
马尾松家系苗期生长性状配合力和 杂种优势分析
2016-11-16 11:42唐效蓉李宇珂曾令文伍新云
湖南林业科技 2016年2期
唐效蓉, 李宇珂, 曾令文, 伍新云, 张 翼, 杨 骏
(1.湖南省林业科学院, 湖南 长沙 410004; 2.北京漫天下风采传媒文化有限公司, 北京 100029;3.城步苗族自治县云马国有林场, 湖南 城步 422500; 4.桂阳县林业局, 湖南 桂阳 424400)
马尾松家系苗期生长性状配合力和 杂种优势分析
唐效蓉1, 李宇珂2, 曾令文3, 伍新云4, 张 翼1, 杨 骏1
(1.湖南省林业科学院, 湖南 长沙 410004; 2.北京漫天下风采传媒文化有限公司, 北京 100029;3.城步苗族自治县云马国有林场, 湖南 城步 422500; 4.桂阳县林业局, 湖南 桂阳 424400)
以湖南省城步2代马尾松种子园人工控制授粉的全同胞家系和全同胞家系的母本与父本的自由授粉半同胞家系种子为试验材料,研究马尾松2代种子园家系苗期的配合力和杂种优势。结果表明:马尾松2代种子园家系间地径、苗高的差异均达极显著,其中地径变幅为0.215 7~0.428 8 cm,苗高变幅为13.425 9~35.568 9 cm。全同胞家系的地径、苗高与其母本的地径、苗高均呈现正相关,其中全同胞家系地径与其母本地径的相关系为0.831,为极显著相关,其它均未达显著水平。在4个杂交组合中,2个一般配合力(GCA)较高的亲本杂交后代81×8的地径均值和特殊配合力效应值均最大,分别为0.428 8 cm和0.024;2个一般配合力较高的亲本杂交后代10×37的苗高均值和特殊配合力效应值并不是最大的,而81×8组合的苗高均值和特殊配合力效应值均是最大的,分别为35.375 0 cm和3.193 4。不同组合间的杂种优势差异很大,其中地径的杂种优的变幅为-16.55%~19.41%,苗高杂种优的变幅为-46.44%~11.84%。
苗期; 生长性状; 配合力; 杂种优势; 二代种子园; 马尾松
1 试验区概况
试验地位于湖南省南部的桂阳县。该区地理位置为112.65°E、25.75°N,地貌属低山丘陵地貌;年均气温17.1~18.6 ℃,7月最高气温40.2 ℃,1月最低气温-9.5 ℃,年均日照时数1 450 h,≥10 ℃的活动积温为5 473.5 ℃,无霜期274天,年均降雨量1 400~1 600 mm,4—6月为多雨季节,冬季偏暖,雨雪较少。试验地海拔281.8~301.8 m;土壤为砂岩发育而成的红壤,土层厚度大部分在35 cm以上,pH值5.2~5.6[17]。
2 材料与方法
2.1试验材料
试验用马尾松种子为湖南省城步2代马尾松种子园人工控制授粉的全同胞家系和全同胞家系的母本、父本的自由授粉半同胞家系种子,其中全同胞家系种子17份,半同胞家系种子15份,对照为桂阳2代马尾松种子园混系种子(CK)。
2.2研究方法
2.2.1 育苗及苗木生长量调查 采用容器育苗法育苗,于2013年4月26—29日播种。2014年1月5—10日,采用完全随机区组设计,每个家系随机抽取30株,调查苗木的地径和苗高,各重复3次[17]。
2.2.2 配合力分析 按测交系统计模式对以10号、81号为母本,以8号、37号为父本的4个测交系组合进行一般配合力(GCA)和特殊配合力(SCA)分析。
2.2.3 杂种优势计算 杂种优势计算公式[21]如下:
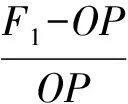
随着冠脉积分数值的增加,UA、TC、LDL明显升高,而TBIL、IBIL、DBIL、HDL均明显降低;组间比较,差异有统计学意义(P<0.05)。见表4。
式中:H——杂种优势程度;
F1——任何1个杂交组合(家系)的平均值;
OP——该F1组合中1个优势亲本的自由授粉子代的平均值。
2.2.4 数据统计与分析 采用Excel 2003和SPSS17.0进行数据计算和方差分析及相关分析。
3 结果与分析
3.1不同家系苗期生长差异性
表1结果表明:马尾松二代种子园不同家系间地径、苗高的差异均达极显著,其中地径变幅为0.215 7~0.428 8 cm,苗高变幅为13.425 9~35.568 9 cm。

表1 家系苗地径、苗高方差分析Tab1 ThevarianceanalysisofDBHandheightoffamilyseedling平方和df均方F显著性地径组间0199310006131530000组内0031640000总数023095苗高组间24629283179449223380000组内227628643557总数269055695
从表2多重比较结果可以看出:具有相同父本的81×8、137×8、10×8、123×8、86×8等5个全同胞家系平均地径大小为81×8>137×8>10×8>123×8>86×8,而其母本单系的平均地径大小为81>137>123>86>10,除10×8号家系外的其它4组全同胞家系的地径生长量与其相应母本单系生长量的变异规律相似;81×8与8号家系间差异不显著,但该2个家系与全同胞家系137×8、10×8、123×8、86×8及母本半同胞家系137、123、86、10间的差异均显著。全同胞家系平均苗高大小为81×8>137×8>123×8>10×8>86×8,其母本单系的平均苗高大小为10>81>123>137>86。全同胞家系的苗高生长量与其相应母本单系生长量的变异规律有一定的差异性。81×8与137×8号家系间差异不显著,但该2个家系与全同胞家系10×8、123×8、86×8及母本半同胞家系10、81、123、137、86间的差异均显著。
同样,从表2中还可以看出:具有相同父本的6×37、81×37、111×37、10×37等4个全同胞家系平均地径大小为6×37>81×37>111×37>10×37,而其母本单系的平均地径大小为6>81>111>10,全同胞家系的地径生长量与其相应母本单系生长量的变异规律相似;81与6×37号家系间差异不显著,但该2个家系与全同胞家系111×37、10×37及母本半同胞家系111、10间的差异均显著。全同胞家系平均苗高大小为111×37>10×37>6×37>81×37,其母本单系的平均苗高大小为111>10>81>6。除6号家系外的其它3组全同胞家系的苗高生长量与其相应母本单系生长量的变异规律均相似。111×37与全同胞家系81×37及母本半同胞家系81、6号家系间的差异均显著。
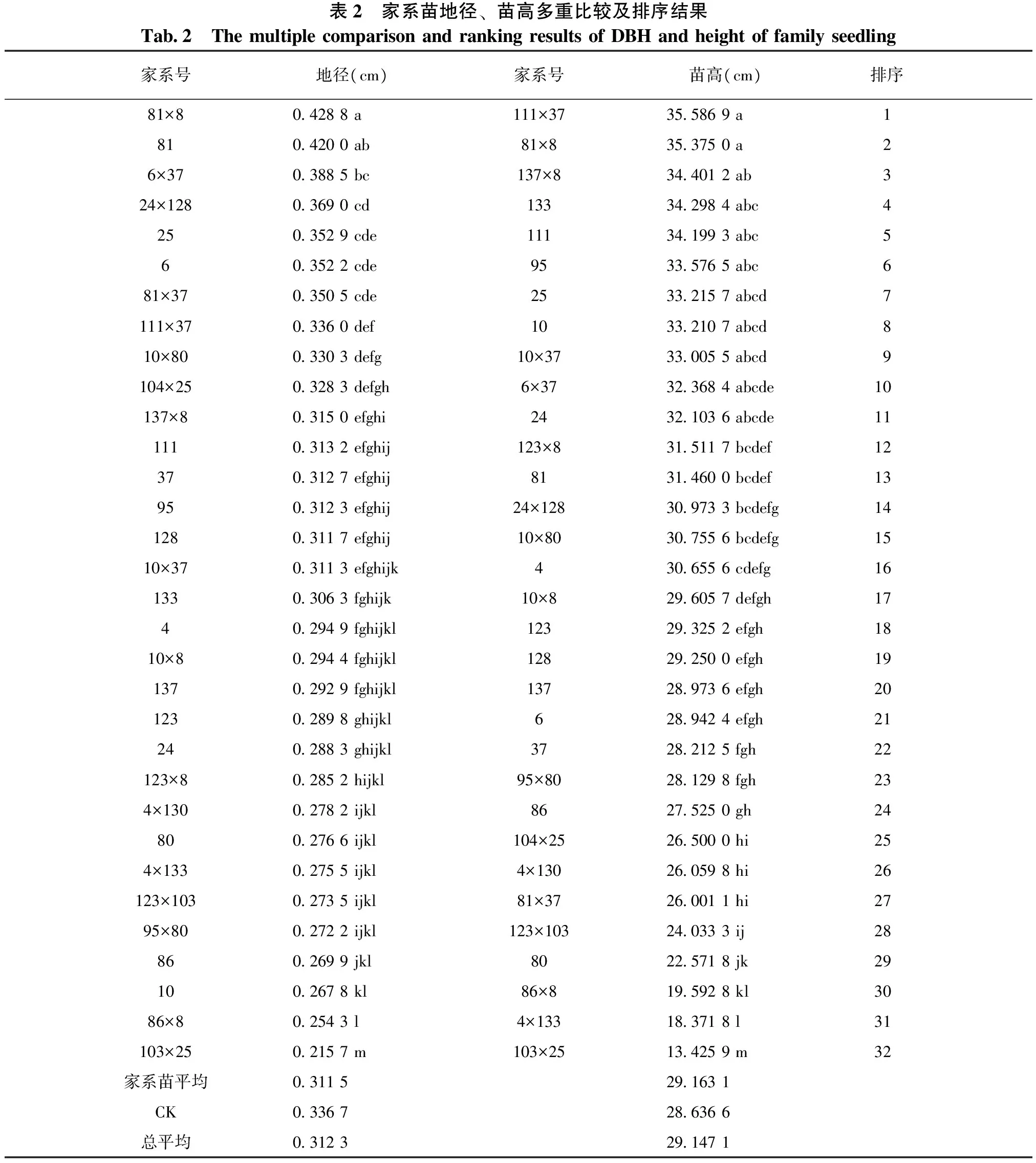
表2 家系苗地径、苗高多重比较及排序结果Tab2 ThemultiplecomparisonandrankingresultsofDBHandheightoffamilyseedling家系号地径(cm)家系号苗高(cm)排序81×804288a111×37355869a18104200ab81×8353750a26×3703885bc137×8344012ab324×12803690cd133342984abc42503529cde111341993abc5603522cde95335765abc681×3703505cde25332157abcd7111×3703360def10332107abcd810×8003303defg10×37330055abcd9104×2503283defgh6×37323684abcde10137×803150efghi24321036abcde1111103132efghij123×8315117bcdef123703127efghij81314600bcdef139503123efghij24×128309733bcdefg1412803117efghij10×80307556bcdefg1510×3703113efghijk4306556cdefg1613303063fghijk10×8296057defgh17402949fghijkl123293252efgh1810×802944fghijkl128292500efgh1913702929fghijkl137289736efgh2012302898ghijkl6289424efgh212402883ghijkl37282125fgh22123×802852hijkl95×80281298fgh234×13002782ijkl86275250gh248002766ijkl104×25265000hi254×13302755ijkl4×130260598hi26123×10302735ijkl81×37260011hi2795×8002722ijkl123×103240333ij288602699jkl80225718jk291002678kl86×8195928kl3086×802543l4×133183718l31103×2502157m103×25134259m32家系苗平均03115291631CK03367286366总平均03123291471
对以上具有相同父本的2组共9个全同胞家系的地径、苗高与其对应母本的地径、苗高进行遗传相关分析。结果表明,全同胞家系的地径、苗高与其母本的地径、苗高均呈现正相关,其中全同胞家系地径与其母本地径的相关系为0.831,为极显著相关,其它均未达显著水平。
据以上结果,有必要进行配合力和杂种优势分析。
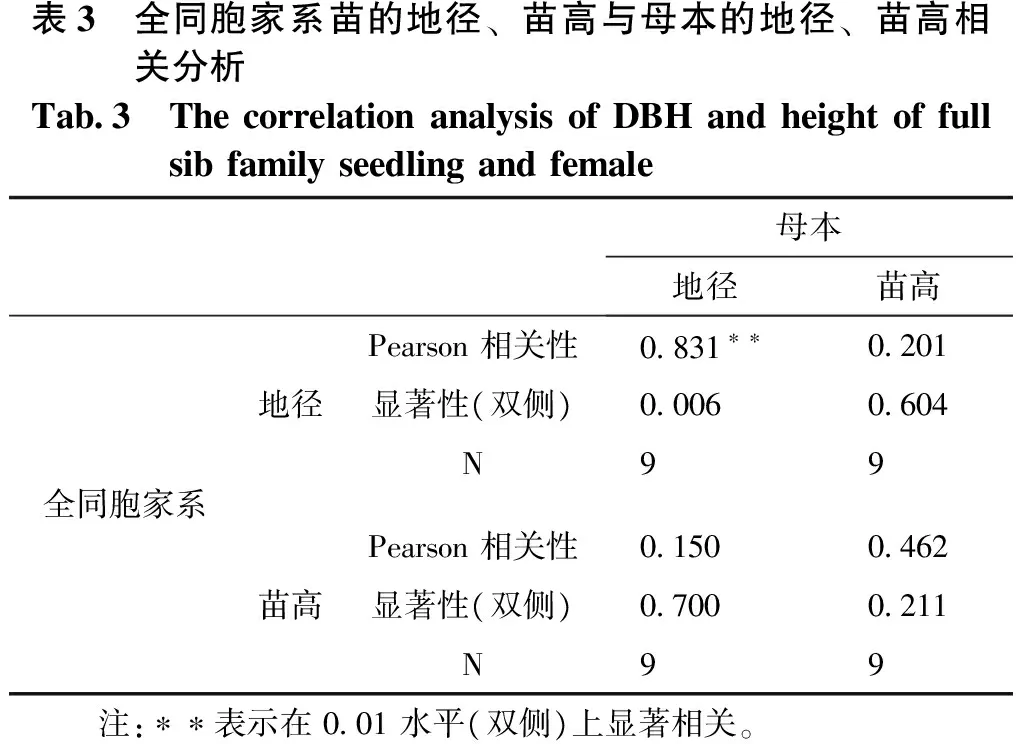
表3 全同胞家系苗的地径、苗高与母本的地径、苗高相关分析Tab3 ThecorrelationanalysisofDBHandheightoffullsibfamilyseedlingandfemale母本地径苗高全同胞家系地径Pearson相关性0831∗∗0201显著性(双侧)00060604N99苗高Pearson相关性0150 0462显著性(双侧)07000211N99 注:∗∗表示在001水平(双侧)上显著相关。
3.2不同测交组合的配合力
一般配合力(GCA)和特殊配合力(SCA)计算结果表明:以10号、81号为母本和以8号、37号为父本的4个测交系组合中,81号母本地径的一般配合力效应值(0.043)较10号的(-0.043)大,8号父本地径的一般配合力效应值(0.015)较37号的(-0.015)大,2个一般配合力(GCA)较高的亲本杂交后代81×8的地径均值和特殊配合力效应值均最大,分别为0.428 8 cm和0.024。10号母本苗高的一般配合力效应值(0.308 7)较81号的(-0.308 7)大较,8号父本苗高的一般配合力效应值(1.493 5)较37号的(-1.493 5)大,但2个一般配合力较高的亲本杂交后代10×37的苗高均值和特殊配合力效应值不是最大的;81×8组合的苗高均值和特殊配合力效应值均是最大的,分别为35.375 0 cm和3.193 4。
3.3苗期杂种优势
由表3可知:不同组合间的杂种优势差异很大,其中地径的杂种优的变幅为-16.55%~19.41%,苗高杂种优的变幅为-46.44%~11.84%;6×37和111×37的地径和苗高以及24×128、10×80的地径均具有明显的杂种优势,其中24×128、10×80地径的杂种优势均在15%以上,6×37的地径和苗高的杂种优势均在10%以上。其它均呈现一定的杂种劣势,其中95×80、81×37、4×133等3个组合的地径和苗高的杂种劣势均在10%以上;特别是4×133,其苗高的杂种劣势达46.4%。这表明只有少数亲本组合的生长表现优良,多数亲本组合的生长表现一般甚至很差。
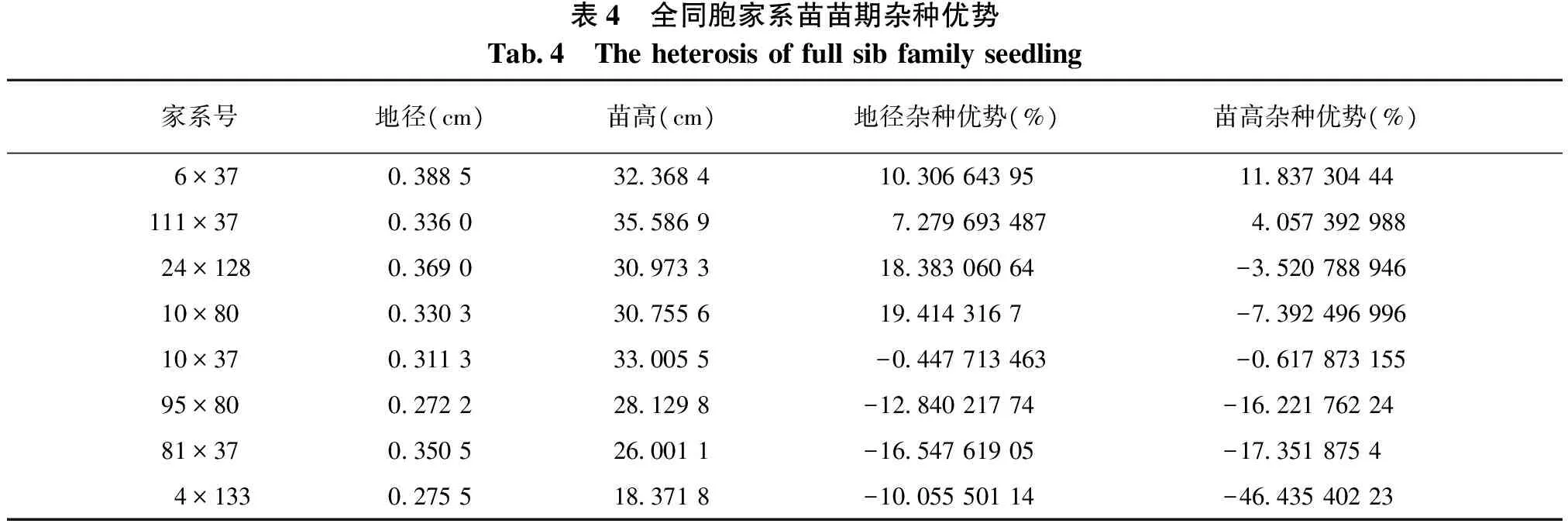
表4 全同胞家系苗苗期杂种优势Tab4 Theheterosisoffullsibfamilyseedling家系号地径(cm)苗高(cm)地径杂种优势(%)苗高杂种优势(%)6×370388532368410306643951183730444111×37033603558697279693487405739298824×128036903097331838306064-352078894610×8003303307556194143167-739249699610×3703113330055-0447713463-061787315595×8002722281298-1284021774-162217622481×3703505260011-1654761905-1735187544×13302755183718-1005550114-4643540223
4 结论与讨论
(1)马尾松二代种子园家系间地径、苗高的差异均达极显著,其中地径变幅为0.215 7~0.428 8 cm,苗高变幅为13.425 9~35.568 9 cm。全同胞家系的地径、苗高与其母本的地径、苗高均呈现正相关,其中全同胞家系地径与其母本地径的相关系为0.831,为极显著相关,其它均未达显著水平。这意味着杂交后代苗期变异丰富,苗期选择可作为创制和筛选下世一代优良杂交组合的参考,并可为进一步的早晚相关研究提供基础数据。
(2)在4个杂交组合中,2个一般配合力(GCA)较高的亲本杂交后代81×8的地径均值和特殊配合力效应值均最大,分别为0.428 8 cm和0.024;2个一般配合力较高的亲本杂交后代10×37的苗高均值和特殊配合力效应值并不是最大的,而81×8组合的苗高均值和特殊配合力效应值均是最大的,分别为35.375 0 cm和3.193 4。一般配合力和特殊配合力是评价亲本组配能力的2个主要指标[23]。以一般配合力较高的亲本进行杂交,会产生强杂种优势,会提高得到杂种优势的机率,但无一定规律可寻[23],即各性状的GCA和SCA无明显的对应关系[23]。
(3)不同组合间的杂种优势差异很大,与周志春的研究结果[20]一致。其中地径的杂种优的变幅为-16.55%~19.41%,苗高杂种优的变幅为-46.44%~11.84%。只有少数亲本组合的生长表现优良,多数亲本组合的生长表现一般甚至很差,与张一等的研究结果[20]一致。
[1] 张一,周志春,金国庆,等.马尾松双列杂交亲本遗传距离与杂种优势相关性分析[J].南京林业大学学报,2010,34(1):9-14.
[2] 王明庥.林木遗传育种学[M].北京:中国林业出版社,2001.
[3] 李午平,伍家荣,李东.马尾松生长性状的早晚期相关与早期预测[J].湖南林业科技,1992,19(3):21-24,8.
[4] 杨章旗.马尾松优良家系选择的适宜年龄研究[J].中南林学院学报,2004,24(4):24-27.
[5] 陈亚斌.马尾松半同胞家系多年度测定与早期选择[J].林业技术开发,2011,25(1):115-118.
[6] 郑仁华.马尾松早期选择的研究[J].福建林学院学报,1999,19(4):368-371.
[7] 廖纯茂,李明贵,何晓红,等.福建柏种源试验苗期选择研究[J].湖南林业科技,1998,25(3):5-11.
[8] 张新华.福建柏优树家系苗期生长性状遗传变异和选择研究[J].湖南林业科技,2003,30(4):19-21.
[9] 曹惠清.湿地松、火炬松与马尾松苗期生长对比试验[J].四川林业科技,1985(1):44-46.
[10] 梁仰贞.湿地松、火炬松与马尾松苗期生长的差异[J].林业科技通讯,1984(2):11-13.
[11] 沈国华,金韵琴.不同种源马尾松苗期试验小结[J].江苏林业科技,1978(1):5-30.
[12] 红安县林科所,黄冈地区林科所.不同种源马尾松苗期试验初报[J].湖北林业科技,1979(2):37-41.
[13] 汪企明.马尾松不同种源苗期生长和生物量的初步研究[J].林业科技通讯,1988(2):3-5.
[14] 温佐吾.广西、贵州两个马尾松种源在新西兰的苗期生长情况比较[J].广西林业科学,1995,24(3):145-148.
[12] 林建东.马尾松一代种子园不同家系半同胞子代苗期生长分析[J].安徽农学通报,2011,17(17):130-134.
[15] 福建省马尾松高产脂类型研究协作组.高产脂马尾松苗期和幼林的生长分析试验报告[J].福建林业科技,1987(1):13-19.
[16] 秦国峰,王培蒂,周志春.马尾松苗期生长性状的遗传分析[J].林业科学,1989,25(6):559-563.
[17] 伍新云,王晓锋,唐效蓉.不同种源的马尾松半同胞家系子代苗期生长表现[J].湖南林业科技,2015,42(2)10-14.
[18] 陈岳武,施季森,刘大林,等.杉木种内杂种优势及亲本配合力的分析[J].南京林产工业学院学报,1982(2):1-19.
[19] 徐清乾,吴成田,许忠坤.杉木全双列子代遗传变异分析[J].湖南林业科技,2015,42(6):7-12.
[20] 周志春,金国庆,秦国峰.马尾松纸浆材重要经济性状配合力及杂种优势分析[J].林业科学,2004,40(4):52-57.
[21] 金国庆,秦国峰,刘伟宏,等.马尾松生长性状交配效应的遗传分析及杂交组合选择[J].林业科学,2008,44(6):28-33.
[22] 黄伟.马尾松全同胞杂交育种试验初报[J].安徽农学通报,2012,18(2):92,105.
[23] 刘青华,金国庆,储德裕,等.基于马尾松测交子代的生长、干形和木材密度的配合力分析[J].南京林业大学学报,2011,35(2):8-14.
TheanalysisongrowthtraitscombiningabilityandheterosisofPinusmassonianafamilywithseedling
TANG Xiaorong1, LI Yuke2, ZENG Lingwen3, WU Xinyun4, ZHANG Yi1, YANG Jun1
(1.Hunan Academy of Forestry, Changsha 410004, China; 2.Beijing Mantianxia Culture Co. Ltd, Beijing 100029, China; 3.Yunma State-owned Forest Farm of Chengbu Miao Autonomous County, Chengbu 422500, China; 4.Forestry Bureau of Guiyang County, Guiyang 424400, China)
Taking the artificial control pollination of full sib and full sib families of female parent and male parent of open pollinated half sib seeds ofPinusmassonianaof the Chengbu Second Generation Seed Orchard ofPinusmassonianaof Hunan Province as the test materials,to research the combining ability and heterosis ofP.massonianafamily with seedling in the orchard.The results showed that:there is a significant difference in growth characters of DBH and height ofP.massonianain the difference of the second generation seed orchard,of which the DBH ranged from 0.215 7~0.428 8 cm,the seedling height ranged from 13.425 9 to 35.568 9 cm.The DBH and height of full sib family and its female parent showed positive correlation,and the DBH related situation of full sib family and its female parent is 0.831,and in significant difference and others were not up to significant level.In four hybrid combinations,two higher general combining ability(GCA)crosses 81×8 to size mean and special effect value of combining ability were the biggest,it’s 0.428 8 cm and 0.024 respectively;two higher general combining ability of hybrid offspring of parents 10×37 seedling height mean and special effect value of combining ability and not the biggest.The 81×8 combinations of seedling height mean and special effect value of combining force were the largest,it’s 35.375 0 cm and 3.193 4 respectively.The heterosis among different combinations is significant difference,the DBH of heterosis ranged from -16.55% to 19.41%,and height of heterrsis ranged from -46.44% to 11.84%.
seedling; growth traits; combining ability; heterosis; second generation seed orchard; Pinus massoniana
2016-01-25
“十二五”国家科技支撑计划课题“南方针叶树种高世代育种技术研究与示范”(2012BAD01B02)。
唐效蓉(1964-),女,湖南省邵东县人,研究员,主要从事林木遗传育种与栽培技术研究。
S 791.248
A
1003 — 5710(2016)02 — 0008 — 05
10. 3969/j. issn. 1003 — 5710. 2016. 02. 002
(文字编校:唐效蓉)
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
蔬菜(2021年7期)2021-11-27
农技服务(2020年1期)2020-12-17
山东农业科学(2019年11期)2019-12-24
现代农业科技(2019年1期)2019-07-11
中国种业(2018年11期)2018-11-16
猪业科学(2018年5期)2018-07-17
现代农业科技(2017年24期)2018-01-22
种子科技(2014年8期)2014-05-18
种子科技(2014年9期)2014-01-22
