
红花玉兰大小孢子发生及雌雄配子体发育的研究
2016-11-10 05:18陈丽园桑子阳陈发菊马履一
西北农林科技大学学报(自然科学版) 2016年9期
陈丽园,桑子阳,陈发菊,马履一
(1 北京林业大学 森林培育与保护教育部重点实验室,北京 100083;2 三峡大学 生物技术研究中心,湖北 宜昌 443002)
红花玉兰大小孢子发生及雌雄配子体发育的研究
陈丽园1,桑子阳1,陈发菊2,马履一1
(1 北京林业大学 森林培育与保护教育部重点实验室,北京 100083;2 三峡大学 生物技术研究中心,湖北 宜昌 443002)
【目的】 对红花玉兰(Magnoliawufengensis)大、小孢子发生及雌、雄配子体发育过程进行显微观察,探讨其自然状态下结籽率低的原因。【方法】 于2012年5月红花玉兰花芽分化开始每月采花芽1次,直至次年开花,采用常规石蜡切片法制片,对红花玉兰大、小孢子发生及雌、雄配子体发育进行详细的细胞学观察。【结果】 红花玉兰雌蕊成熟时倒生胚珠,双珠被,厚珠心,大孢子为单胞子发生型,成熟胚囊为7胞8核胚囊,蓼型胚囊发育方式。雄蕊花药四室,腺质型绒毡层,成熟花粉粒为二细胞型。【结论】 红花玉兰大、小孢子发生以及雌、雄配子体发育过程中未见异常现象,因此该物种雌、雄蕊发育不是导致其濒危的因素。
红花玉兰;大孢子发生;小孢子发生;雌、雄配子体发育;细胞学观察
红花玉兰(MagnoliawufengensisL.Y.Ma et L.R.Wang)属木兰科木兰属玉兰亚属,主要分布在湖北省五峰县西南部,生长于海拔1 000~1 500 m的山坡[1-2]。红花玉兰性状原始,自然变异丰富,具有很高的研究价值。该树种树形优美,花大芬芳,具有极高的观赏利用价值。但是由于该物种种群分布区域狭小,加上大量采挖,生境破坏严重,导致其数量稀少,处于濒危状态。
对木兰科几个种的胚胎发育过程进行研究发现,生殖发育中的异常现象成为导致其濒危的重要因素,如对西畴含笑的研究发现,其花粉胚胎发育异常,授粉昆虫少且近亲繁殖[3];香港木兰在空气湿度低的条件下,单核花粉败育[4];观光木在阳光不足的条件下花粉大量败育[5],而且大孢子发生与雌配子体发育各个阶段都会出现败育现象,导致其濒危[6];广西含笑在小孢子母细胞减数分裂时期败育[7];凹叶厚朴的大、小孢子和雌、雄配子体都会部分败育[8]。然而,对焕镛木[9]、天女木兰[10]和红花木莲[11]的研究发现,生殖生物学变化不是造成树种濒危的主要原因。近年来,对红花玉兰的研究主要集中在种质资源保护、抗逆性、遗传多样性以及花发育调控等[12-15]方面,而对该物种胚胎发育的细胞学研究未见报道。因此,本研究采用常规石蜡切片技术,对红花玉兰大、小孢子发生及雌、雄配子体发育进行了详细的细胞学观察,拟从生殖生物学角度分析该物种结实率低的原因,为该物种的保护提供决策依据。
1 材料与方法
1.1材料
用于红花玉兰胚胎学研究的材料采自湖北五峰县梨子坪,从2012年5月花芽分化开始,每月采花芽1次,直至次年开花,每次采5个花芽于FAA溶液(体积分数70%乙醇90 mL,醋酸5 mL,福尔马林5 mL)中固定24 h,然后转入体积分数70%乙醇中保存备用。
1.2方法
采用石蜡切片法[16]制片,切片厚度8 μm,连续切片,用番红-固绿(Safranin-Fast Green)对染法染色[17],脱水透明,加拿大树胶封片,最后于Nikon Eclipse 80i显微镜下观察并照相。
2 结果与分析
2.1红花玉兰大孢子发生和雌配子体发育
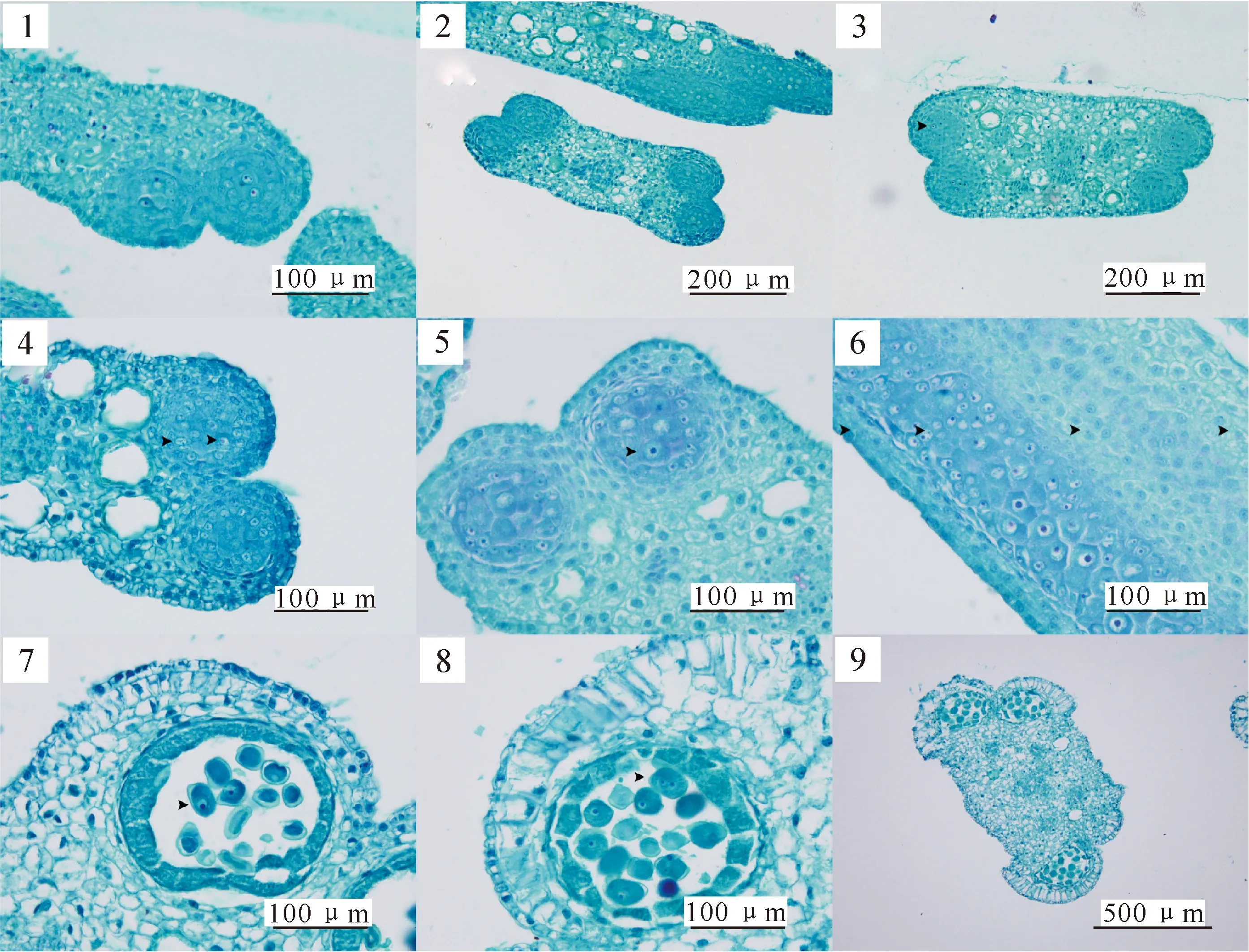
红花玉兰大孢子的发生和雌配子体的发育见图版Ⅰ。
红花玉兰具淡紫红色的离心皮雌蕊群,呈螺旋状排列(图版Ⅰ-1)。1个雌蕊由1个类似叶片的心皮向内折叠卷曲,与边缘连接形成一室子房,并在边缘形成胎座,其表皮下层细胞分裂产生突起,形成一团珠心组织,即初生的胚珠原基,珠心组织中间产生1个孢原细胞(图版Ⅰ-2),继而通过有丝分裂(图版Ⅰ-3)形成1个初生周缘细胞和1个初生造孢细胞(图版Ⅰ-4)。在图版Ⅰ-5中可见红花玉兰的子房具1室,每室2个胚珠,每个胚珠含1个大孢子母细胞。
胚珠近珠孔端的初生周缘细胞经过垂周和平周分裂形成2~4层细胞,将合点端由初生造孢细胞发育的大孢子母细胞包裹在其中(图版Ⅰ-6)。大孢子母细胞经过2次减数分裂形成大孢子四分体,其中合点端的1个成为功能大孢子,而其余3个退化,随后进入胚囊发育阶段。
胚囊发育之初,功能大孢子的体积开始增大,同时出现了液泡,即单核胚囊(图版Ⅰ-7)。第1次有丝分裂后形成2个游离核(图版Ⅰ-8),分别移到胚囊两极,中间无细胞壁,即二核胚囊。第2次有丝分裂后形成四核胚囊,第3次有丝分裂后共生成8个游离的核,4个在珠孔端,4个在合点端;在珠孔端与合点端的4个核中,各有1个核向胚囊中心靠拢(图版Ⅰ-9),融合成为中央极核。与此同时,核之间出现细胞膜,把胚囊隔成7个细胞,中央极核同处于一个中央细胞(图版Ⅰ-10);合点端的另外3个细胞聚在一起成为反足细胞;珠孔端的3个核形成2个助细胞(图版Ⅰ-11),与长梨形卵细胞组成卵器(图版Ⅰ-12)。从大孢子发生到雌配子体发育完成,胚珠的珠柄与珠被的不对称生长使胚珠的珠孔一端逐渐向下方弯转,形成倒生型胚珠,双珠被,厚珠心。红花玉兰大孢子发生与雌配子体发育的特点与木兰科其他植物的细胞学特征[18]一致。
2.2红花玉兰小孢子发生和雄配子体发育
红花玉兰雄蕊多数,离生,螺旋状排列,花丝极短,基部着生在圆锥状的花托上,上部花药原基的下表皮细胞体积增大,细胞核明显且富含细胞质(图版Ⅱ-1);4个角隅细胞快速分裂形成4个瓣裂(图版Ⅱ-2),后期发育成4个花粉囊,在近轴面呈线性排列。每一瓣表皮下分化出孢原细胞(图版Ⅱ-3),较其他细胞大,径向延长并有显著的细胞核。孢原细胞进行一次平周分裂,形成2个细胞,外面的是初生周缘细胞,里面是初生造孢细胞(图版Ⅱ-4)。经过有丝分裂形成次生造孢细胞(图版Ⅱ-5),并进一步发育为小孢子母细胞。
初生周缘细胞进行一次平周分裂,生成两层次生周缘细胞,外层发育为药室内壁,即纤维层;内层再进行一次平周分裂,所形成的两层细胞分别发育为中层和绒毡层(图版Ⅱ-6),在雄配子体的整个形成过程中提供充足的营养。小孢子形成初期,绒毡层为单核,细胞质浓厚并紧贴小孢子母细胞,在小孢子发育时期绒毡层细胞十分活跃,可见其细胞核多分裂为二核或多核。小孢子母细胞经过减数分裂形成四分体小孢子,被共同的胼胝质壁包围并分隔,随着胼胝质壁溶解消失,小孢子释放到药室中,即单核花粉(图版Ⅱ-7)。这时的绒毡层已经开始解体,小孢子体积增大,细胞质液泡化,细胞壁加厚,核转移到细胞壁边,在这里又进行一次有丝分裂。接着发生胞质分裂,大液泡、大部分细胞质和营养核被围在同一个细胞内,形成二细胞型花粉而生殖核呈梭状贴于细胞壁上(图版Ⅱ-8),仅含少量细胞质。此时中层贮藏的营养物质逐渐被吸收而解体,生殖细胞逐渐脱离花粉内壁,游离在营养细胞的细胞质中(图版Ⅱ-9)。当花药发育成熟时,药室内壁细胞径向延长并呈带状加厚,加厚的物质主要是纤维素,绒毡层几乎消失殆尽,继而相邻2个小孢子囊连接处裂开,花粉散出。

图版Ⅰ红花玉兰大孢子的发生和雌配子体的发育
1. 雌蕊纵切;2.孢原细胞;3.孢原细孢有丝分裂;4.初生造孢细胞和初生周缘细胞;5.子房1室,2胚珠;6.大孢子母细胞;
7.单核胚囊;8.单核胚囊有丝分裂后期;9.未融合的中央细胞;10.反足细胞和中央细胞;11.2个助细胞;12.卵器
Plate ⅠMegasporogenesis and female gametophyte development ofMagnoliawufengensis
1.Pistil;2.Sporogonium;3.Mitosis of sporogonium;4.Primary sporogenous cell and primary parietal cell;5.Ovary with 1 room and 2 ovules;6.Megasporocyte;7.Mononuclear embryo sac;8.Mitosis anaphase of mononuclear embryo sac;9.Nuclear-merging central cell;10.Antipodal cell and central cell;11.2 synergid cells;12.Egg apparatus
图版Ⅱ红花玉兰小孢子的发生和雄配子体的发育
1.花药原基下表皮细胞;2.花药原基的4个角隅;3.花药横切,4个花粉囊和药隔,孢原细胞;4.初生周缘细胞和初生造孢细胞;5.次生造孢细胞;6.从外到内依次为表皮、纤维层、中层、绒毡层;7.单核花粉;8.二细胞型花粉,生殖核;9.成熟花粉囊
Plate ⅡMicrosporogenesis and male gametophyte development ofMagnoliawufengensis
1.Anther primordia epidermal cells;2.Anther primordial 4 corners;3.Anther,4 pollen sac and sporogonium;4.Primary sporogenous cell and primary parietal cell;5.Secondary sporogenous cell;6.From outside to inside:epidermis,ffibrous layer,middle lamella,tapetum;7.Mononuclear pollen;8.Two-cellulae pollen;9.Mature pollen sac
3 讨论与结论
从对红花玉兰大、小孢子发生和雌、雄配子体形成过程的大量切片观察结果中发现,红花玉兰胚珠倒生,双珠被,厚珠心,四分体呈直线排列,合点端为功能大孢子,胚囊为单孢子发生类型,胚囊的发育方式与木兰科其他植物的胚囊发育类型相同,属于原始的发育类型[11,18-22]。在同科植物香木莲和红花木莲有性生殖过程研究中发现,红花木莲成熟胚囊的卵细胞败育率高达79.1%[18];香木莲功能大孢子退化率为27.9%,受精前卵器退化率高达80%[20]。
红花玉兰的花药有4个花粉囊。在花粉发育过程中,花药壁由外到内依次为表皮、药室内壁、中层和绒毡层,但是当花粉发育成熟时,药室内壁细胞纤维化并增厚,最终破裂并释放出成熟的二核细胞的花粉,这些发育特征与香木莲、红花木莲、灰木莲以及华木莲等木兰科植物基本相同[18-20,22]。但红花玉兰的绒毡层仅由一层大型多核细胞构成,与巴东木莲和华木莲[22-23]一致,而与香木莲、红花木莲、灰木莲由多层细胞构成的绒毡层不同[11,19-20],这种发育上的差异值得进一步研究。
由于在红花玉兰大、小孢子发生发育以及雌、雄配子体形成过程中,未观察到胚珠败育、卵器退化和小孢子败育等异常现象,因此红花玉兰大、小孢子发生发育以及雌、雄配子体的形成过程对该物种的结实影响不大,其结实率低及濒危原因有待进一步研究。
[1]马履一,王罗荣,贺随超,等.中国木兰科木兰属一新种 [J].植物研究,2006,26(1):4-7.
Ma L Y,Wang L R,He S C,et al.A new species ofMagnolia(Magnoliaceae) from Hubei,China [J].Bull Bot Res,2006,26(1):4-7.
[2]马履一,王罗荣,贺随超,等.中国木兰科木兰属一新变种 [J].植物研究,2006,26(5):516-519.
Ma L Y,Wang L R,He S C,et al.A new variety ofMagnolia(Magnoliaceae) from Hubei,China [J].Bull Bot Res,2006,26(5):516-519.
[3]Zhao X F,Sun W B.Abnormalities insexual development and pollinator limitation inMicheliacoriacea(Magnoliaceae),a critically endangered endemic to Southeast Yunnan,China [J].Flora,2009,204:463-470.
[4]王亚玲,张寿洲.香港木兰小孢子发生及雄配子体发育的研究 [J].武汉植物研究,2008,26(6):547-553.
Wang Y L,Zhang S Z.Studies on the microsporogenesis and development of the male gametophyte ofMagnoliachampioniiBenth. [J].Journal of Wuhan Botanical Research,2008,26(6):547-553.
[5]付琳,徐凤霞,曾庆文.观光木的小孢子发生及雄配子体发育的研究 [J].热带亚热带植物学报,2009,17(5):419-426.
Fu L,Xu F X,Zeng Q W.Studies on microsporogenesis and development of male gametophyte inTsoongiodendronodorum[J].Journal of Tropical and Subtropical Botany,2009,17(5):419-426.
[6]唐源江,叶秀麟,曾庆文,等.观光木的大孢子发生与雌配子体形成 [J].热带亚热带植物学报,2003,11(1):20-22.
Tang Y J,Ye X L,Zeng Q W,et al.Studies on megasporogenesis and development of female gametophyte inTsoongiodendronodorum[J].Journal of Tropical and Subtropical Botany,2003,11(1):20-22.
[7]付琳,徐凤霞,曾庆文,等.广西含笑的小孢子发生及雄配子体形成的研究 [J].广西植物,2011,31(3):312-317.
Fu L,Xu F X,Zeng Q W,et al.Studies of microsporogenesis and male gametophyte formation ofMicheliaguangxiensis[J].Guihaia,2011,31(3):312-317.
[8]王利琳,胡江琴,庞基良,等.凹叶厚朴大小孢子发生和雌雄配子体发育的研究 [J].实验生物学报,2005,38(6):490-500.
Wang L L,Hu J Q,Pang J L,et al.Studies on the megasporogenesis and microsporogenesis and the development of their female and male gametophyte inMagnoliabiloba[J].Acta Biologiae Experimentalis Sinica,2005,38(6):490-500.
[9]谭金桂,吴鸿,李勇,等.焕镛木小孢子发生及雄配子体发育研究 [J].西北植物学报,2009,29(5):937-944.
Tan J G,Wu H,Li Y,et al.Microsporogenesis and male gametophyte formation ofWoonyoungiaseptentrionalis(Dandy) Law. [J].Acta Bot Boreal-Occident Sin,2009,29(5):937-944.
[10]张风娟,徐兴友,陈凤敏,等.天女木兰小孢子发生及雄配子体发育的观察 [J].经济林研究,2008,26(4):71-75.
Zhang F J,Xu X Y,Chen F M,et al.Observation of microsporogenesis and male gametophyte development inMagnoliasieboldiiK. [J].Nonwood Forest Research,2008,26(4):71-75.
[11]潘跃芝,梁汉兴,龚洵.濒危植物红花木莲小孢子发生和雄配子体发育的研究 [J].云南植物研究,2001,23(1):85-90.
Pan Y Z,Liang H X,Gong X.Studies on the formation of microspores and development of male gametes inManglietiainsignis[J].Acta Botanica Yunnanica,2001,23(1):85-90.
[12]桑子阳.红花玉兰花部性状多样性分析与抗旱性研究 [D].北京:北京林业大学,2011.
Sang Z Y.Diversity analysis of the floral traits and study on the drought resistance ofMagnoliawufengensis[D].Beijing:Beijing Forestry University,2011.
[13]Wu W T,Chen F J,Jing D L,et al.Isolation and characterization of an agamous-like gene fromMagnoliawufengensis(Magnoliaceae) [J].Plant Mol Biol Report,2012,30:690-698.
[14]Jing D L,Liu Z X,Zhang B,et al.Two ancestral APETALA3 homologs from the basal angiospermManoliawufengensis(Magnoliaceae) can affect flower development ofArabidopsis[J].Gene,2014,537:100-107.
[15]Chen L Y,Chen F J,He S C,et al.High genetic diversity and small genetic variation among populations ofMagnoliawufengensis(Magnoliaceae),revealed by ISSR and SRAP markers [J].Electronic Journal of Biotechnology,2014,17(6):268-274.
[16]李正理.植物组织制片学 [M].北京:北京大学出版社,1996:129-137.
Li Z L.Plant tissue sectioning technology [M].Beijing:Peking University Press,1996:129-137.
[17]胡宝忠,常缨. 植物学实验 [M].北京:中国农业出版社,2005:24-26.
Hu B Z,Chang Y.Botany experiments [M].Beijing:China Agriculture Press,2005:24-26.
[18]潘跃芝,龚洵.濒危植物红花木莲大孢子发生和雌配子体发育的研究 [J].西北植物学报,2002,22(5):1209-1214.
Pan Y Z,Gong X.Megasporegenesis,development of female gametophyte ofManglietiainsignis(Magnoliaceae) [J].Acta Bot Boreal-Occident Sin,2002,22(5):1209-1214.
[19]Liao J P,Chen Z L,Cai X C.Embryo ofManglietiaglaucavar.sumatranaandMicheliaguangxisisand the abnormal development [C]//Proc Internat Symp Fam Magnoliaceae.Beijing:Science Press,2000:177-187.
[20]Pan Y Z,Liang H X,Gong X.Studies on the reproductive biology and endangerment mechanism of the endangered plantManglietiaaromatic[J].Acta Botanica Sinica,2003,45(3):311-316.
[21]Xiao D X,Xu F X.Megasporogenesis and development of female gametophyte inManglietiadecidua(Magnoliaceae) [J].Annales Botanici Fennici,2006,43:437-444.
[22]肖德兴,俞志雄.华木莲花药的发生发育 [J].热带亚热带植物学报,2004,12(4):309-312.
Xiao D X,Yu Z X.Anther development inSinomanglietiaglauca(Magnoliaceae) [J].Journal of Tropical and Substropical Botany,2004,12(4):309-312.
[23]Chen F J,Li F L,Liang H W,et al.Megasporogenesis,microsporogenesis and development of gametophytes in the rare endangered plantManglietiapatungensisHu [J].Bull Bot Res,2008,28(6):657-662.
Sporogenesis and gametophytes development ofMagnoliawufengensis
CHEN Liyuan1,SANG Ziyang1,CHEN Faju2,MA Lüyi1
(1KeyLaboratoryforSilvicultureandConservationoftheMinistryofEducation,BeijingForestryUniversity,Beijing100083,China;2BiotechnologyResearchCenter,ChinaThreeGorgesUniversity,Yichang,Hubei443002,China)
【Objective】 The genesis of megaspore,microspore,and development of male and female gametophyte populations ofMagnoliawufengensiswere studied to understand the causes of natural regeneration difficulties.【Method】 The genesis of megaspore and microspore, and development of male and female gametophyte were observed and studied by cytology observation ofM.wufengensisparaffin section every month from May 2012 to flowering of next year.【Result】 Characteristics of matureM.wufengensispistil include anatropous ovule,two layers of integument,thick nucellus,single-spore megasporogenesis,and seven-cell structured polygonum embryo sac assembling with eight nuclei.Stamens have four roomed anthers,glandular tapetum,and two-cellular mature pollen.【Conclusion】 Microsporogenesis and development of female and male gametophytes ofM.wufengensiswere normal,which is not the reason of species scarcity.
Magnoliawufengensis;megasporogenesis;microsporogenesis;female and male gametophyte development;cytological observation
时间:2016-08-0909:41DOI:10.13207/j.cnki.jnwafu.2016.09.024
2015-01-30
国家自然科学基金项目“红花玉兰及多瓣红花玉兰花器官特征决定B类基因表达模式和功能分析”(31170625);国家林业公益性行业科研专项“红花玉兰品种选育与规模化繁殖技术研究”(201504704);北京市科技计划项目“首都平原百万亩造林科技支撑工程”(Z121100008512002);国家林业局林业科学技术研究项目“红花玉兰种质资源收集保护、遗传测定与开发”(2006-39)
陈丽园(1985-),女,山西晋中人,博士,主要从事森林培育研究。E-mail:yuanyuan.0009@163.com
马履一(1957-),男,云南大理人,教授,博士,博士生导师,主要从事森林培育研究。E-mail:maluyi@bjfu.edu.cn
S685.15
A
1671-9387(2016)09-0181-05
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160809.0941.048.html
猜你喜欢
贵州林业科技(2022年2期)2022-06-22
西北植物学报(2022年4期)2022-05-24
科学大众·教师版(2022年6期)2022-05-23
北京航空航天大学学报(2021年9期)2021-11-02
锦绣·上旬刊(2019年12期)2019-09-10
新青年(2019年8期)2019-08-22
食品与健康(2018年8期)2018-08-23
创作评谭(2014年3期)2014-05-12
体育教学(2009年7期)2009-09-07
读书(1983年8期)1983-07-15
