
桑天牛越冬幼虫的空间分布与抽样技术
2016-10-31 03:26:09苏振国刘永辉刘建波朱红涛
浙江农业学报 2016年2期
苏振国,刘永辉,刘建波,朱红涛
(云南省农业科学院 蚕桑蜜蜂研究所,云南 蒙自 661101)
桑天牛越冬幼虫的空间分布与抽样技术
苏振国,刘永辉,刘建波,朱红涛
(云南省农业科学院 蚕桑蜜蜂研究所,云南 蒙自 661101)
为明确桑天牛(AprionagermariHope)越冬幼虫的空间分布,采用6种聚集度指标(m*/m,C,k,Iδ,I,CA)和2种回归模型(Taylor的幂法则和m*-m)测定其空间分布,并利用Blackith种群聚集均数λ解析该幼虫种群的聚集成因。结果表明,其呈均匀分布,建立了桑天牛越冬幼虫抽样数公式(N=t2(1.0097m-0.1971)/D2)和序贯抽样模型(T(n)=0.2n± 0.4405),该模型可为桑天牛的预测预报及防治提供理论依据。
桑天牛;越冬幼虫;空间分布;抽样技术
桑天牛(AprionagermariHope),别名褐天牛、铁泡虫等,属鞘翅目(Coleoptera),天牛科(Cerambycidae),分布极广泛,在河北、辽宁、江苏、浙江、山东、江西、安徽、云南、四川等省普遍发生。该虫寄主范围广,除为害桑树(MorusalbaL.)外,还可为害杨树、苹果、枇杷、柳等树种,以幼虫蛀食桑树枝干,造成桑树生长不良,严重时致全株枯死。桑天牛越冬幼虫扩散能力有限,生活在树干内经历2~3个冬季[1-2]。研究昆虫的种群空间分布在理论上有助于了解昆虫的生态学特性及种群所处环境的相互关系,在实践上是确定生物统计分析的资料代换方式和制定抽样技术方案的依据[3]。为此,我们于2014年12月至2015年1月对桑天牛越冬幼虫的空间分布进行了调查研究,以期为该害虫的预测预报和防治提供理论依据。
1 材料与方法
1.1试验地概况及取样方法
试验地位于云南省农业科学院蚕桑蜜蜂研究所试验桑园,调查的桑树是2~3年树龄的云桑2号,属于新种植桑园,园内海拔1 280 m,地势平坦,土壤为红色黏土,属亚热带季风气候类型,年平均气温18.6 ℃,无霜期337 d,降雨主要集中在6—9月份,冬、春季干旱少雨,年均日照时数2 234 h,最高气温35.2 ℃,最低气温2.0 ℃[4],园区周围种植有枇杷、石榴、葡萄等果树。调查共选取21块样地,每样地逐株调查桑树195株,由于天牛幼虫在枝干内取食,难以直接得到天牛幼虫数量。因此,根据天牛幼虫蛀孔行为特征,以有新鲜虫粪的虫孔数作为幼虫的数量指标[5]。
1.2空间分布型测定的分析方法
根据田间调查得到的数据,以样地为单位,计算出幼虫均值即虫口密度m(头/株)、方差(S2)。通过计算平均拥挤度m*、扩散系数C、David指标I、Cassie指标CA、Waters指标k、Lloyd聚块性指标m*/m、Morisita指标Iδ等参数指标测定该幼虫的空间分布型。其中C<1为均匀分布,C=1为随机分布,C>1为聚集分布;I<0为均匀分布,I=0为随机分布,I>0为聚集分布;CA<0为均匀分布,CA=0为随机分布,CA>0为聚集分布;k<0为均匀分布,k>0为聚集分布;m*/m<1为均匀分布,m*/m=1为随机分布,m*/m>1为聚集分布;Iδ<1为均匀分布,Iδ=1为随机分布,Iδ>1为聚集分布[6]。运用Iwao法m*=a+bm[7]和Taylor法S2=amb[8-9]对分布型及成因进行分析检验。
1.3聚集成因分析
利用Blackith提出的种群聚集均数λ=mr/2k[10],式中m为虫口密度均值,k为负二项分布指数,r是自由度为2k时0.5概率值对应的χ2。当λ<2时,其聚集是由某些环境因素所致,而不是由于昆虫本身的聚集习性活动的缘故;当λ≥2时,其聚集是由于上述2个因素中的任一个因素所引起。
1.4最适理论抽样数模型
运用Iwao提出m*-m直线回归式为基础的最适理论抽样数模型,即运用公式:
式中,N为最适理论抽样数;t为一定置信度下t分布值(取t=1);D为允许误差,一般取D=0.1,0.2,0.3;a,b为m*-m直线回归式中的常数;m为平均虫口密度。
1.5序贯抽样
根据Iwao提出的新序贯抽样理论,确定序贯抽样表。公式为:
式中n为抽样株数,m0为临界防治密度,a,b为m*-m直线回归式中的常数。
1.6数据处理
调查数据用Excel和DPS软件进行计算处理。
2 结果与分析
2.1空间分布型的测定
2.1.1聚集度指标法的测定结果
应用6种聚集度指标测定桑天牛越冬幼虫空间分布型结果见表1。其中扩散系数C<1,David指标I<0,Cassie指标CA<0,Waters指标k<0,Lloyd聚块性指标m*/m<1,Morisita指标Iδ<1,判定桑天牛越冬幼虫在桑树上的空间分布为均匀分布。
2.1.2Iwao线性回归法的测定
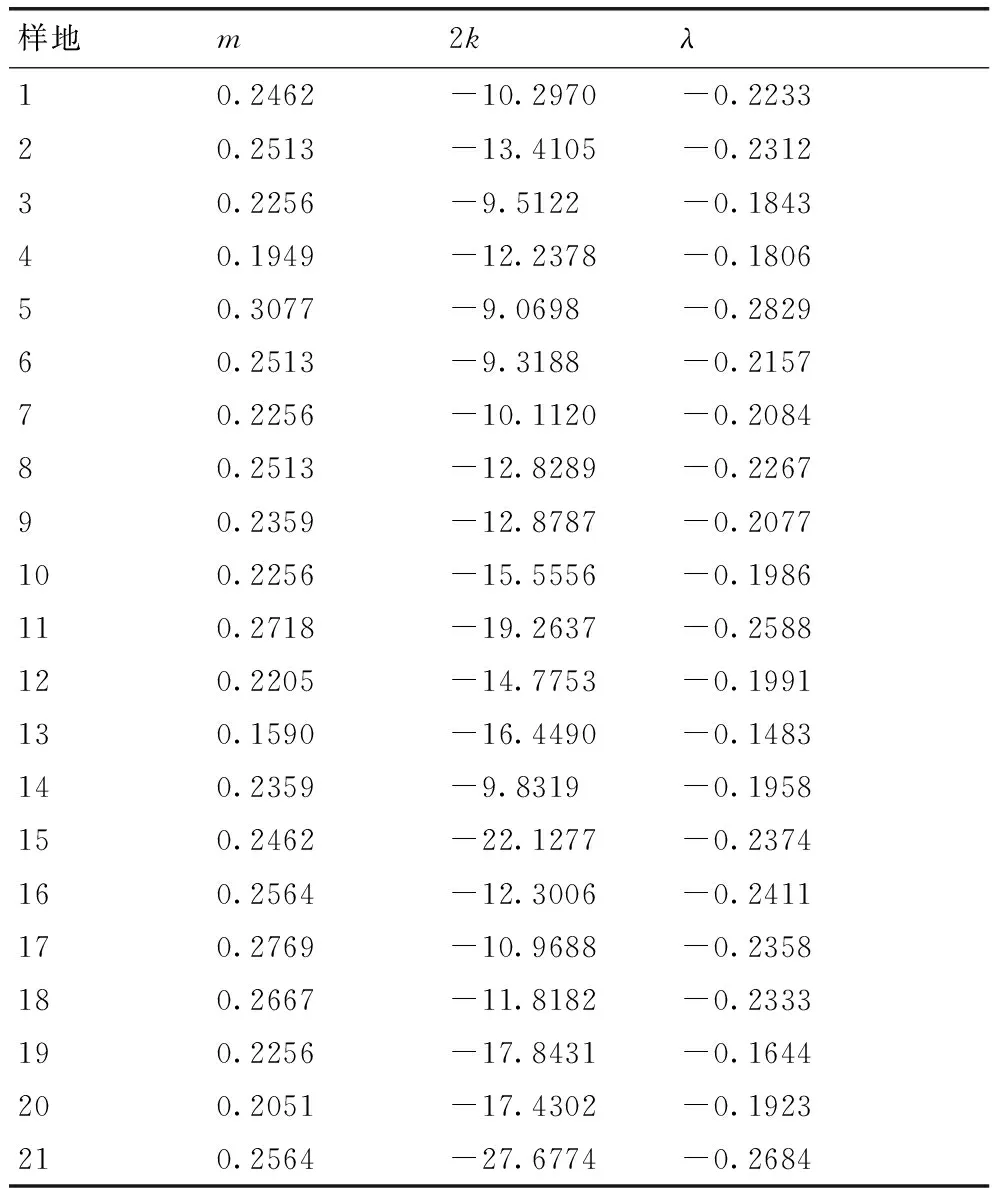
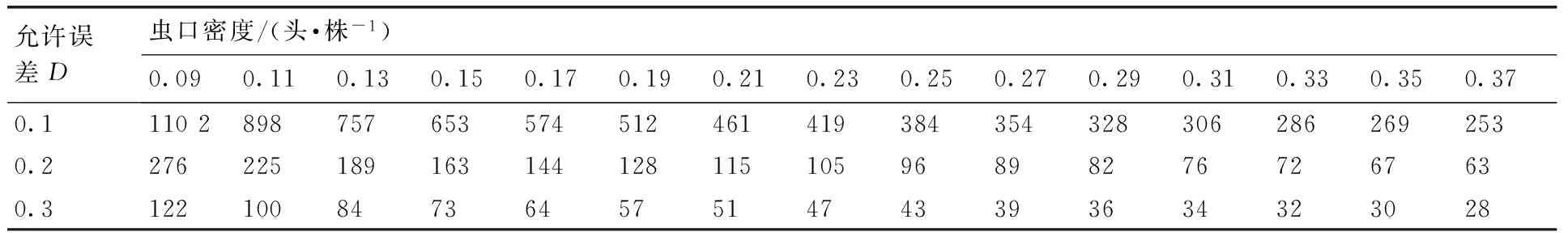
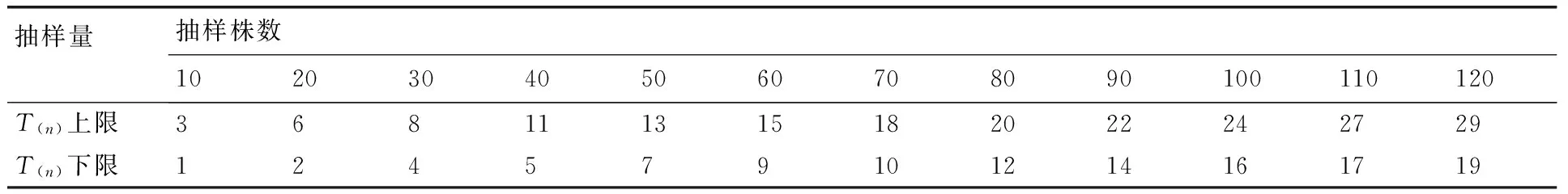
根据相关指标计算得到m*-m直线回归式为m*=0.0097+0.8029m,r=0.937 7,式中0 2.1.3Taylor的幂法则的测定 根据相关指标计算得到logS2=0.9533logm-0.0485,r=0.997 3,loga<0,b<1,说明该幼虫为均匀分布,种群密度越大分布越均匀。 2.2聚集因素分析 运用Blackith的种群聚集均数公式,计算不同样地的聚集均数,结果如表2所示。各样地的λ值均小于2,说明幼虫的均匀分布原因是某些环境因素所引起[11]。 2.3Iwao理论抽样数模型 将建立的m*-m回归模型中a,b的值带入Iwao最适理论抽样数模型,得出最适抽样数公式: 表1桑天牛越冬幼虫的聚集度指标 Table 1Aggregation indexes of overwintering larvae ofAprionagermariHope 样本采集地编号平均虫口密度m/(头·株-1)平均拥挤度m*方差S2扩散系数CWaters指标kCassie指标CALloyd聚块性指标m*/mMorisita指标IδDavid指标I10.24620.19830.23440.9522-5.1485-0.19420.80570.8229-0.047820.25130.21380.24190.9625-6.7053-0.14910.85080.8685-0.037530.21030.16600.20100.9558-4.7561-0.21030.78970.8094-0.044240.19490.16300.18870.9682-6.1189-0.16340.83650.8591-0.031850.30770.23980.28680.9321-4.5349-0.22050.77940.7927-0.067960.24100.18930.22860.9483-4.6594-0.21460.78530.8024-0.051770.22560.18100.21560.9554-5.0560-0.19780.80220.8208-0.044680.25640.21640.24620.9600-6.4145-0.15590.84410.8613-0.040090.23590.19930.22730.9634-6.4393-0.15530.84470.8634-0.0366100.21540.18770.20940.9723-7.7778-0.12860.87140.8926-0.0277110.27180.24360.26410.9718-9.6319-0.10380.89610.9134-0.0282120.22050.19070.21390.9702-7.3877-0.13540.86460.8852-0.0298130.15900.13960.15590.9807-8.2245-0.12160.87840.9076-0.0193140.23080.18380.21990.9531-4.9160-0.20340.79650.8146-0.0469150.24620.22390.24070.9778-11.0638-0.09040.90960.9289-0.0222160.26150.21900.25040.9575-6.1503-0.16260.83740.8541-0.0425170.27690.22640.26290.9495-5.4844-0.18230.81760.8330-0.0505180.26670.22150.25460.9549-5.9091-0.16920.83070.8470-0.0451190.17950.15940.17590.9799-8.9216-0.11210.88790.9140-0.0201200.20510.18160.20030.9765-8.7151-0.11470.88520.9079-0.0235210.28210.26170.27630.9796-13.8387-0.07230.92770.9449-0.0204 表2桑天牛越冬幼虫种群密度与聚集均数分析 Table 2Analysis of average aggression size of overwintering larvae ofAprionagermariHope 样地m2kλ10.2462-10.2970-0.223320.2513-13.4105-0.231230.2256-9.5122-0.184340.1949-12.2378-0.180650.3077-9.0698-0.282960.2513-9.3188-0.215770.2256-10.1120-0.208480.2513-12.8289-0.226790.2359-12.8787-0.2077100.2256-15.5556-0.1986110.2718-19.2637-0.2588120.2205-14.7753-0.1991130.1590-16.4490-0.1483140.2359-9.8319-0.1958150.2462-22.1277-0.2374160.2564-12.3006-0.2411170.2769-10.9688-0.2358180.2667-11.8182-0.2333190.2256-17.8431-0.1644200.2051-17.4302-0.1923210.2564-27.6774-0.2684 N=t2/D2(1.009 7m-0.197 1),取t=1,允许误差D为0.1,0.2,0.3时,虫口密度为0.09,0.11,0.13,0.15,0.17,0.19,0.21,0.23,0.25,0.27,0.29,0.31,0.33,0.35,0.37头·株-1时,求出桑天牛越冬幼虫最适理论抽样数量结果见表3。 2.4序贯抽样模型 拟设桑天牛幼虫防治指标m0=0.2头·株-1,t=1,a=0.009 7,b=0.802 9,代入可得序贯抽样模型:T(n)=0.2n±0.4405,令n=10,20,30,40,……200,可得出序贯抽样表见表4。 本文通过对桑天牛越冬幼虫的6种聚集度指标法、Iwao和Taylor回归方程、聚集均数的判定表明,桑天牛越冬幼虫空间分布型呈均匀分布,此种分布是由幼虫个体间相互排斥和环境因素引起。而黄大庄等[12]研究认为,桑天牛幼虫在杨树上为聚集分布,且具有密度依赖性。刘康成[13]则认为,桑天牛幼虫在桑树上的空间分布型为聚集分布,且幼虫个体间相互吸引,可见,桑天牛幼虫在不同寄主或同种寄主的不同生长环境中其空间分布会有所不同。具体分析形成均匀分布的原因可能是,桑天牛产卵更喜欢选择在树体高大粗壮的寄主上产卵[14],本次调查的桑树树龄2~3年,树形、树势尚处于生长阶段还未完全定型,会降低桑天牛产卵量和引起桑天牛幼虫间的食物竞争[15];调查所采用的方法是根据新鲜排粪孔作为幼虫虫口数的判定指标,桑天牛的幼虫活动如排粪随季节、光照、温度的变化,冬季气温低于10 ℃排粪也几乎停止,可见环境因素当中的温度因子也会带来一定影响[16],可以解释此种均匀分布是由幼虫个体间相互排斥和环境因素相互作用引起。 文中建立了最适抽样数公式:N=t2/D2(1.0097m-0.1971)和序贯抽样模型:T(n)=0.2n±0.4405。在虫口密度低的情况下,抽样量较多,虫口密度高的情况下,抽样量较少。在相同虫口密度下,允许误差的大小也决定抽样量的多少。在田间应用序贯抽样调查时,若调查样本的累计虫量超过上限,即判为防治对象田;低于下限则认为暂时不需要防治;当累计虫量在上下限之间,则应继续进行调查,直到最大抽样数。 表3桑天牛越冬幼虫理论抽样数 Table 3Theoretical sampling number of overwintering larvae ofAprionagermariHope 允许误差D虫口密度/(头·株-1)0.090.110.130.150.170.190.210.230.250.270.290.310.330.350.370.111028987576535745124614193843543283062862692530.2276225189163144128115105968982767267630.312210084736457514743393634323028 表4桑天牛越冬幼虫序贯抽样表 Table 4Sequential sampling table of overwintering larvae ofAprionagermariHope 抽样量抽样株数102030405060708090100110120T(n)上限368111315182022242729T(n)下限124579101214161719 [1]郑声镛, 高祖训, 徐国宇,等. 桑树病虫害防治学[M].北京:农业出版社,1990:310-312. [2]黄大庄. 桑天牛的研究进展[J]. 河北林学院学报, 1996, 11(3-4): 290-295. [3]于新文, 况荣平.咖啡天牛幼虫种群的空间分布型及应用[J]. 动物学研究, 1997, 18(1): 39-44. [4]杜伟, 储一宁, 李腾芳, 等. 云南省桑园内茶黄螨的发生规律与防治方法[J]. 蚕业科学,2013, 39(6):1072-1073. [5]王希蒙, 李崇阳, 张真. 黄斑星天牛空间分布型的初步研究[J].林业科学, 1988, 24(3): 352-357. [6]张文军. 生态学研究方法[M]. 广州: 中山大学出版社, 2007: 45-55. [7]IWAO S. A new regression method for analyzing the aggregation pattern of animal populations[J].ResearchesonPopulationEcology, 1968, 10(1): 1-20. [8]TAYLOR L R. Aggregation, variance and the mean[J].Nature, 1961, 189(4776): 732-735. [9]TAYLOR L R. Assessing and interpreting the spatial distributions of insect populations[J].AnnualReviewofEntomology,1984,29(1):321-357. [10]丁岩钦. 昆虫数学生态学[M]. 北京:科学出版社,1980:1-36, 84-124. [11]赵阳, 朱景乐, 李芳东, 等. 杜仲梦妮夜蛾幼虫的空间分布型及抽样技术[J]. 环境昆虫学报, 2014, 36(4): 629-634. [12]黄大庄, 岳书奎, 闫浚杰, 等. 北方桑天牛水平与垂直双向分布规律的研究[J]. 东北林业大学学报,1993, 21(2): 36-39. [13]刘康成. 桑天牛幼虫空间分布型及抽样技术[J]. 昆虫知识,1997, 34(2): 80-81. [14]王志刚, 黄大庄, 阎浚杰. 桑天牛在不同树种或品种间产卵选择性试验[J]. 河北林学院学报,1994, 9(1): 65-68. [15]董廷宣, 彭炳香. 桑天牛的产卵规律的初步探讨与防治[J]. 中国蚕业,2003, 24(4): 30-31. [16]黄大庄, 王志刚, 阎浚杰. 桑天牛幼虫活动规律[J]. 东北林业大学学报,1996, 25(6): 83-86. (责任编辑张韵) Spatial distribution pattern and sampling technique of overwintering larvae of Apriona germari Hope SU Zhen-guo, LIU Yong-hui, LIU Jian-bo, ZHU Hong-tao (InstituteofSericultureandApiculture,YunnanAcademyofAgriculturalSciences,Mengzi661101,China) The spatial distribution patterns of overwintering larvae ofAprionagermarilarva inMorusalbawere studied using six spatial distribution pattern analysis methods(m*/m,C,k,Iδ,I,CA)and two regression equations(Taylor power and m*-m), the assembling reasons were analyzed with average aggression size λ. The results indicated that spatial distribution patterns ofAprionagermarilarva population was in uniform distribution. The optimal sampling equation(N=t2(1.0097m-0.1971)/D2) and sequential sampling equation(T(n)=0.2n±0.4405) were established. This paper may provide the basis for prediction and control ofAprionagermari. AprionagermariHope; overwintering larvae; spatial distribution pattern; sampling techniques 10.3969/j.issn.1004-1524.2016.02.20 2015-07-11 现代农业产业技术体系建设专项资金“蚕桑”(CARS-22-SYZ27) 苏振国(1983—),男,山西右玉人,硕士,助理研究员,从事桑树病虫害防治工作。E-mail:szgmcn@163.com Q968.1 A 1004-1524(2016)02-0302-04 苏振国,刘永辉,刘建波,等. 桑天牛越冬幼虫的空间分布与抽样技术[J].浙江农业学报,2016,28(2): 302-305.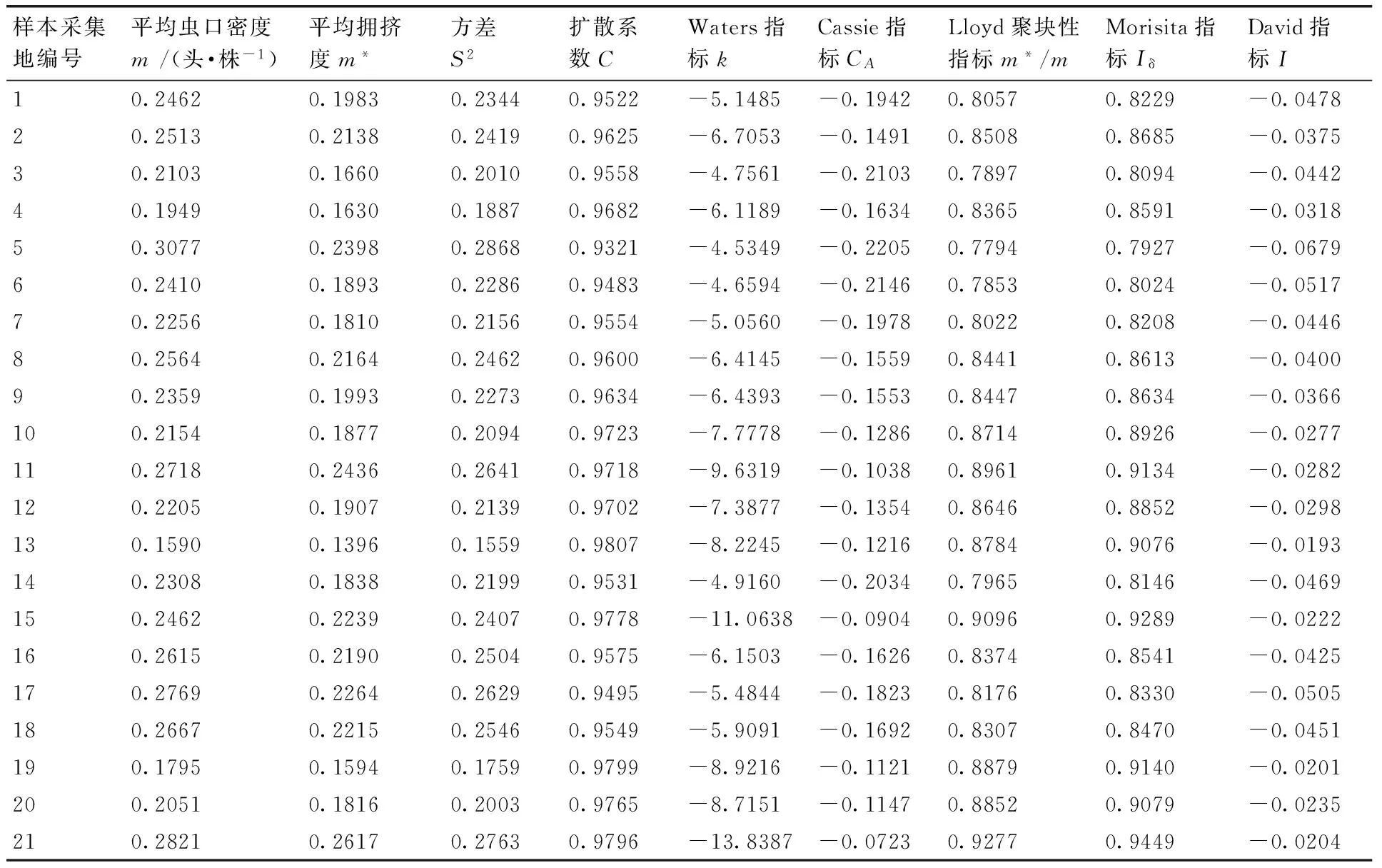
3 讨论
猜你喜欢
音乐教育与创作(2022年1期)2022-04-26 02:21:20
今日农业(2021年3期)2021-12-05 01:46:23
今日农业(2021年9期)2021-07-28 07:08:22
今日农业(2021年4期)2021-06-09 06:59:44
天津农林科技(2021年2期)2021-04-16 02:54:24
今日农业(2020年16期)2020-12-14 15:04:59
工程与建设(2019年5期)2020-01-19 06:22:26
小学生作文(中高年级适用)(2018年3期)2018-04-18 01:24:40
中学生数理化·高二版(2017年3期)2017-07-07 08:47:25
学生天地(2016年16期)2016-05-17 05:46:06
