
体胚诱变结缕草属突变体形态特征及生长特性评价
2016-10-31 03:26:05方敏彦孔维亮
浙江农业学报 2016年2期
方敏彦,章 明,孔维亮
(1.江苏农林职业技术学院,江苏 句容 212400 ;2.句容市园林管理中心,江苏 句容 212400)
体胚诱变结缕草属突变体形态特征及生长特性评价
方敏彦1,章明2,孔维亮1
(1.江苏农林职业技术学院,江苏 句容 212400 ;2.句容市园林管理中心,江苏 句容 212400)
对结缕草属3个种的119份体胚诱变突变体的13个外部形态特征及生长特性进行了相关分析。结果表明:突变体的所有13个性状的变异均较大,其中匍匐枝总长度、单株覆盖面积、叶毛及密度的变异系数均高于1。主成分分析结果表明,单株覆盖面积、匍匐枝总长度、密度以及直立枝节间长4个性状对结缕草属突变体的遗传多样性研究具有重要意义。在聚类分析基础上,将119份突变体种源划分为4组,分别为细叶多毛型突变体、粗径早绿型突变体、短叶少毛型突变体、宽叶晚绿型突变体。
结缕草属;体胚诱变突变体;形态特征;主成分分析;聚类分析
结缕草属(ZoysiaWilld)植物为禾本科画眉草亚科的多年生低矮草本植物,是一种优良的暖季型草坪草。由于其具有广泛的土壤适应性、较强的抗性(抗旱性、耐盐性、抗病虫害等)、耐粗放管理和寿命长等特性,是优良的草坪建植材料[1]。该属植物虽具有许多优良特性,但由于其野生资源被无节制地原始性采挖,破坏严重,且繁殖速度慢,可直接利用的种植材料极少,目前在我国的应用很有限。此外,结缕草属培育的新品种很少,已有品种存在低温适应范围窄等问题。
近年来,体细胞无性系变异开始应用在植物种质创新上。其中,体细胞胚(体胚)变异具有特殊的科学价值。植物体胚的发生与合子胚的发育相似,是由植物某些组织的单个体细胞形成胚胎创造出新个体的过程(称为无性胚胎发生/体胚形成)[2]。体胚发生是细胞全能性表达最为完全的一种方式,它不仅说明植物体细胞具有完整的全套遗传信息,而且重演了合子胚形态发生的进程[3]。体胚诱变育种,变异频率高,其中单基因变异可改变植物的个别性状,而保持其他优良性状不变[4]。因此,通过体胚诱变开展结缕草属植物种质创新,能在尽可能保存原有优良性状的同时创造出具目标性状的新种质,且该属植物均以无性繁殖为主,体胚诱变获得同质突变体后,在M1代即可对整株进行优良性状选择,大大优化了育种程序,具有重要的科学意义。
试验选用结缕草属中的中华结缕草(ZoysiasinicaHance)、沟叶结缕草[Zoysiamatrella(L.)Merr.]、兰引Ⅲ号结缕草(ZoysiajaponicaSteud. cv. Lanyin No.3),通过体胚不同发育时期,采用60Co-γ辐射诱变及低温处理等复合处理技术,得到了性状稳定的同质突变体600余份,通过初选对其中119份突变体进行了形态特征及生长特性评价,以期为突变群体的遗传变异多样性研究及优良突变种质的筛选奠定基础。
1材料与方法
1.1供试材料
供试材料为中华结缕草、兰引Ⅲ号结缕草、细叶结缕草诱变的119份突变体(A1-F29依次编号),突变体是项目组通过60Co-γ辐射诱变、及低温处理这3个品种的体细胞胚后获得的。于2009年栽植于江苏农林职业技术学院草坪实验地内,常规养护管理,2015年5—7月进行田间观测及样品采集。
1.2观测项目和方法
直立枝叶片长、宽:直立枝顶部向基部第4片成熟叶[5],用0.5 mm刻度直尺测定,重复10次,用cm表示。
直立枝节间长、直径:直立枝顶端向基部第4节茎或直立枝中部节间,用0.5 mm刻度直尺测定,重复10次,用cm表示。
直立枝株高:地表至直立枝自然顶部高度,将一张A4纸轻轻放在草坪上,用直尺从地面量至纸底[6],用cm表示。
匍匐枝叶片长、宽:取匍匐枝顶部向基部第4片成熟叶,重复5次,用cm表示。
例如给出带电粒子的质量为m、带电量为q以及初速度v0,使粒子从两极板中部穿过匀强电场,并给出极板长度L,极板间距离为d,极板间的电压U(如图一),那么粒子将会有两种运动结果,其一是带电粒子可能落在平行板电容器的下极板上,其二是带电粒子可能逸出平行板电容器。
匍匐枝节间长:匍匐枝顶端向基部第4节茎,重复5次,用cm表示。
匍匐枝总长度:测定匍匐枝在不同方向上的总长度[7],用cm表示。
密度:具体指草层中直立枝的密度。观测时采用9级制打分法[8],其中1为密度最低,6为可以接受,9为密度最低,3人同时打分取平均值。
叶毛(表面/背面):分五级制[9]。其中0为无毛,1为叶毛很稀疏,2为叶毛较稀疏,3为叶毛较密,4为叶毛很密,3人同时打分取平均值。
叶色:利用日本产SPAD-502叶绿素仪在距离叶片基部50%左右处测定[10-11],以叶片叶绿素的相对含量表示。
单株覆盖面积:按椭圆计算[12],以最大扩展距离为长轴a、与最大扩展距离方向垂直的距离为短轴b,S=πab,用(cm2·株-1)表示。
1.3数据与统计
利用Excel对不同种类结缕草形态学上的数量性状指标测定值进行初步统计分析,计算平均数、标准差、变异系数,进而比较各个种的种内遗传变异程度。利用SPSS 17.0软件对不同种类结缕草的形态特征和生长习性进行指标间相关性分析、主成分分析及聚类分析。
2 结果与分析
2.1突变体形态特征、生长特性变异及相关性分析
2.1.1突变体形态特征、生长特性变异
在13个外部性状中,匍匐枝总长度、单株覆盖面积、叶毛及密度的变异系数均高于1,其中匍匐枝总长度为7.2~30.6 cm,变异系数为1.06,单株覆盖面积为193.4~2 859.9 cm2·株-1,变异系数为1.01,叶毛的变异系数为1.58,密度的变异系数为1.02。其次是叶色、直立枝直径、匍匐枝节间长,其变异系数都大于0.25。变异系数是揭示样本差异的参数,变异系数越大,则样本间的差异越大,一般认为变异系数大于10%就说明样本间的差异较大[13]。本研究中所有性状的变异系数都大于10%,说明突变体种质资源外部性状种内变异很大,遗传多样性非常丰富(表1)。
表1突变体形态特征、生长特性变异
Table 1Morphological and growth characteristics ofZoysiamutants
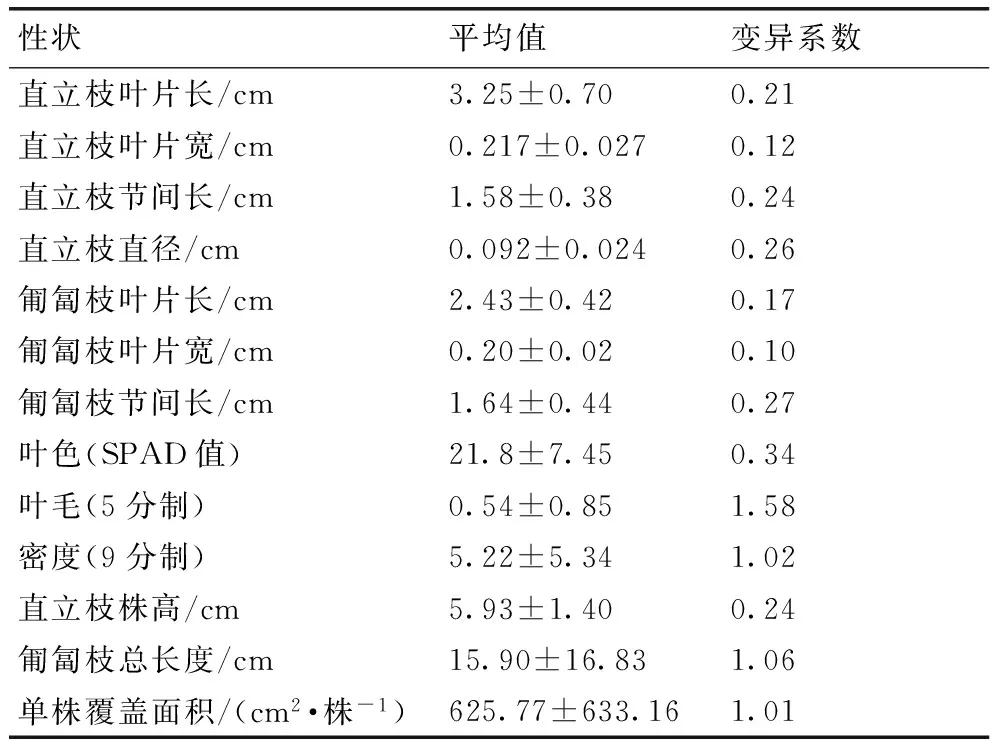
性状平均值变异系数直立枝叶片长/cm3.25±0.700.21直立枝叶片宽/cm0.217±0.0270.12直立枝节间长/cm1.58±0.380.24直立枝直径/cm0.092±0.0240.26匍匐枝叶片长/cm2.43±0.420.17匍匐枝叶片宽/cm0.20±0.020.10匍匐枝节间长/cm1.64±0.440.27叶色(SPAD值)21.8±7.450.34叶毛(5分制)0.54±0.851.58密度(9分制)5.22±5.341.02直立枝株高/cm5.93±1.400.24匍匐枝总长度/cm15.90±16.831.06单株覆盖面积/(cm2·株-1)625.77±633.161.01
2.1.2突变体形态特征、生长特性相关性分析
由表2可以看出,突变体外部性状之间不是相互独立的,而是存在一定的相关关系。结缕草属突变体植物的直立枝叶片长与直立枝直径之间呈显著的正相关,相关系数为0.250;直立枝叶片宽与直立枝叶片长、直立枝直径之间都呈显著的正相关,相关系数分别是0.233和0.187;直立枝节间长与直立枝直径呈显著的负相关,相关系数为-0.207;因此,结缕草突变体种质的很多外部性状在遗传上是相互连锁的,在选育新品种时,可以同时获得几个性状都优良的新品种。
2.2突变体表型性状的主成分分析
从主成分分析结果(表3)中可以看出,对于结缕草突变体而言,在第一主成分中,单株覆盖面积、匍匐枝总长度是比较稳定的性状,这些性状解释了结缕草属的大部分变异,也成为结缕草属分组的主要依据。在第二主成分中,密度、直立枝节间长是比较重要的指标。直立枝叶片宽、叶色、叶毛归属第三主成分,匍匐枝叶片长、匍匐枝叶片宽归属第四主成分,而直立植株高、直立枝叶片长、匍匐枝节间长、直立枝直径归属第五主成分。单株覆盖面积、匍匐枝总长度、密度以及直立枝节间长这4个性状可作为研究结缕草属突变体种内差异的主要依据。
表2突变体形态特征、生长特性的相关系数
Table 2Correlation coefficient of morphological and growth characteristics ofZoysiamutants
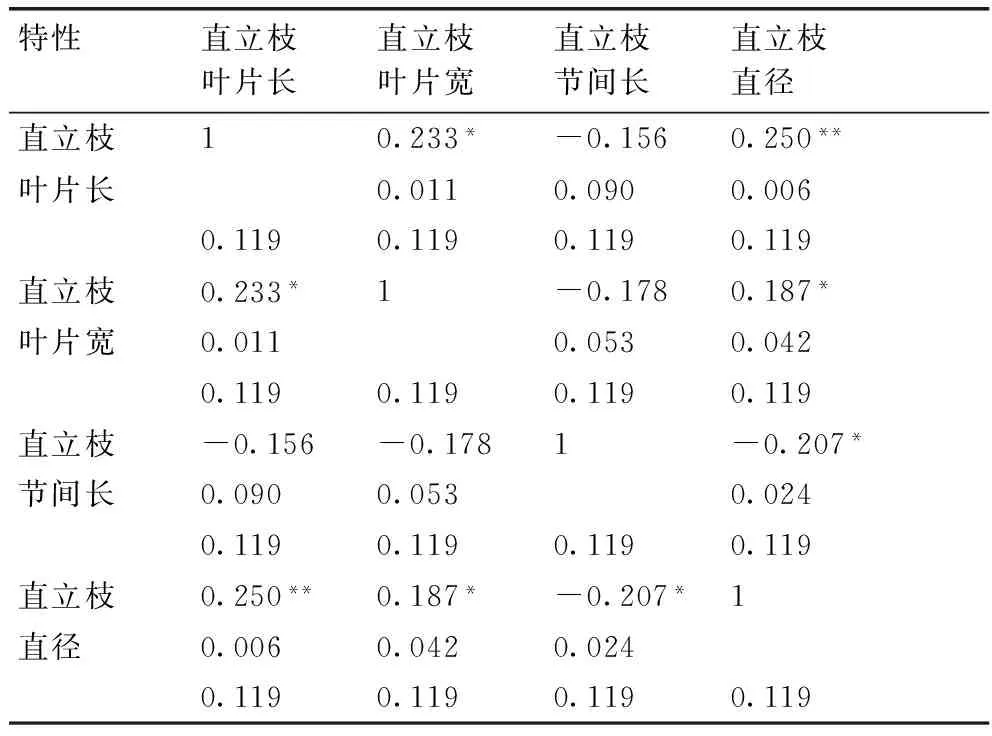
特性直立枝叶片长直立枝叶片宽直立枝节间长直立枝直径直立枝10.233*-0.1560.250**叶片长0.0110.0900.0060.1190.1190.1190.119直立枝0.233*1-0.1780.187*叶片宽0.0110.0530.0420.1190.1190.1190.119直立枝-0.156-0.1781-0.207*节间长0.0900.0530.0240.1190.1190.1190.119直立枝0.250**0.187*-0.207*1直径0.0060.0420.0240.1190.1190.1190.119
注:*和**分别表示相关系数达到显著和极显著水平。
表3主成分分析结果
Table 3The results of principal component analysis
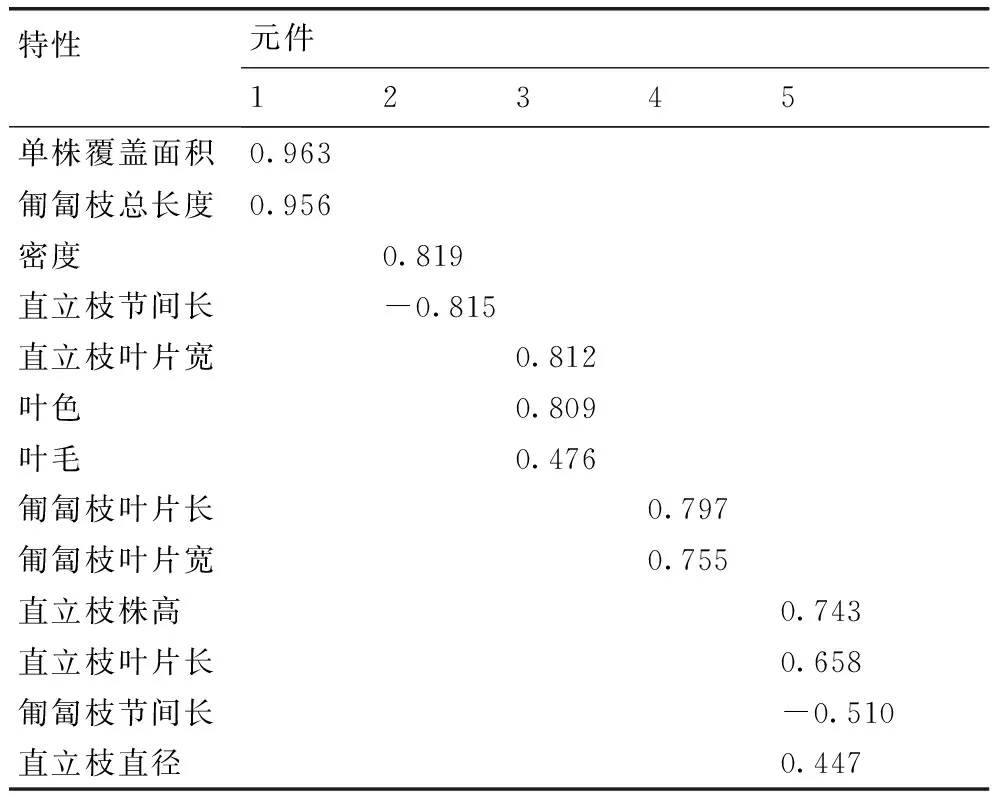
特性元件12345单株覆盖面积0.963匍匐枝总长度0.956密度0.819直立枝节间长-0.815直立枝叶片宽0.812叶色0.809叶毛0.476匍匐枝叶片长0.797匍匐枝叶片宽0.755直立枝株高0.743直立枝叶片长0.658匍匐枝节间长-0.510直立枝直径0.447
2.3突变体聚类分析
根据13个性状的聚类结果,在欧式平方距离18处可以将119份突变体种质材料分成4组(图1)。由表4可知,Ⅰ组有4份种源(A15,C30,E22,F7),主要特点表现在叶片长(3.5~5.3 cm)而窄(0.15~0.22 cm),直立枝直径细(0.05~0.1 cm),叶绿素含量低(11.4~20.3),匍匐枝节间短(0.6~1.8 cm),直立植株矮(4.1~6.5 cm),密度低而叶毛密集。可应用于公共绿地、公园和小区绿化。Ⅱ组有3份种源(A11,B4,E17),主要特点是直立枝直径粗(0.1~0.12 cm),叶绿素含量高(15.7~26.8 cm),匍匐枝长(11.6~ 15.1 cm),密度高且返青早。可进行营养繁殖,应用于运动场及水土保持。Ⅲ组有9份种源(A18,B12,B27,C2,C6,D4,E5,E15,F3),主要特点表现为直立枝叶片短(2.4~4.1 cm),叶表面无毛或稀疏,匍匐枝和直立枝的节间长(1.2~2.5 cm),但返青比较迟。Ⅳ组有103份种源,主要特点则表现为匍匐枝短(7.2~17.8 cm),直立枝和匍匐枝的叶片都比较宽(0.21~0.29 cm)。
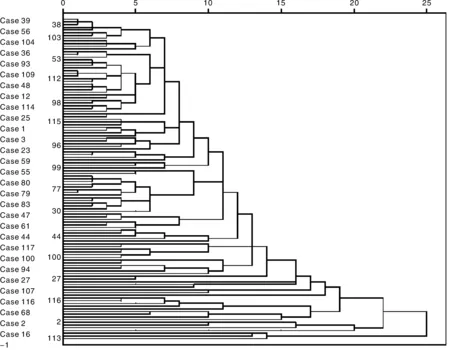
图1 突变体形态聚类图Fig.1 Cluster figure of Zoysia mutants
表4四个突变体组的形态特征和生长特性
Table 4Morphological and growth characteristics of the four groups ofZoysiamutants
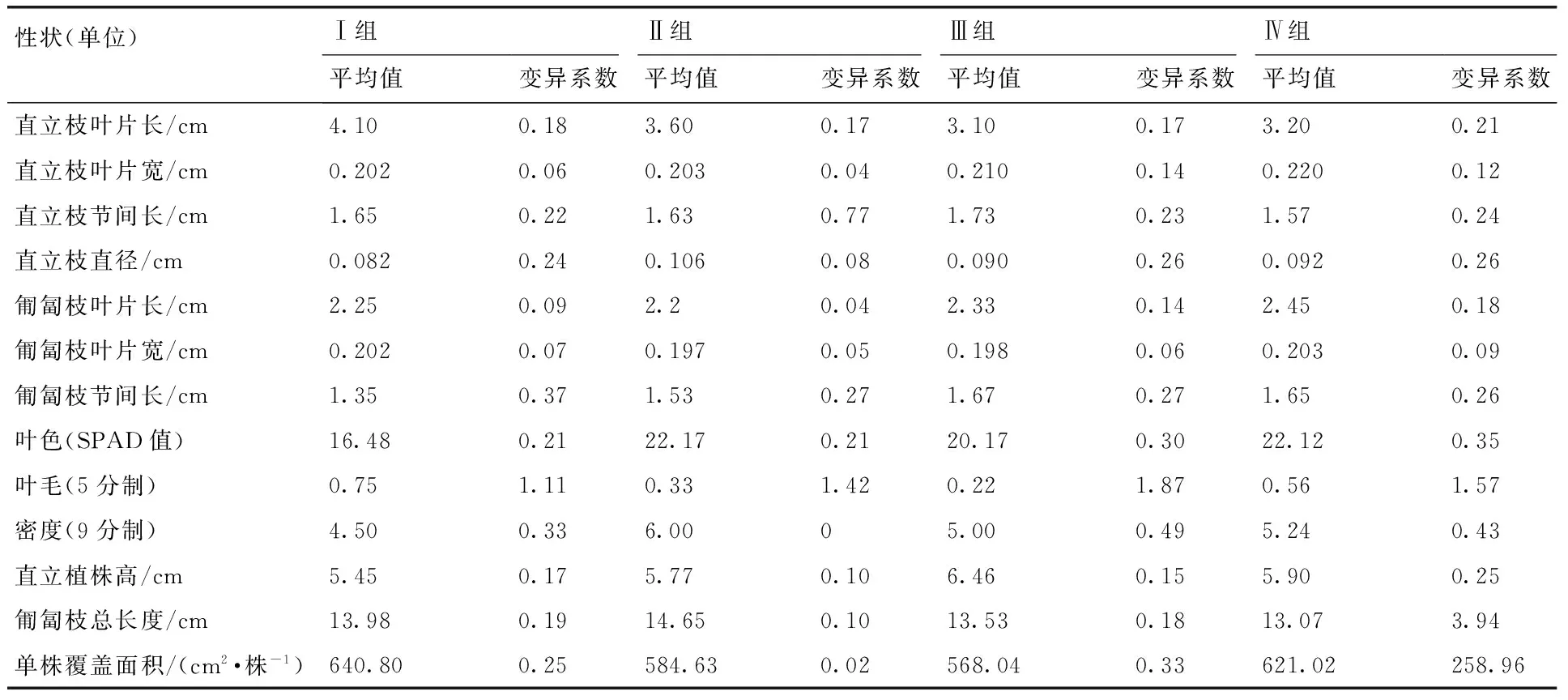
性状(单位)Ⅰ组Ⅱ组Ⅲ组Ⅳ组平均值变异系数平均值变异系数平均值变异系数平均值变异系数直立枝叶片长/cm4.100.183.600.173.100.173.200.21直立枝叶片宽/cm0.2020.060.2030.040.2100.140.2200.12直立枝节间长/cm1.650.221.630.771.730.231.570.24直立枝直径/cm0.0820.240.1060.080.0900.260.0920.26匍匐枝叶片长/cm2.250.092.20.042.330.142.450.18匍匐枝叶片宽/cm0.2020.070.1970.050.1980.060.2030.09匍匐枝节间长/cm1.350.371.530.271.670.271.650.26叶色(SPAD值)16.480.2122.170.2120.170.3022.120.35叶毛(5分制)0.751.110.331.420.221.870.561.57密度(9分制)4.500.336.0005.000.495.240.43直立植株高/cm5.450.175.770.106.460.155.900.25匍匐枝总长度/cm13.980.1914.650.1013.530.1813.073.94单株覆盖面积/(cm2·株-1)640.800.25584.630.02568.040.33621.02258.96
3 结论与讨论
3.1结缕草属突变体表型变异特征
由于结缕草属突变体的诱变手段不同,导致在外部形态上存在很大的差异,本研究在对119份突变体的13个外部性状观测分析后发现,突变体的外部形态特征变异系数都大于10%,即遗传多样性非常丰富。在13个外部性状中,平均变异系数(CV)由大到小依次为叶毛、匍匐枝总长度、密度、单株覆盖面积、叶色、匍匐枝节间长、直立枝直径、直立枝株高、直立枝节间长、直立枝叶片长、匍匐枝叶片长、直立枝叶片宽、匍匐枝叶片宽。结缕草属突变体的部分外部性状之间存在一定的相关性,直立枝叶片长与直立枝叶片宽、直立枝直径都呈显著的相关性。本结论与Diesburg[14]的研究结果一致,即高度性状(包括生殖枝高度和草层高度)、叶片长度和叶片宽度相互间呈正相关。
3.2基于表型性状的主成分分析及聚类分析
通过主成分分析,发现单株覆盖面积、匍匐枝总长度、密度以及直立枝节间长这4个性状可作为研究结缕草属体胚诱变突变体遗传多样性的主要依据。宣继萍[8]指出,结缕草属植物叶片宽度、匍匐茎长度、直径、每穗小穂数、小穗柄长度、小穗长度、生殖枝高度、叶舌的颜色及有无毛9个性状是结缕草属形态分类的重要依据,与本文结论有一些差异,但在匍匐茎长度等方面是一致的,这可能是结缕草属体胚诱变突变体变异后的群体特性。
基于聚类分析的结果,可以将结缕草突变体划分为4组,Ⅰ组细叶多毛型结缕草,Ⅱ组粗径早绿型结缕草,Ⅲ组短叶少毛型结缕草,Ⅳ组宽叶晚绿型结缕草。本研究结果说明,获得的结缕草体胚诱变突变群体具有丰富的遗传多样性,可为草坪地被体胚诱变育种提供理论依据。同时,证明体胚诱变可拓宽结缕草属植物的遗传基础、实现优良种质创新、选择优异基因,并为大规模结缕草突变体库的构建及功能基因组学研究奠定了基础。
[1]贺小霞.狗牙根和海滨雀稗的形态特征与 AFLP遗传多样性研究[D].上海:上海交通大学, 2011.
[2]陶雷.刺五加体细胞胚发生机制的初步研究[D].哈尔滨:东北林业大学,2013.
[3]ANTONY C, IGNACIMUTHU S. Efficient somatic embryogenesis and plant regeneration from shoot apex explants of different Indian genotypes of finger millet (Eleusinecoracana(L.)Gaertn.)[J].InVitroCellular&DevelopmentalBiology-Plant, 2008,44(5):427-435.
[4]王婷婷,徐国荣,张举仁,等.细胞工程技术选育的草地早熟禾新种质的耐热性分析[J].草业学报,2009,18(2):60-65.
[5]李亚,凌萍萍,刘建秀,等.中国结缕草属植物(Zoysiaspp.)地下部分分布和形态类型的多样性[J].植物资源与环境学报,2002,11(2):39-44.
[6]胡利珍.野生结缕草品系生物学特性及坪用现状的研究[D].长沙:湖南农业大学,2007.
[7]杜永吉,金芳,孙吉雄,等.兰引Ⅲ号结缕草分株繁殖建坪的研究[J].草原与草坪,2008(1):54-58.
[8]宣继萍.结缕草属(ZosiaWilld.)植物种质资源多样性研究[D].南京:南京农业大学,2008.
[9]贾蕴琪.结缕草绿期与抗寒性研究[D].兰州:甘肃农业大学,2009.
[10]鱼欢,祖超,杨建峰,等.应用SPAD叶绿素仪测定不同位置胡椒叶片的SPAD值[J].热带作物学报,2012,33(10):1890-1895.
[11]薛香,吴玉娥.小麦叶片叶绿素含量测定及其与SPAD值的关系[J].湖北农业科学,2010,49(11):2701-2702.
[12]董沁.基于形态特征和AFLP的野生早熟禾遗传多样性研究[D].兰州:甘肃农业大学,2013.
[13]郭海林,陈宣,薛丹丹,等.结缕草属植物青绿期的遗传分析[J].草业学报,2009,18(4):147-153.
[14]DIESBURG K L. Expanded germplasm collections set stage for increased zoysiagrass breeding for turf use[J].Diversity,2000,16(1/2):49-50.
(责任编辑张韵)
Evaluation of morphological and growth characteristics of somatic embryo mutants of Zoysia
FANG Min-yan1,ZHANG Ming2,KONG Wei-liang1
(1.JiangsuPolytechnicCollegeAgricultureandForestry,Jurong212400,China; 2.JiangsuJurongGardenManagementCenter,Jurong212400,China)
Thirteen morphological and growth characteristics of somatic embryo mutants ofZoysiaincluding 3 species and 119 mutants were analyzed. Results showed that the variation of all the 13 traits were large, among which the variation coefficient of total length of creeping branch, covering area per plant, lamellar hair and density were higher than 1. The results of principal component analysis showed that single cover area, total length of creeping branch, density and erect internode length were meaningful for the study on genetic diversity ofZoysiamutants. Based on the clustering analysis, 119Zoysiamutants were divided into four groups, including narrow leaf and hairy type mutants, large diameter and early green mutants, short leaf and little hair type mutants, and wide leaf and lately green mutants.
Zoysia; somatic embryo mutants; morphological characteristics; principal component analysis; cluster analysis
10.3969/j.issn.1004-1524.2016.02.15
2015-10-30
奥林高尔夫教育与研究基金(AL2014004);江苏农林职业技术学院草坪创新团队
方敏彦(1980—),女,浙江永康人,副研究员,博士,主要从事园林植物栽培和育种研究。E-mail: fangyanhome@163.com
S812
A
1004-1524(2016)02-0269-05
方敏彦,章明,孔维亮. 体胚诱变结缕草属突变体形态特征及生长特性评价[J].浙江农业学报,2016,28(2): 269-273.
猜你喜欢
西北植物学报(2023年6期)2023-08-14 09:24:12
儿童故事画报·自然探秘(2022年12期)2022-11-24 07:29:15
工矿自动化(2022年7期)2022-08-13 08:01:04
吉林林业科技(2021年2期)2021-05-12 13:20:22
热带农业科学(2020年2期)2020-05-22 12:48:32
种子(2017年11期)2018-01-24 10:51:40
植物研究(2017年3期)2017-11-10 10:44:12
天津职业技术师范大学学报(2016年1期)2016-04-27 02:35:36
陕西林业科技(2016年5期)2016-04-05 22:57:54
哈尔滨商业大学学报(自然科学版)(2014年6期)2014-09-14 04:49:30
