova N, et al. Reactive nitrogen species and the role of NO in abiotic stress emerging technologies and management of crop stress tolerance [J]. A Sustainable Approach, 2014, 2: 249-266
[41] Delldonne M, Xia Y, Dixon R A, et al. Nitric oxide functions as a signal in plant disease resistance [J]. Nature, 1998, 394(6693): 585-588
[42] Ahmckeness S, Surplus S L, Blake P, et al. Ultraviolet-B-induced stress and changes in gene expression in Arabidopsis thaliana: Role of signalling pathways controlled by jasmonic acid, ethylene and reactive oxygen species [J]. Plant, Cell and Environment, 1999, 22(11): 1413-1423
Effect of H2O2and NO on Atrazine Stress Resistance in Chlorella vulgaris
Chen Si1, Fan Xiaoji2, Jin Yujian2, Li Xingxing1, Song Hao1, Sun Liwei3, Qian Haifeng1,3,*
1. Ocean College, Zhejiang University of Technology, Hangzhou 310032, China 2. College of Biotechnology and Bioengineering, Zhejiang University of Technology, Hangzhou 310032, China 3. College of Environment, Zhejiang University of Technology, Hangzhou 310032, China
Hydrogen peroxide (H2O2) and nitric oxide (NO) have been suggested to function as signaling molecules in plants to regulate growth, development and stress responses. In this study, we investigated the roles of endogenous H2O2and NO in herbicide stress of the algae, Chlorella vulgaris. We treated algae with the herbicide atrazine (100 μg·L-1) alone, in combination with the H2O2-degrading enzyme catalase (CAT; 1 kU·mL-1), or with the NO scavenger 2-phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl3-oxide (PTIO; 20 μmol·L-1) for 48 h and analyzed algal growth, H2O2and NO contents. Atrazine treatment strongly induced H2O2levels by 1.5 to 4.6 folds, mildly stimulated NO levels, and significantly increased the rate of algal cell death. CAT supplementation degraded all detectable H2O2, further increased NO levels and completely reversed the inhibitory effect of atrazine on algal growth while PTIO (the NO scavenger) had no effect on atrazine toxicity in Chlorella vulgaris. Therefore, the herbicidal effect of atrazine in Chlorella vulgaris is mediated mainly by overproduction of H2O2, and endogenous NO had no protective properties against this toxicity.
atrazine; Chlorella vulgaris; CAT; NO; H2O2; PTIO
国家自然科学基金(21577128);浙江省大学生科技创新活动计划(新苗人才计划)
陈思(1992-),女,硕士,研究方向为环境毒理学,Email: cslock@126.com
*通讯作者(Corresponding author), E-mail: hfqian@zjut.edu.cn
10.7524/AJE.1673-5897.20160414002
2016-04-14 录用日期:2016-05-05
1673-5897(2016)4-102-06
X171.5
A
简介:钱海丰(1973—),男,博士,教授,研究方向为环境毒理学。
陈思, 范晓季, 金瑜剑, 等. 过氧化氢和一氧化氮在小球藻抗阿特拉津胁迫中的作用[J]. 生态毒理学报,2016, 11(4): 102-107
Chen S, Fan X J, Jin Y J, et al. Effect of H2O2and NO on atrazine stress resistance in Chlorella vulgaris [J]. Asian Journal of Ecotoxicology, 2016, 11(4): 102-107 (in Chinese)
猜你喜欢
中国机械工程(2023年3期)2023-02-20
西南农业学报(2022年8期)2022-10-28
现代农业科技(2022年4期)2022-03-08
中国机械工程(2022年3期)2022-02-24
环境保护与循环经济(2021年7期)2021-11-02
中国机械工程(2021年11期)2021-06-23
中国机械工程(2019年17期)2019-09-19
甘肃农业大学学报(2019年4期)2019-09-18
陶瓷学报(2019年5期)2019-01-12
贵州农业科学(2016年10期)2016-04-11
