
甜菜育种中遗传漂变现象的分析与探讨
2016-12-02 01:30:12胡华兵贺碧微闫甜甜
中国糖料 2016年4期
胡华兵,贺碧微,闫甜甜
(新疆石河子农业科学研究院甜菜研究所,石河子832011)
在一个小群体内,从基因库抽样形成下一代个体的配子时,就会产生较大的误差,这种误差会引起群体内基因频率、基因型频率的偶然变化,叫作随机遗传漂变(random genetic drift),简称遗传漂变(genetic drift)[1]。
在甜菜不育系和保持系提纯、利用自交系创新选育不育系和保持系育种实践中,由于需要不断自交、回交,分离选育出多对不育系和保持系,这些保持系作为姊妹系,虽然存在血缘关系,但由于发生遗传漂变,各材料之间产质量、抗病性、叶片、叶丛等经济学性状、生物学性状发生严重分化,生成各种类型新群体。同样,在甜菜良种繁育中,抽样群体过小或其他栽培措施、隔离条件等原因亦能导致遗传漂变,致群体基因频率、基因型频率发生改变,种性也发生改变。
本文将从理论、育种实践二方面分析甜菜发生遗传漂变的影响,提出相应对策,探讨甜菜育种、良种繁育实践操作方法。
1 遗传漂变理论分析
从理论上讲,某一群体内各个个体经过随机交配,其后代组成的群体基因频率、基因型频率会达到一种平衡。但是由于某些原因使这种平衡会遭到破坏,比如群体内混杂其他个体、人为干预选择、环境胁迫、取样时样本量过小、基因突变等。样本量过小,样本所含有的基因频率、基因型频率与原有群体有误差,小群体内基因分离与重组不能达到完全的自由分离与重组,致使后代基因频率、基因型频率发生改变,即发生遗传漂变。样本量越小,遗传漂变发生越严重。
基因自然变异概率极低,短时间内对后代群体基因影响有限。小群体自交导致基因频率、基因型频率改变的主要原因是遗传漂变。遗传漂变会导致后代群体某些性状发生改变。徐孟良等人[2]研究表明,不同世代温敏核不育水稻96-5-2S育性对低温的反应不同,该材料在加代繁殖过程中,临界温度有缓慢升高的趋势,第1~4代临界温度保持相对稳定,第5~6代时临界温度显著升高。之所以发生这种现象,可能是有较多等位基因控制临界温度的缘故,并且一些等位基因频率较小,在加代繁殖中所抽样本量有限,各基因自由分离与重组不够充分,后代发生遗传漂变。所以,抽取一定样本繁殖代数越多,遗传漂变作用发生越明显,致使后代性状发生显著变化。
盖钧镒[3]研究自花授粉作物大豆结果表明,随着群体繁殖次数增加,标记基因型变异和稀少标记基因型丢失风险增加,特别是在标记基因型频率小于0.05时;研究异花授粉作物玉米结果表明,随着群体繁殖次数增加,标记基因型变异和稀少标记基因型丢失风险增加。同时,为了保持稀少基因频率、基因型频率,控制试验规模和费用,通过分析计算,给出了这二种作物的繁殖样本容量。
2 育种实践
2.1 不育系和保持系提纯
甜菜育种实践中,不育系和保持系出现纯度下降,即不育率降低,粒性紊乱,需要重新提纯。常用的方法是株对株提纯。在株对株提纯过程中,保持系单株需要不断自交,然后与不育株回交,这种由保持系单株自交、回交而来组成的群体极容易发生遗传漂变现象。即使经过多代多次的自交,个体基因纯合度不断提高,但仍然有一定杂合基因,也就是说基因纯合是相对的,杂合才是绝对的。杂合的微量基因有控制叶片、叶形、抗病性、抗逆性、块根产量、含糖率等生物学性状和经济学性状的基因,这些基因经过不充分分离与重组,某些基因纯合或丢失,导致一些性状得到强化或削弱。例如,原不育系和保持系叶面小波浪皱褶,但新不育系和保持系群体出现大波浪皱褶,叶面严重扭曲变形,这种叶面性状得到进一步加强。新不育系和保持系作为亲本材料,不仅其经济学性状、生物学性状发生改变,而且配合力尤其是一般配合力发生变化。某一不育系、保持系群体达到遗传平衡,其基因频率、基因型频率保持相对稳定,产质量、配合力等各性状也处于一种稳定状态,因遗传漂变导致基因频率、基因型频率改变,产质量、配合力等性状亦将处于另外一种稳定状态。这也是育种实践中经过提纯后的新不育系配合力发生变化的原因。因此不能将新的不育系、保持系群体等同于原不育系、保持系群体。
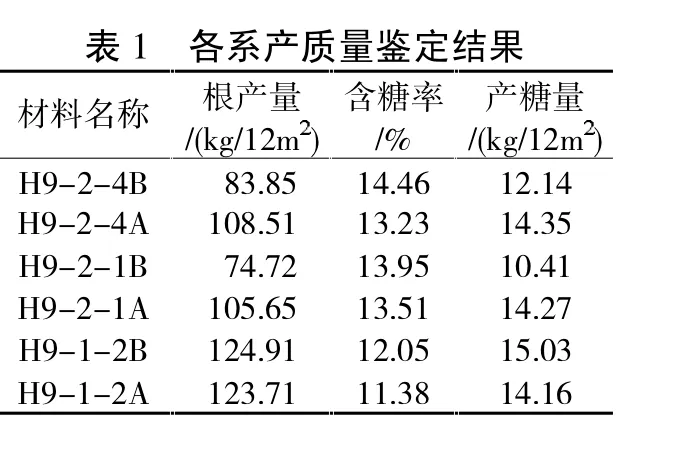
在株对株提纯过程中,会出现多对不育系、保持系的姊妹系,由于遗传漂变姊妹系之间各性状出现明显不一样。例如以H9A、H9B为单株材料进行株对株提纯,多年多代之后产生了3对不育系、保持系姊妹系,经2014年大田鉴定其产质量发生明显分化。各姊妹系田间产质量鉴定结果见表1。
利用EXCEL进行块根产量、含糖率、产糖量多重比较分析。表2反映了各系小区块根产量多重比较关系,可以看出,如果单纯以各保持系来比较,H9-2-1B块根产量与H9-2-4B差异不显著,但和H9-1-2B差异极显著;H9-1-2B块根产量与H9-2-4B差异极显著。表明各保持系虽有血缘关系,但由于多代自交,块根产量分化明显。
材料名表称 根产量1各系产质量鉴定结果H9-2-4B 83.85 14.46 12.14 A 101 323 435 H9-2-1B 74.72 13.95 10.41 A 10565 51 427 H9-1-2B 24.91 12.05 15.03 H9-1-2A 123.71 11.38 14.16 /(kg/12m) 含糖率/% 产糖量/(kg/12m)

H9-1-2B H9-1-2A H9-2-4A H9-2-1A H9-2-4B H9-2-1B表2各系小区根产量(kg )多重比较—LSD法材料及产量 124.91 123.71 108.51 105.65 83.85 74.72 H9-2-1B 74.72 1.16217E-05** 1.47446E-05** 0.0004226** 0.0008554** 0.2176484 H9-2-4B 83.85 7.83583E-05** 0.000102437** 0.0042713** 0.0090959** H9-2-1A 105.65 0.01777804* 0.024380136* 0.6902131 4 81 363186 5122 H9-1-2A 123.71 0.867101904 B 49注:*表示0.05水平上显著性,**表示0.01水平上显著性

H9-2-4B H9-2-1B H9-2-1A H9-2-4A H9-1-2B H9-1-2A各系块根含糖率(%表3)多重比较—LSD法材料及产量 14.46 13.95 13.51 13.23 12.05 11.38 H9-1-2A 11.38 0.0004408** 0.0017712** 0.0061935** 0.0139022* 0.3202548 H9-1-2B 12.05 0.0027541** 0.011917* 0.0419917* 0.0905623 H9-2-4A 13.23 0.0799101 0.2847189 0.6709504 H9-2-1A 13.51 0.1653031 0.5067796 B 9 4474 H9-2-4B 14.46注:*表示0.05水平上显著性,**表示0.01水平上显著性
材料及产量 15.03 14.35 14.27 14.16 12.14 10.41 H9-2-1B 10.41 0.0035961** 0.0096456** 0.0107627* 0.0126634* 0.2001122 H9-1-2B H9-2-4A H9-2-1A H9-1-2A H9-2-4B H9-2-1B表4各系小区产糖量(kg )多重比较—LSD法H9-2-4B 12.14 0.0438669* 0.1114469 0.1229769 0.1420217 H9-1-2A 14.16 0.5093903 0.8859665 0.9317019 21 27 645052 9342 H9-2-4A 14.35 0.6034001 12B 50注:*表示0.05水平上显著性,**表示0.01水平上显著性
表3表示了各系块根含糖率多重比较关系,可以看出,H9-2-4B含糖率与H9-2-1B差异不显著,但与H9-1-2B差异极显著;H9-2-1B含糖率与H9-1-2B呈显著性关系。表明各保持系由于自交多次,含糖率之间差异明显。
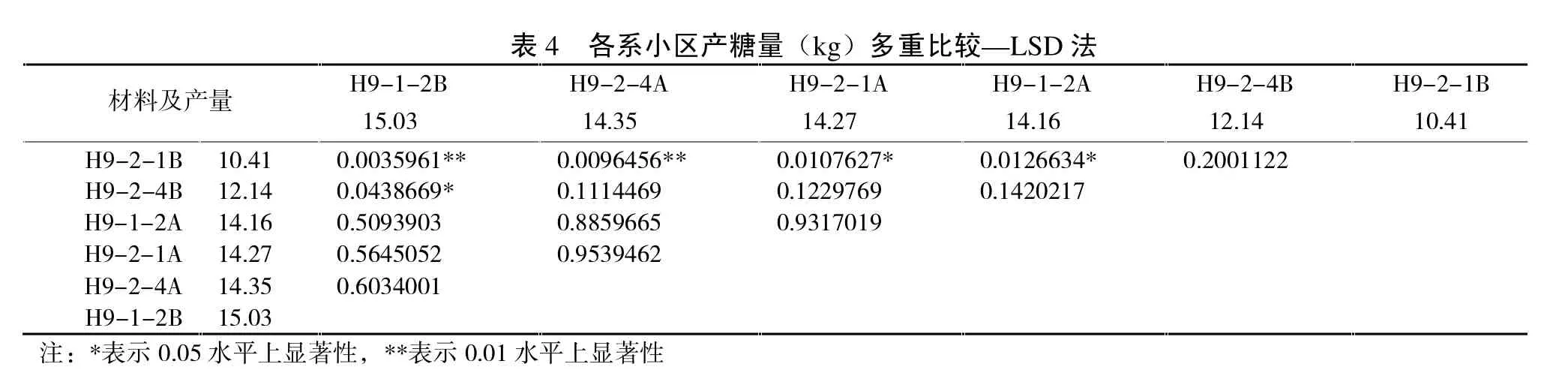
表4表示了各系小区产糖量多重比较关系,可以看出H9-2-4B产糖量与H9-2-1B差异不显著,与H9-1-2B差异显著;H9-2-1B产糖量与H9-1-2B差异极显著。表明各保持系由于多代自交,遗传漂变导致产糖量之间发生显著区别。
2.2 利用自交系创新选育不育系和保持系
利用自交系创新选育不育系和保持系是甜菜育种中常用的方法,在自主选育不育系和保持系中发挥了重要作用。以自交系N3019为原始群体,株对株创新选育出多对不育系和保持系,各保持系存在姊妹关系,经2014年大田鉴定各材料相互之间表型性状差异较大,有些性状得到强化或削弱,与原始群体比较也有较大差异(详见表5)。不仅材料抗病性发生了分化,叶片、叶丛、叶色等田间生物学性状也发生了明显的分化。
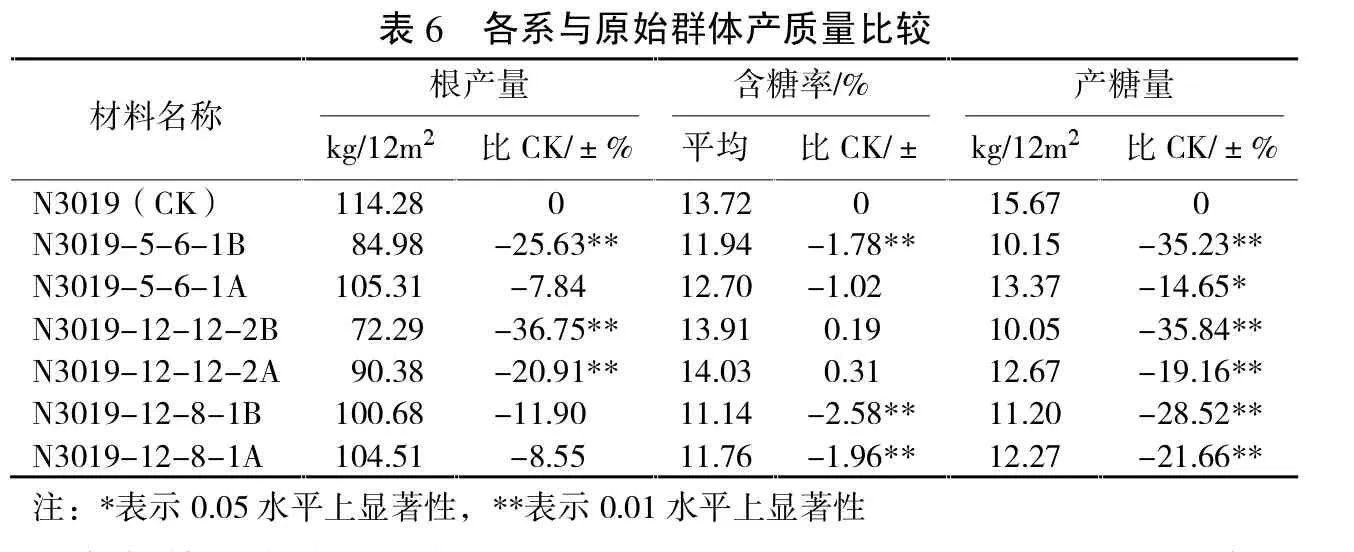
表6反映了创新后不育系、保持系与原自交系产质量比较情况。可以看出,经过多年多代强制自交,块根产量、产糖量减产幅度大,如果单纯以各保持系来分析比较,N3019-5-6-1B、N3019-12-12-2B块根产量较对照减产差异达到极显著水平,N3019-12-8-1B减产不显著;N3019-5-6-1B、N3019-12-8-1B含糖率较对照降低达到极显著水平,但N3019-12-12-2B较对照反而提高0.19度,但差异不显著;3份新保持系产糖量均较对照减产,都呈极显著关系。
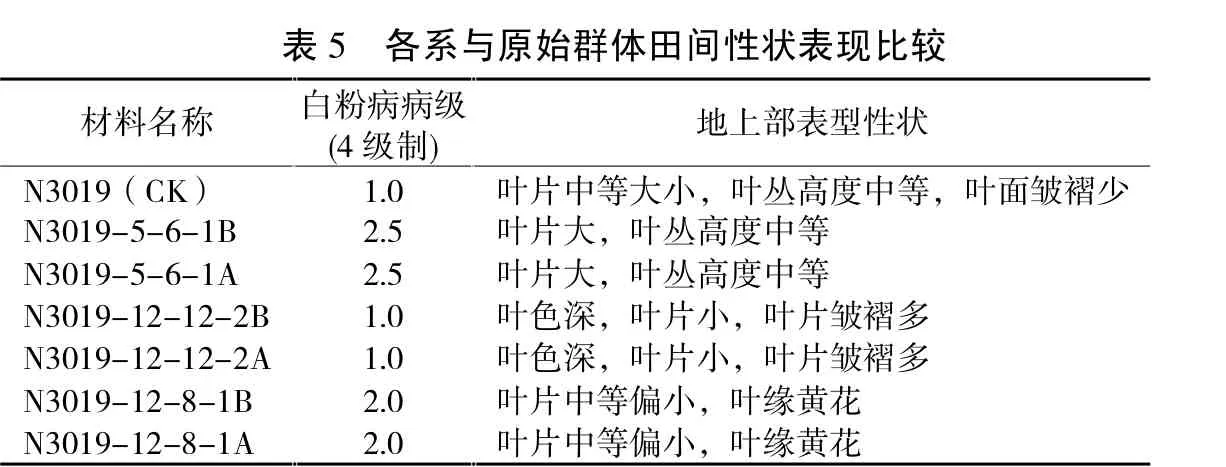
表5各系与原始群体田间性状表现比较N3019(ck) 1.0 叶片中等大小,叶丛高度中等,叶面皱褶少材料名称 白粉病级(4级制) 地上部表型性状N3019-5-6-1B 2.5 叶片大,叶丛高度中等N3019-5-6-1A 2.5 叶片大,叶丛高度中等12-12-2B 10 色深片小,叶片皱褶多N3019-A . 叶,叶N3019-12-8-1B 2.0 叶片中等偏小,叶缘黄花A
材料名称 kg/12m 比cK/±% 平均 比cK/± kg/12m 比cK/±% N3019(cK) 114.28 0 13.72 0 15.67 0表6根产量各系与原始群体产质量比较含糖率/%产糖量N3019-5-6-1B 84.98 -25.63** 11.94 -1.78** 10.15 -35.23** N3019-5-6-1A 105.31 -7.84** 12.70 -1.02** 13.37 -14.65** N3019-12-12-2B 72.29 -36.75** 13.91 0.19** 10.05 -35.84** N3019-12-12-2A 90.38 -20.91** 14.03 0.31** 12.67 -19.16** N3019-12-8-1B 100.68 -11.90** 11.14 -2.58** 11.20 -28.52** N3019-12-8-1A 104.51 -8.55** 11.76 -1.96** 12.27 -21.66**注:*表示0.05水平上显著性,**表示0.01水平上显著性
3 结论与探讨
小群体样本授粉繁殖中,由于基因自由分离重组不充分,导致后代群体基因频率和基因型频率发生改变。如果以单株材料进行繁殖,将会得到基因更加纯合的个体,同时部分基因丢失,遗传漂变发生更加严重,后代群体各性状发生明显的分化,一些性状表现或加强或削弱。
以单株材料提纯不育系和保持系,其后代各姊妹系之间块根产量、含糖率、产糖量均发生明显的分化,相互之间甚至呈极显著关系。
以自交系创新选育不育系和保持系,其后代成系材料与原始群体产质量、抗病性、叶丛、叶片等经济学性状和生物学性状亦有明显区别,块根产量、产糖量均减产,含糖率出现高低分化,与原始群体多呈极显著关系。
理论上,像异花授粉作物甜菜,如果群体数量足够大,在没有突变、选择、混杂和胁迫的前提下,通过完全随机交配,其原有的基因频率、基因型频率可以保持不变[4]。但在实际育种实践中,种子更新繁育群体不可能足够大;并且在实际种子繁育中,风向、隔离措施、田间种植环境等都会严重影响花粉随机传粉,这样就会发生遗传漂变,导致后代的基因频率、基因型频率发生改变。遗传漂变是种性改变的重要原因之一。
可以采取以下措施控制、减缓遗传漂变的发生。
(1)加大繁育群体数量,数量越大越好。对于甜菜最低繁育样本容量,目前还没有相关深入研究。样本群体数量越大,可繁育代数相对越多。(2)材料一旦成系,尽量大群体繁殖足够多的种子,减少繁育代数。(3)在群体繁殖时,做好田间管理措施,减少因干旱、湿涝、盐碱、杂草、病虫害等带来的胁迫,避免非人为定向选择压产生遗传漂变。(4)甜菜作为异花授粉作物,主要传粉媒介是风,因此在繁育中应该重视隔离条件、种植方向与规模和风向的关系,尽量使群体自由传粉。(5)在甜菜亲本繁育中,尽量采用保纯繁殖法、循环选择繁殖法和株系循环繁殖法[5]。
[1]浙江农业大学.遗传学[M].北京:中国农业出版社,2001:284
[2]徐孟良,康公平,梁满中,等.不同世代温敏核不育水稻96-5-2S育性对低温的反应[J].作物学报,2006,32(1):20-24
[3]盖钧镒.植物种质群体遗传结构改变的测度[J].植物遗传资源学报,2005,6(1):l-8
[4]柴建方.种质库异花授粉作物种子的更新繁殖[J].作物品种资源,1996(2):38-40
[5]张天真.作物育种学总论[M].北京:中国农业出版社,2003:326-327
猜你喜欢
中老年保健(2022年2期)2022-11-25 23:46:31
中老年保健(2021年7期)2021-12-02 16:50:22
园艺与种苗(2021年10期)2021-11-28 23:31:51
中老年保健(2021年4期)2021-08-22 07:09:10
现代农业科技(2020年4期)2020-04-08 01:23:28
祝您健康(2020年1期)2020-04-02 17:17:19
中国种业(2018年8期)2018-08-28 06:40:46
新疆农垦科技(2016年1期)2016-04-12 12:04:45
农学学报(2015年9期)2015-05-30 19:06:00
种子科技(2015年8期)2015-05-24 16:14:56
