
北草蜥热适应性的生理基础
2016-11-29 02:54杜卫国
哈尔滨师范大学自然科学学报 2016年2期
沙 威, 杜卫国
(1.黑龙江省科学院自然与生态研究所;2.中国科学院动物研究所)
北草蜥热适应性的生理基础
沙 威1, 杜卫国2*
(1.黑龙江省科学院自然与生态研究所;2.中国科学院动物研究所)
以宁德和舟山两个北草蜥种群为研究对象,测定其在相同环境条件下的行为和生理的差异,推测其适应气候变化的潜在对策.在个体水平上,测定了北草蜥的热敏感性,包括不同地理种群的耐受高温(CTmax)和耐受低温(CTmin),不同温度下的疾跑速度,在生理生化水平上,结合不同温度下呼吸代谢数据来研究种群对热环境的适应.研究结果表明,不同分布区的北草蜥种群的耐受高低温差异显著,舟山蜥蜴的CTmin小于宁德个体,但其CTmax大于宁德个体.两个草蜥种群从18 ~28 ℃疾跑速度都随着环境温度的升高而增加,但从28~38 ℃过程中随温度的升高开始下降,且在相同温度下,舟山北草蜥的跑速较宁德北草蜥快.同样,两个种群的呼吸代谢率均随着温度的升高而呈增加,舟山种群的呼吸代谢水平在不同温度水平均高于宁德种群.呼吸代谢的种群间差异与个体体重差异有关,因为,去除体重差异因素后两个种群的呼吸代谢率无显著差异.
耐受高温;耐受低温;种群间差异;热敏感性;代谢率; 热适应;疾跑速度;ABSRACT
0 引言
气候变暖影响着物种遗传以及生态特性,进而可以改变群落结构以及生物多样性.近几十年来,由于人类活动影响等诸多因素,极端气候频繁发生,导致生物多样性下降.动物如何应对气候变化成为当今动物学领域的一个国际热点,一些重要成果常见于Science, Nature 等国际顶级科学刊物.掌握动物热适应性的内在本质,便成为认识动物如何应答气候变化的关键.作为外温动物,爬行类的行为和生理功能直接受到环境温度的显著影响,温度变化已经成为导致全球爬行动物多样性下降的重要原因.因此,爬行动物是研究动物如何适应温度变化的适宜对象,开展其热适应性的研究具有重要的科学价值和现实意义.该文以宁德和舟山两个北草蜥种群为研究对象,测定其在相同环境条件下的行为和生理的差异,推测其适应气候变化的潜在对策.在个体水平上,测定了北草蜥的热敏感性,包括不同地理种群的耐受高温(CTmax)和耐受低温(CTmin),不同温度下的疾跑速度,在生理水平上,结合不同温度下呼吸代谢数据来研究种群对热环境的适应.
1 材料与方法
1.1 标本采集和饲养
2012年的4~5月,分别在舟山和宁德两地采集40条成体,捕获北草蜥带回北京中国科学院动物研究所.所有的蜥蜴在实验室适应3 d,然后测量个体体长(SVL)和尾长(TL)(Mitutoyo数显游标卡尺,日本),测定体重(BM)(±0.01g)(电子天平),并进行剪趾编号,之后将同一种群的北草蜥随机分配到塑料整理箱(600 mm×450 mm×400 mm)内饲养,每个饲养箱内10~12只个体,箱子底部垫纸,每天向箱子内均匀喷水以保持饲养箱内的湿润.用纵截半片PVC管为蜥蜴提供躲避场所.饲养箱一侧顶端悬挂100W加热白炽灯泡,光周期设为12:L:12D(7:00~19:00),光周期中,饲养箱内部具有一定温度梯度,蜥蜴自行进行体温调节.饲养箱另一端摆放塑料食盘,提供充足的食物和饮水由蜥蜴随机采食.食物为黄粉虫幼虫,并且每周喂一次混有维生素的蟋蟀,每次每只蜥蜴2只蟋蟀.
1.2 热耐受性测定
2012年6月开始,在实验室内开展实验.随机从两个种群的饲养箱中分别挑选20只成体,每个种群个体雌雄各10只,每次取同一种群中的两个个体.
耐受低温(CTmin)的测定:将待测个体移入事先设置为28 ℃的生化培养箱内适应1 h,按每5 min降低箱内温度1 ℃的速率调节培养箱内的温度,直到蜥蜴个体在强烈刺激下不能做出反应(如眨眼、翻身和爬动等)时,迅速取出个体,将点温计探头快速插入蜥蜴个体的泄殖腔,根据电子点温计记录即时的泄殖腔温度,作为CTmin.蜥蜴移出培养箱后在常温进行恢复,后放回原有饲养箱内继续饲养.
耐受高温(CTmax)的测定:将待测个体放入28 ℃的恒温培养箱内适应1 h,然后从28 ℃开始,每5 min增高培养箱内温度1 ℃,直到待测个体不能翻身眨眼,在强烈刺激下不能做出反应,立即从培养箱内取出,用点温计测定其泄殖腔内的温度作为其CTmax,测定后个体在常温进行恢复,后放回原有的饲养箱内继续饲养.
1.3 运动能力测定
每个种群随机选取20只个体,在恒温室条件下测定其运动能力.所有个体分别在5个温度(18 ℃、23 ℃、28 ℃、33 ℃、38 ℃)条件下测定运动能力(疾跑速).实验在自制1800 mm×8150 mm×200 mm的木质粗糙跑道中进行,跑道一边每250mm标记,另一边为透明玻璃能使摄像机捕捉到动物的运动.实验开始前,将待测种群个体放置在设定好的测定温度的生化培养箱内适应2 h,使其适应实验测定温度.适应后,迅速将北草蜥个体从跑道一端放入,在其尾部用毛笔沿其运动方向的垂直方向驱赶,但不推动,使其跑过1.5 m的路程.同时,用数码摄像机(SONY, DCR-SR220E)拍摄其运动的过程.每个个体重复测量3次,每次测定至少间隔1 h.疾跑速度为北草蜥跑过250 mm的最大速度,将通过1.5 m的速度记为平均速度,最终数据结果以3次重复测定的平均值来表示.3次测定后将全部实验个体放回原有饲养箱内继续饲养.实验测定温度的顺序随机化,且每两个温度水平实验间至少间隔1 d.实验中对受伤个体或长时间停在跑道某处不跑或来回往返跑动的北草蜥个体进行标号记录,在之后的数据统计记录中进行排除.
1.4 呼吸代谢测定
用北草蜥宁德和舟山种群各16只成体(雌雄各8只),实验前期动物在常温实验室饲养适应.实验于6月中旬开始.每天随机选一个测定温度进行基础代谢率的实验.测定过程按Li等2010年的方法进行测定.在实验前,将随机选取的北草蜥放入塑料饲养盒中,需禁食12 h.每次选取2只放置在预先设定好温度的生化培养箱内进行1 h的热适应.利用开放气流陆生动物呼吸系统(Stable Systems International Inc., Las Vegas, NV, USA)进行代谢率测定,将适应测定温度的蜥蜴个体放入呼吸室,装入蜥蜴个体的呼吸室放置于预先设定好温度的生化培养箱内,利用干燥剂对呼吸室进出气体进行干燥,然后调定气体流速为70 mL/min,动物个体吸入的氧气量与呼出的二氧化碳量均被电脑记录,每次呼吸实验时间为30 min,每只个体进行1 h 2个循环的同一温度的呼吸测定.每个循环测定的开始和结束时都进行基线测定,所有实验测定时间均在每日8:00~18:00间进行.该实验一共设置5个不同温度水平的呼吸测定(18 ℃,23 ℃,28 ℃,33 ℃,38 ℃).实验温度顺序随机化安排,每个种群个体在当天温度水平测定完后带回常温实验室恢复一天.氧气消耗率利用下列计算公式计算:
其中FR=flow rate (mL/min), Fi=input fractional concentration (%), Fe=excurrent fractional concentration (%).然后记录连续最小的氧气含量(5 min)进行计算.体重用于计算BMR.
1.5 数据分析
所有数据在做参数统计分析前,分析检验其正态性 (Kolmogorov-Smirnov test)和方差同质性(Statistic统计软件包).经检验,所有数据符合参数统计条件.应用单因子方差分析(ANOVA),文中描述性统计值用平均值±标准误表示,显著性水平设置在α=0.05.
2 结果与分析
2.1 环境温度对热耐受性的影响
本研究中宁德北草蜥种群成体的CTmax为41.09±0.29 ℃,CTmin为3.50±0.74 ℃;舟山北草蜥种群成体的CTmax为 42.22±0.16 ℃,CTmin为2.72±0.99 ℃.处于较高纬度的舟山北草蜥种群的CTmax比较低纬度的宁德北草蜥种群要高,而CTmin又比宁德种群低.
2.2 环境温度对运动能力的影响
热环境显著影响北草蜥的运动能力,从18~33 ℃,宁德和舟山北草蜥的疾跑速度逐渐增加,在环境温度达到33 ℃时,两个地理种群的北草蜥均达到最大疾跑速,而当环境温度继续增加至38 ℃时,两个地理种群的北草蜥的疾跑速都有不同程度的下降,如图1所示.同样,两个地理种群北草蜥的平均速度也受热环境影响显著,从18~33 ℃平均速度随温度的增加而增加,当环境温度达到33 ℃时,两个地理种群个体均达到最大平均速度,当环境温度超过33 ℃时,两个地理种群的平均速度随温度的增加有不同程度的下降,如图2所示.在同一温度水平下,舟山北草蜥的疾跑速和平均速度均大于宁德北草蜥.
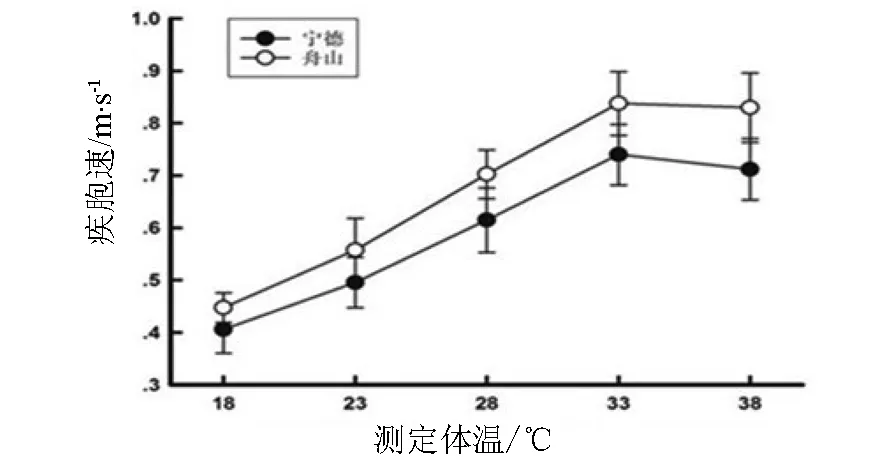
图1 宁德和舟山北草蜥的疾跑速

图2 宁德和舟山北草蜥的平均速度
2.3 环境温度对呼吸代谢的影响
在实验室条件下,热环境显著影响北草蜥的呼吸代谢率.舟山种群北草蜥的代谢率要显著高于宁德种群的北草蜥(F4,72=5.107 ,P=0.001),两个种群北草蜥在一定温度范围内,呼吸代谢随着温度的升高而递增,且在相同温度条件下,舟山种群的北草蜥的呼吸代谢率均高于宁德种群的北草蜥.去除体重因素(舟山北草蜥>宁德北草蜥),两个地理种群的北草蜥的基础呼吸率随温度的增加而升高,在同一温度条件下无显著差异(F4,72=1.488,P=0.214)如图3、图4所示.

图3 宁德和舟山种群北草蜥不同环境温度下的标准呼吸代谢率

图4 宁德和舟山种群北草蜥体重校正后的呼吸代谢率
3 讨论
爬行动物的体温受外界环境温度影响显著,不同环境地区的爬行动物种群的热耐受限度也是长期受环境温度影响的结果.有关变温动物耐受高温和耐受低温的研究已有很多,包括许多极地的鱼类,跨纬度分布的昆虫[1],不同栖息环境的蜥蜴等.综合这些变温动物实验结果分析来看,这些动物的热耐受性普遍与当地的栖息环境,特别是环境温度密切相关.高纬度地区的动物较同种低纬度分布种群的动物具有相对较低的耐受低温和相对较高的耐受高温.高纬度地区常年气候变化明显,冬季温度较低,很多地区长期低于0 ℃,而低纬度地区气候多属冬暖夏凉,全年温差波动不大.生存于高纬度地区的动物面对常年高温和低温的波动环境温度,其进化策略相对于同种低纬度地区的动物发生改变.
舟山北草蜥种群比宁德北草蜥种群有较高的耐受高温,同时又具有较低的耐受低温,生态幅相对较宽.不同地理种群分布的北草蜥种群的耐受高温和耐受低温与其栖息地气候环境密切相关,当全年不同季节温度波动较大,昼夜温差同样较大,在波动温度范围较大的热环境中生存,其机体生理机能需要对外界的高温和低温具有较好的适应.在高温气候条件下,具有较高的耐受高温,减少了为调控体温所消耗的能量,也使体内的生理过程在高温下平稳的反应.在低温环境中,具有较低的耐受低温可以较少体内热量的散失,减少了低温环境中为增加过多的产热而消耗的能量,同时,在较低的温度下,降低了体内的生理生化速率,减慢了代谢速度,使个体在较低温的环境中耗能减慢,可以避免低温季节食物不足而挨饿,能更好的使个体度过低温季节.舟山(北纬29°32′~31°04′)地区年平均气温16.5 ℃,气温波动变化大,3月至8月温度较高,约27.2 ℃,冬季温度较低,约5.6 ℃.宁德(北纬26°18′~27°4′)地区气温变化不大,夏季气温月25 ℃,冬季约16 ℃.舟山北草蜥经历比宁德种群低10 ℃的冬季温度,而夏季又要在更高的环境中生存,其栖息地的全年气候环境波动温度超过20 ℃,导致了舟山种群有很大的热生态幅,符合同种不同纬度分布种群热耐受性趋势的规律.
自然选择对动物整体功能的作用比对形态和生理功能特征的作用可能更为直接.近年来,温度对动物整体功能的作用备受关注,其中,运动能力是广泛用于评估动物整体功能的重要指标[2].因为运动能力影响动物的许多活动,其中包括逃避敌害、捕食及成体的交配,从而影响其适合度.对于爬行动物,特别是蜥蜴而言,由于是变温动物,其行为功能的实现受周围环境的限制,特别是受热环境的影响.在一定温度范围内,相对较高的体温可以使其行为性能最大化.
很多相关的实验都表明蜥蜴的运动能力受环境温度影响显著,如石龙子的运动能力在较低的温度下随着温度的上升而增强,而当温度超过34 ℃时其运动能力又开始下降.温度能够显著影响北草蜥的运动能力,在较低的温度条件下,北草蜥的运动能力随着体温的上升而增加,当环境温度超过33 ℃时又开始下降.这种现象可归因于体温在生理功能中的促进作用,尽管潜在的生理机制很复杂,但在一定范围内,相对较高的体温更有利于动物的生理活动.
在实验中,两个地理种群北草蜥在不同温度下的疾跑速度和平均速度差异显著,可能反应了不同温度水平下肌肉的生理特性以及肌肉内酶活性的差异.运动能力除了可能与自身的生理机能有关,还可能与自身形态有关.结合两个地理种群成体的形态数据分析,舟山种群北草蜥体长显著大于宁德种群北草蜥,舟山北草蜥种群的体重也显著大于宁德种群北草蜥.运动能力的种群间差异可能与其个体大小的种群间差异有关.结合当地气候环境分析,在全年温差较大的舟山地区,北草蜥每年要经受的环境温度波动较宁德地区大,每年平均经历的低温和高温气候要相对更长,因此在适应进化过程中体型较大,体重更沉,能经受住较大的环境温度波动.同时,在相对波动的环境中,其捕食、逃避的效率也对其生存发展至关重要,在更低或更高的温度条件下,如果运动能力的改变没有相对的提升,将很难提高捕食效率或逃避天敌能力,在进化中被淘汰的几率将会增大.因此,相比全年温度波动不大的宁德地区,舟山地区的北草蜥很可能通过适应进化,提升其不同温度下的运动能力来增加适合度.
爬行动物种类众多,分布广泛,一直都是比较代谢研究的重要类群.对于爬行动物能量代谢的研究不论是从生态、生理还是进化的角度都具有非常重要的意义.在当前全球温度变化的背景下,作为变温动物的爬行动物如何调整自身的适应对策是当今研究的热点问题,特别是同种不同地理分布的动物是否根据环境采取不同的对策也日益成为生态研究的重点.本实验以北草蜥为实验动物,对舟山和宁德两个种群的北草蜥进行生理代谢的实验,在5个不同的实验温度下测量,结果发现,两个地理种群的呼吸代谢率均随着环境温度的上升而增加,这种现象在爬行动物中很普遍,是大多数变温动物所采取的生理适应方式,比如丽斑麻蜥(Eremias argus)和草原沙蜥(Phrynocephalus frontalis)的代谢率随着温度的上升而上升,在多疣壁虎(Gekko japonicas)的类似研究中也得出了个体的代谢率随着温度的上升而上升的结果.北草蜥属变温动物,在一定温度范围内,其体温随环境温度的变化而平行的升高或降低,其体内的生理机能也随之发生相应的改变.北草蜥两个不同种群的基础代谢率均随温度升高而平行增加,变化趋势无显著差异.虽然在不同温度条件下舟山种群的北草蜥的代谢率均高于同温度下宁德种群的北草蜥,但是当排除体重因素,两个地理种群的平均代谢率在同一温度均无差异,表明其生理适应能力的差异不显著.舟山北草蜥大于宁德北草蜥,舟山北草蜥具有更大的表面积和身体质量,使其相对表面积减小,单位体重的热散失减少,有利于抵抗寒冷的季节.因此,不同地理分布的北草蜥可能偏向于依赖形态的适应,而生理代谢功能的适应则不明显.这对于北草蜥种群来说,避免了因环境温度的波动为自身代谢调整而导致的过多的能量消耗,有利于北草蜥在低温环境中通过形态特征的进化来弥补不利环境温度带来的冲击,而在高温条件下,又避免了基础代谢水平过度的增高而消耗更多的能量.
[1] Bale J S. Insects and low temperatures: from molecular biology to distributions and abundance[J].Philosophical transactions of the royal society,2002,29:849-862.
[2] Brana F. JiInfluence of incubation temperature on morphology, locomotor performance, and early growth of hatchling wall lizards (Podarcis muralis)[J]. Journal of Experimental Zoology,2000,286:422-433.
(责任编辑:季春阳)
The Physiological Bases of Thermal Adaptation in the northern Grass Lizard, Takydromus Septentrionalis
Sha Wei1, Du Weiguo2
(1.Institute of Natural Resources And Ecology;2.Key Laboratory of Animal Ecology and Conservation Biology, Chinese Academy of Sciences)
The behavioral and physiological differences between populations of the lizards from two geographically-separated localities of Ningde and Zhoushan are measured, and predictd its potential strategies by which the animal adapt to climate changes. At an individual level, the thermal sensitivity, including critical thermal maximum (CTmax), critical thermal minimum (CTmin), and sprint speed at different temperatures are measured in different geographic populations. The between-population differences of respiratory metabolism are also determined to examine the adaptation of this lizard in response temperature changes.
Between-population differences; Critical thermal maximum; Critical thermal minimum; Locomotor performance; Metabolic rate Thermal adaptation; Thermal sensitivity
2016-01-12
*通讯作者
Q958
A
1000-5617(2016)02-0117-05
T. septentrionalis from the two geographic localities show significant differences in CTmax and CTmin. Lizards from Zhoushan had lower CTmin, but higher CTmax than those from Ningde. The sprint speed increases with the increasing temperatures from 18 ℃ to 38 ℃, decrease at the temperature range of 28 - 38 ℃ in both populations. Moreover, lizards from Zhoushan ran faster than those from Ningde group at same temperatures. The respiratory metabolism increase as the temperature increase in both populations, with higher metabolic rate in Zhoushan lizards than Ningde lizards. However, this between-population difference in respiratory metabolism disappeares after controlling for individual weight, suggesting that the respiratory metabolism was correlated with individual weight.
猜你喜欢
世界建筑(2022年11期)2022-12-05
知识窗(2022年6期)2022-07-08
工会博览(2022年9期)2022-06-30
中老年保健(2021年10期)2021-11-30
Bone Research(2021年2期)2021-09-11
海峡姐妹(2020年6期)2020-07-25
劳动保护(2019年3期)2019-05-16
能源(2018年7期)2018-09-21
汽车与新动力(2018年2期)2018-05-09
汽车维护与修理(2016年10期)2016-07-10
