
小麦β-酮脂酰CoA合成酶基因KCS的克隆与酵母表达
2016-10-27 01:03夏凌峰杨昊虹李春莲王中华
麦类作物学报 2016年9期
夏凌峰,史 雪,杨昊虹,李春莲,王中华,权 力
(西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌 712100)
小麦β-酮脂酰CoA合成酶基因KCS的克隆与酵母表达
夏凌峰,史 雪,杨昊虹,李春莲,王中华,权 力
(西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌 712100)
超长链脂肪酸是生物体内众多重要物质的合成底物。KCS基因编码β-酮脂酰CoA合成酶,该酶具有底物特异性,参与超长链脂肪酸延伸的缩合反应,是超长链脂肪酸合成的限速步骤。为了探究小麦KCS基因在超长链脂肪酸合成中的功能,采用同源克隆的方法从小麦(TriticumaestivumL.) 中克隆出KCS基因后,利用生物信息学对其编码序列进行分析,并在酿酒酵母(Saccharomycescerevisiae)中对其进行真核表达。结果表明,小麦 TaKCS6基因的开放阅读框为1 287 bp,编码428个氨基酸残基。结构域预测结果显示,TaKCS6蛋白含有III型聚酮合酶脂肪酸延伸酶和C末端3-酰基ACP合酶III结构域,属于KCSs蛋白家族。序列比对分析结果显示,TaKCS6氨基酸序列与拟南芥及其他植物的KCS6氨基酸序列在两个功能结构域上和活性位点保守。酵母表达结果显示, TaKCS6基因编码的蛋白参与C24以上超长链脂肪酸的延伸。
普通小麦;超长链脂肪酸;KCS基因;酵母表达
超长链脂肪酸(Very-long-chain fatty acids, VLCFAs)是指烃链长度超过18个碳原子的脂肪酸[1-3],其在生物体内具有多种生理功能,主要存在形式为:三酰甘油、甘油磷脂、鞘磷脂和蜡质[4]。在植物体中,超长链脂肪酸主要以三酰甘油的形式作为储藏能源存在于种子中[1, 5-6]。甘油磷脂和鞘磷脂是生物膜结构的主要成分,并且作为信号分子参与生物体内的生命活动[7]。此外,超长链脂肪酸是构成植物表皮蜡质的前体物质[2],蜡质可以保护植物免受包括干旱、冷害、空气污染、紫外光照射、病菌侵染等多种逆境胁迫的伤害[8]。
超长链脂肪酸的生物合成分为两个阶段,首先在质体中经过从头合成的16碳与18碳脂肪酸被转运到内质网上的脂肪酸延伸酶复合体(Fatty acid elongase complex),然后16碳与18碳脂肪酸经过缩合、还原、脱水和再还原4个步骤后,形成增加了2个碳原子的超长链脂肪酸[2, 9]。其中,缩合反应为限速步骤,参与该反应的β-酮脂酰CoA合成酶由KCS基因编码,具有底物特异性,决定了超长链脂肪酸的碳链长度[1, 10-11]。拟南芥KCS家族有21个基因,其中 AtFAE1/AtKCS18是第1个从拟南芥中克隆出来的KCS基因[10],突变体表现出种子中超长链脂肪酸含量大大降低[12]。 AtKCS1与 AtFAE1/AtKCS18序列相似性很高,参与蜡质的生物合成,当其被沉默时,导致蜡质中的C26和C30的醇和醛含量降低[13]。 AtKCS2/DAISY和 AtKCS20参与C20到C22超长链脂肪酸的延伸[14],参与植物角质层蜡质和根木栓质的合成[15]。 AtFDH/AtKCS10主要在花和幼嫩叶片中表达,参与表皮细胞中超长链脂肪酸的合成[16]。 AtKCS6/AtCer6/AtCUT1主要负责碳链长度在C24以上脂肪酸的合成,参与茎及花粉表皮蜡质的合成[17-18]。此外,在番茄中 AtKCS6的同源基因 SlCer6的突变导致表皮蜡质中C28以上的组分含量显著减少,果实的失水速率增快[19]。在棉花中 AtCer6同源基因的突变引起C26以上的超长链脂肪酸含量的减少,致使棉纤维伸长受到抑制[20]。因此,对KCS基因的克隆与功能分析有助于阐明植物超长链脂肪酸的生物合成机理。
小麦是主要的粮食作物,其稳产与高产具有重要意义。超长链脂肪酸作为生物体内众多物质的共同合成底物,其合成与降解对生物体正常生长发育至关重要。小麦叶片的表皮蜡质含量与保水性相关,含量越高,叶片保水性越好[21]。在水稻中研究表明,调节脂肪代谢的基因突变会影响花粉的正常生长,造成花粉不育[22]。Schneiter等[23]在酵母中的研究表明,C26的磷脂酰肌醇含量的减少,造成核孔的缺失。小麦KCS基因的功能研究还比较少,通过克隆和功能分析小麦KCS基因,能够深入了解小麦脂质代谢和蜡质合成中超长链脂肪酸的合成。本研究拟通过同源克隆方法从小麦中克隆出KCS基因后,利用生物信息学对其编码序列进行分析,并将其转化到酵母中进行表达,以期为小麦超长链脂肪酸的生物合成提供一定的理论依据。
1 材料与方法
1.1材 料
试验材料为普通六倍体小麦中国春(Chinese Spring),种植于西北农林科技大学科研温室。
1.2方 法
1.2.1小麦总RNA的提取
于苗期四叶期取小麦叶片,使用植物RNA小量提取试剂盒(Magen,R4151)提取总RNA。
1.2.2cDNA第一链的合成
取1 ng总RNA,按照PrimeScript 反转录试剂盒(TaKaRa,RR047A)说明书进行cDNA第一链的合成。
1.2.3小麦KCS基因的克隆
根据拟南芥中的KCS基因序列,分别在NCBI(https://blast.ncbi.nlm.nih.gov)和Ensembl (http://plants.ensembl.org/Triticum_aestivum/Tools/Blast)的小麦数据库中进行BLASTn搜索,寻找相似性较高的片段。利用Primer Premier 5设计引物F1/R1(F1:5′-GAAGGTCACATCATCATCATC-3′;R1:5′-TTCTGTTCCTATAGCCATTTA-3′), 交由上海生工(Sangon)生物技术公司合成。以1.2.2中得到的cDNA为模板,利用引物F1/R1进行PCR扩增。扩增体系(25 μL):2×PCR Buffer for KOD FX 12.5 μL、dNTPs(2 mmol·L-1)5 μL、KOD FX(1.0 U·μL-1)0.3 μL、上下游引物(10 μmol·L-1)各0.6 μL、cDNA 1 μL、ddH2O 5 μL。扩增程序:94 ℃预变性2 min;98 ℃变性10 s,58 ℃退火30 s,68 ℃延伸90 s,31个循环;68 ℃延伸10 min。扩增产物在160 V、200 mA的条件下,经1%琼脂糖凝胶电泳检测后,回收并纯化(TIANGEN,DP214)目的片段,加入rTaq酶和dATP,72 ℃温浴30 min,完成末端加A。将目的片段连接于pMD18-T载体(TaKaRa)上,转化大肠杆菌DH5α,筛选阳性克隆,摇菌后吸取100 μL菌液寄往上海生工生物技术公司测序。
1.2.4序列分析
利用DNAStar软件包的EditSeq程序和ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)寻找最大的开放阅读框;在NCBI网站(http://www.ncbi.nlm.nih.gov/BLAST)对核苷酸序列进行BLASTp分析;运用ProtParam(http://web.expasy.org/protparam/)对氨基酸序列进行基本理化性质分析;运用SMART(http://smart.embl-heidelberg.de/)和InterPro(http://www.ebi.ac.uk/interpro/)对蛋白的功能结构域进行分析;运用TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)进行跨膜结构域分析;运用DNAMAN 8.0软件对氨基酸序列进行同源比对;使用MEGA 6.0构建基因进化树。
1.2.5酵母表达载体的构建与酵母表达
根据酿酒酵母(Saccharomycescerevisiae)表达载体pYES2和目的基因的DNA序列,设计带酶切位点的引物F2/R2(F2:5′-CCC AAGCTTAAAAAAATGTCTATGGTCACCGT GCCCGT T-3′,下划线处为Hind III酶切位点,引入ATG;R2:5′-CCGGAATTCCAGTTAATG CTTGAAAACATCAGT-3′,下划线处为EcoR I酶切位点)。以pMD18-T- TaKCS6为模板,利用引物F2/R2扩增KCS基因完整的编码区。将扩增的片段经Hind III和EcoR I双酶切后,用T4 DNA连接酶连接到经过相同双酶切的pYES2载体上,转化大肠杆菌DH5α,经过菌落PCR鉴定和测序鉴定,确定目标基因的序列无突变后,将重组载体pYES2- TaKCS6和空载体pYES2质粒DNA分别转入酿酒酵母INVSc1菌株中,利用尿嘧啶营养缺陷型培养基(SC-Ura,北京泛基诺)筛选阳性菌株。酵母的转化和诱导表达参照Denic和Weissman[11]。
1.2.6脂肪酸的抽提及其色谱分析
将经过诱导表达的酵母菌体进行裂解,抽提脂肪酸,经甲醇酯化反应后,进行气相质谱检测。甲酯化反应如下:20 mL诱导菌液中加入20 mL 0.9%的NaCl,混匀,4 000 r·min-1离心5 min,弃上清液。加入2 mL 2.5%硫酸-甲醇溶液,80 ℃水浴1.5 h。冷却后吸取上层抽提液,置于玻璃容器中,过滤后用氮气吹干,加入正己烷回溶混合物,转移至色谱分析瓶中。利用GCMS-QP 2010气相色谱质谱联用仪对反应后的脂肪酸进行检测。毛细管色谱柱为RTX-1 (60 m×0.32 mm, 0.25 μm),进样口温度为280 ℃。毛细管柱柱箱从50 ℃开始升温,保持2 min;然后以8 ℃·min-1的速度升温至320 ℃,保持10 min。载气为高纯氮气,试验使用的分流比是2∶1,检测器温度为320 ℃,氢气流速为40 mL·min-1,空气流速为400 mL·min-1,进样量为1 μL。通过目的峰的面积计算超长链脂肪酸的相对含量。
2 结果与分析
2.1 TaKCS6基因的克隆及其序列分析
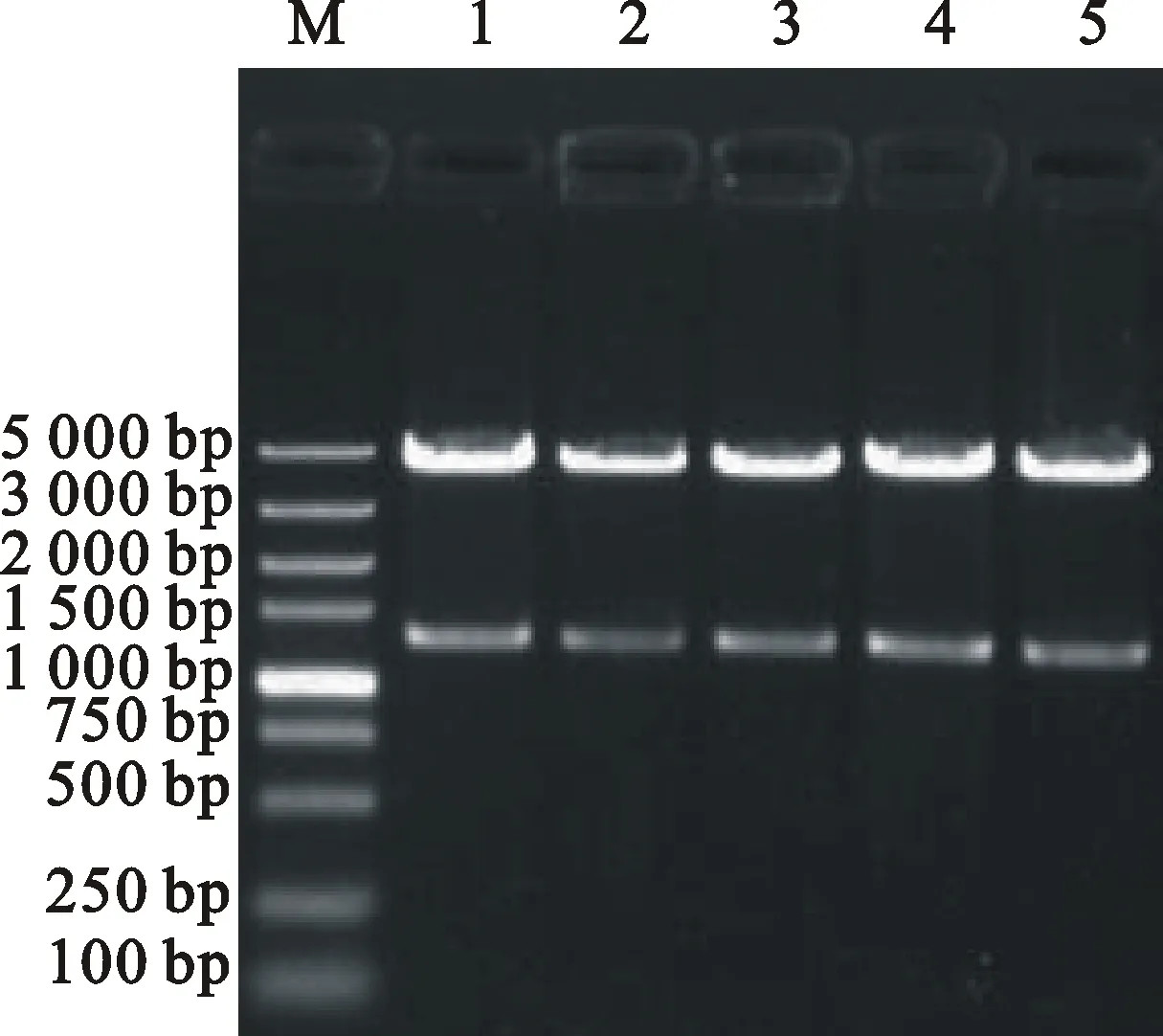
利用特异性引物F1/R1对中国春cDNA进行PCR扩增,经1%琼脂糖凝胶检测后,得到1条大小约1 300 bp的特异性条带(图1)。对目的条带进行切胶回收,将回收到的目的片段连接到pMD18-T载体上,转入大肠杆菌DH5α中,菌落PCR初步鉴定后,选取3个阳性克隆进行测序。测序结果显示,3个阳性克隆序列一致,目的片段推测的编码区全长为1 287 bp,编码428个氨基酸残基,将该基因命名为 TaKCS6。

M:DNA Marker DL2000(TaKaRa);1~3:PCR扩增产物。
M:DNA Marker DL2000(TaKaRa);1-3:PCR products.
图1 TaKCS6基因的PCR扩增结果
Fig.1PCR amplification of TaKCS6 gene
2.2TaKCS6蛋白的理化性质分析及结构与功能预测
ProtParam对TaKCS6蛋白基本理化性质分析显示,该蛋白的相对分子量为47 280.90,理论等电点pI为8.96,半衰期为30 h,在280 nm处消光系数为0.83,不稳定指数为38.50,亲脂性为92.59。说明该蛋白为稳定、亲脂性蛋白。
结构功能域分析表明,TaKCS6蛋白属于β-酮脂酰辅酶A合成酶 (KCSs,IPR012392)家族成员,含有III型聚酮合酶脂肪酸延伸酶(PF08392, 48~302 aa)和C末端3-酰基ACP合酶III (PF08541, 318~399 aa)结构域(图2),该结构域为KCS蛋白功能所必需。跨膜结构分析表明,TaKCS6蛋白具有明显的跨膜区(图3),说明该蛋白可能定位于膜上。

FAE1_CUT1_RppA:III型聚酮合酶脂肪酸延伸酶;ACP_syn_III_C:3-酰基ACP合酶III;横线下方数字表示氨基酸位置。
FAE1_CUT1_RppA:FAE1/Type III polyketide synthase-like protein; ACP_syn_III_C:3-Oxoacyl-[acyl-carrier-protein (ACP)] synthase III, C-terminal; Numbers under the line indicate the position of amino acids.
图2TaKCS6蛋白的功能域预测
Fig.2Predicted function domains of TaKCS6 protein
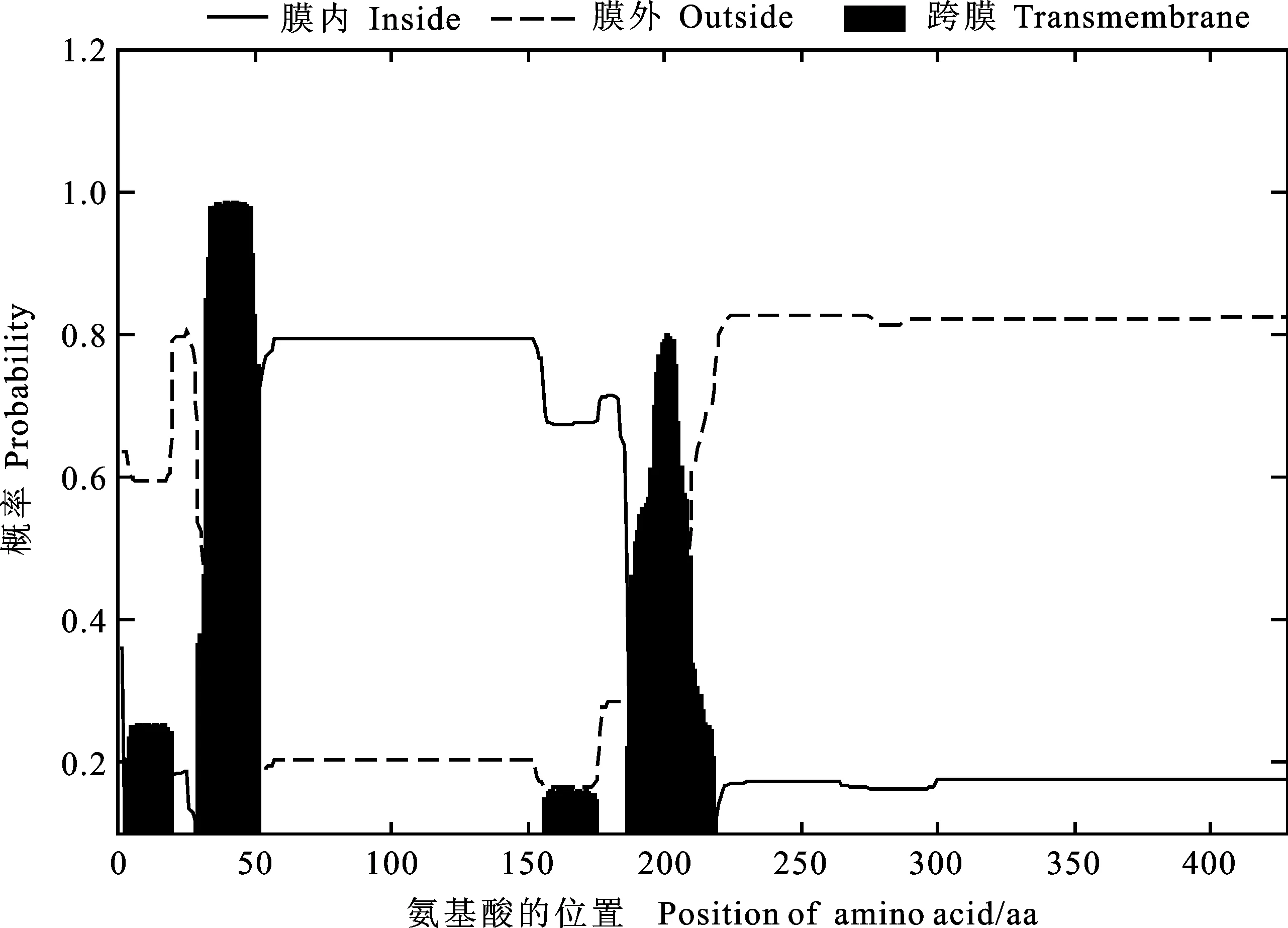
图3 TaKCS6蛋白的跨膜区预测
2.3TaKCS6蛋白的进化分析与序列比对分析
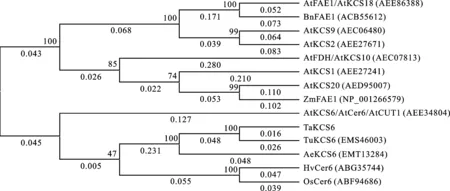
利用克隆到的 TaKCS6基因编码的氨基酸序列与拟南芥及其他6个近缘物种KCS基因编码的氨基酸序列构建系统进化树,结果(图4)发现,这14条序列主要聚类成两大类, KCS6同源基因聚为一类,其他的KCS基因聚为一类。在 KCS6同源基因分支中,来源于不同植物的 KCS6基因聚为两类,一类为双子叶植物拟南芥 AtKCS6/AtCer6/AtCUT1,另一类为单子叶植物。在单子叶植物分支中,小麦 TaKCS6与乌拉尔图小麦 TuKCS6和节节麦 AeKCS6聚为一类。
利用DNAMAN 8软件对来源于不同物种的 KCS6基因编码的氨基酸序列进行多重序列比对分析,结果(图5)表明, TaKCS6基因编码的氨基酸序列与其他物种的 KCS6基因编码的氨基酸序列在III型聚酮合酶脂肪酸延伸酶(PF08392,图5 I)和C末端3-酰基ACP合酶III (PF08541,图5II)功能结构域上具有一定的相似性。TaKCS6氨基酸序列与拟南芥AtKCS6氨基酸序列一致性为50.00%,与乌拉尔图小麦、节节麦、高粱和水稻相似性分别为71.54%、72.73%、52.77%和52.96%。TaKCS6氨基酸序列与拟南芥AtKCS6氨基酸序列的第225位半胱氨酸残基、第388位、第392位与第421位的组氨酸残基和第425位的天冬酰胺残基具有一致性,而这五个残基与第304位的组氨酸残基是拟南芥AtKCS6蛋白正常催化所必需的活性位点。
At:拟南芥;Ae:节节麦;Bn:甘蓝型油菜;Hv:高粱;Os:水稻;Tu:乌拉尔图小麦;Ta:普通小麦;Zm:玉米。下同。
At:Arabidopsisthaliana;Ae:AegilopstauschiiCoss.;Bn:BrassicanapusL.;Hv:HordeumvulgareL.;Os:Oryzasativa;Tu:Triticumurartu;Ta:TriticumaestivumL.;Zm:ZeamaysL..The same as below.
图4不同植物中KCS基因编码氨基酸序列的系统进化树分析
Fig.4Phylogenetic tree of the deduced amino acid sequences ofKCSgenes in different plants

I和II分别为AtKCS6蛋白的FAE1_CUT1_RppA(PF08392)与ACP_syn_III_C (PF08541)功能结构域;1~6分别表示 AtKCS6蛋白的C225、H304、H388、H392、H421和N425活性位点。
I and II:Function domains FAE1_CUT1_RppA(PF08392) and ACP_syn_III_C (PF08541) of AtKCS6 protein;1-6:Activity sites C225, H304, H388, H392, H421 and N425 of AtKCS6 protein.
图5TaKCS6氨基酸序列与其他植物的KCS6氨基酸序列的比对
Fig.5Multiple alignment of KCS6 amino-acid sequences inTriticumaestivumL. and other plants
2.4酵母表达载体pYES2- TaKCS6的构建及其转化诱导
将带酶切位点的 TaKCS6基因片段与环状的pYES2载体分别进行双酶切后,利用T4连接酶连接,即目的片段插入到半乳糖诱导的启动子pGAL1与终止子CYC1 TT间(图6)。将构建好的pYES2- TaKCS6载体转入大肠杆菌DH5α中,经双酶切(图7)与测序确保阅读框的正确性后,转化到酿酒酵母INVSc1菌株中,筛选得到单克隆阳性菌株。利用尿嘧啶缺陷型培养基(SC-Ura)与半乳糖培养酵母并诱导 TaKCS6基因的表达。
2.5气相质谱联用仪(GC-MS)分析酵母超长链脂肪酸的组成
提取经诱导的酵母中的脂肪酸后,使用GC-MS联用仪对甲酯化的脂肪酸进行检测,结果(图8、图9)表明,与空载体相比, TaKCS6基因表达产物中,棕榈酸(16∶0)相对含量由70.60%减少到了58.70%, 二十六酸(26∶0)相对含量由3.06%增加到了12.42%,并且检测到了二十八酸(28∶0)的积累,相对含量为2.32%。证明该基因的编码产物参与了超长链脂肪酸的延伸,使得C26以下的超长链脂肪酸向C26及以上的超长链脂肪酸延伸。鉴于二十六酸的相对含量显著增加,推测TaKCS6蛋白的特异性反应底物为C24的超长链脂肪酸。
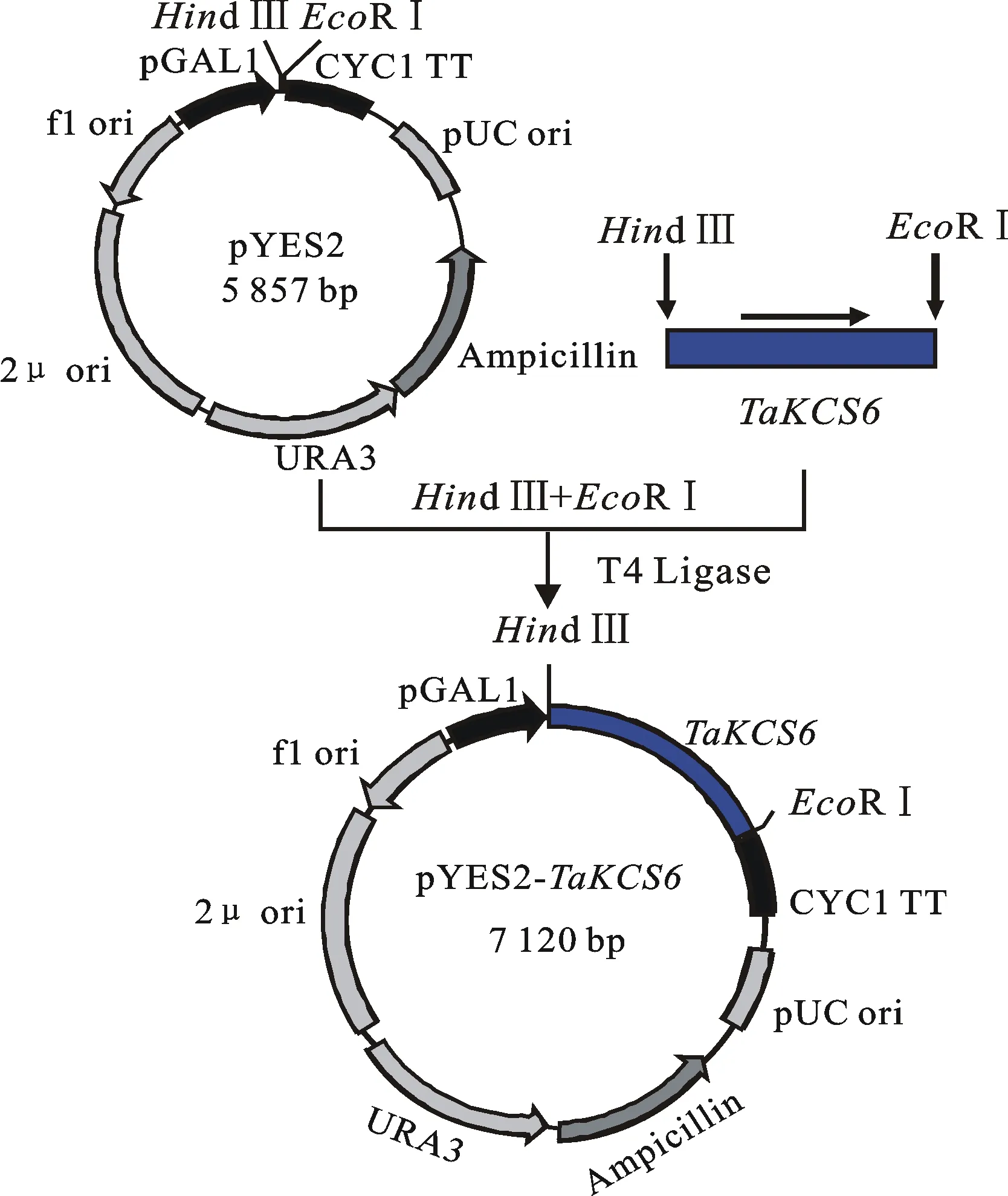
图6 酵母表达载体pYES2- TaKCS6的构建
M:DNA Marker DL5000(TaKaRa);1~4:双酶切产物。
M:DNA Marker DL5000(TaKaRa);1-4:Products of double enzyme digestion.
图7pYES2- TaKCS6的双酶切鉴定
Fig.7Double enzyme digestion of pYES2- TaKCS6

16∶0:棕榈酸;18∶0:硬脂酸;20∶0:花生酸;22∶0:山萮酸;24∶0:木蜡酸;26∶0:二十六酸;28∶0:二十八酸。下同。
16∶0:Palmitic acid; 18∶0:Stearic acid; 20∶0:Arachic acid; 22∶0:Behenic acid;24∶0:Lignoceric acid;26∶0:Hexacosanic acid;28∶0:Octacosanoic acid. The same as below.
图8酵母超长链脂肪酸的相对含量
Fig.8Relative content of very-long-chain
fatty acids in yeast
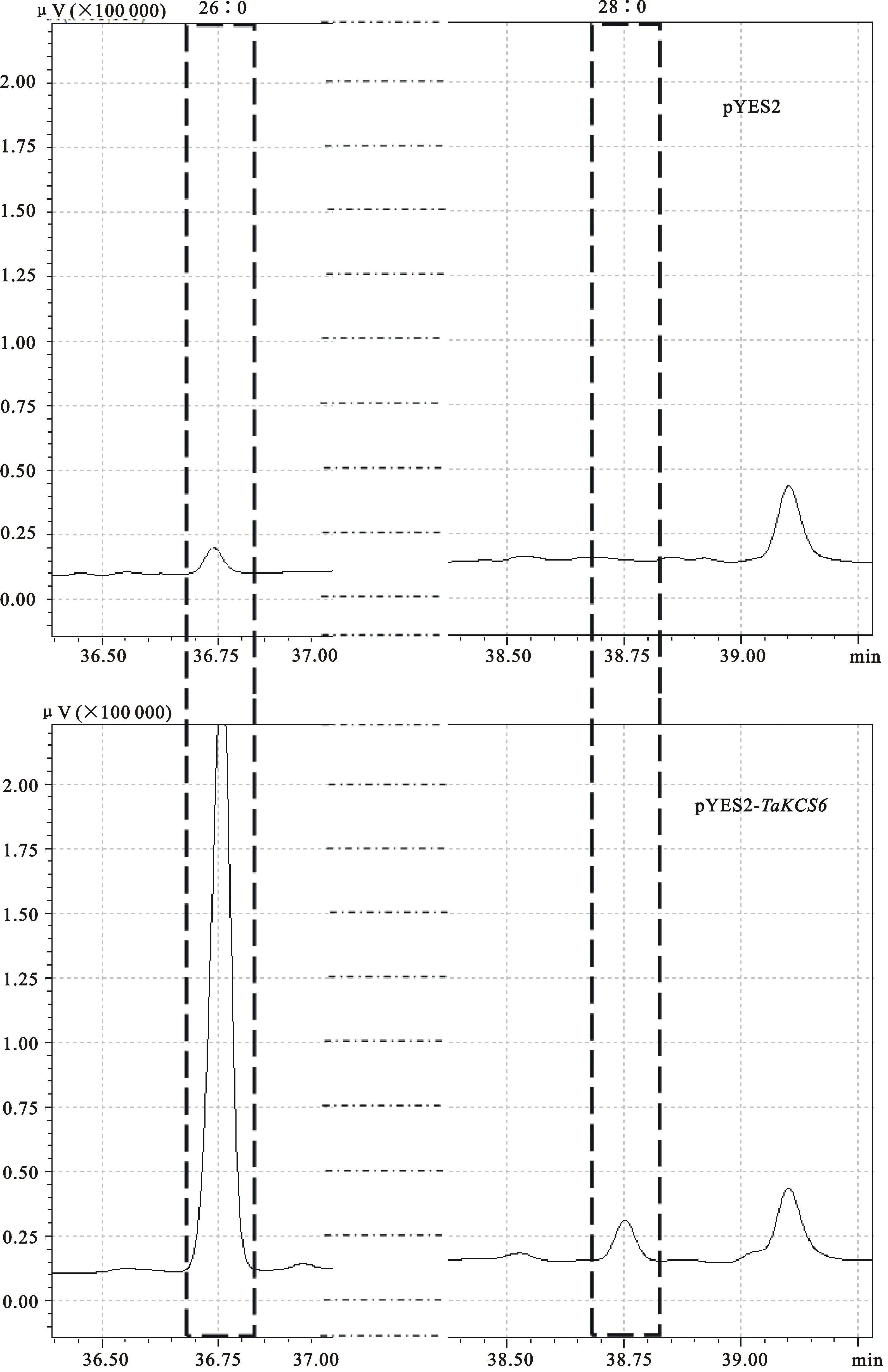
图9 GC-MS分析酵母超长链脂肪酸成分
3 讨 论
KCS基因作为VLCFAs合成过程中的关键基因,对VLCFAs的碳链长度有着决定性的作用[10-11],并参与植物体内众多重要物质的合成[4]。目前,在拟南芥、棉花、番茄、水稻等植物中克隆得到了部分KCS基因[10, 14, 16-17, 19, 24-25],并对其功能进行了分析。本研究通过同源克隆方法,克隆得到了含有完整编码序列的小麦 TaKCS6基因,序列分析表明,开放阅读框全长为1 287 bp,可编码428个氨基酸残基,具有亲脂性。结构域分析表明,TaKCS6蛋白属于β-酮脂酰辅酶A合成酶 (KCSs,IPR012392)家族,分别在N端和C端含有III型聚酮合酶脂肪酸延伸酶和3-酰基ACP合酶III结构域,并且在N段存在两个跨膜结构,这都与Hooker等[17]和Millar等[18]所克隆得到的 AtKCS6基因所编码的蛋白序列相吻合,推测TaKCS6蛋白定位于内质网,这有待进一步的研究。系统进化树分析结果表明,不同物种中的 KCS6基因聚为一类,其中小麦的 KCS6基因与乌拉尔图小麦、节节麦、水稻和高粱的 KCS6基因聚为了一类,证明这些物种之间该基因在进化上保持一致。
蛋白序列的多重比对表明,KCS6在关键的功能结构域处,序列较为保守,都存在KCS蛋白所必需的III型聚酮合酶脂肪酸延伸酶和3-酰基ACP合酶III结构域,TaKCS6蛋白序列和其他物种的KCS6蛋白与AtKCS6蛋白的5个及以上活性位点的氨基酸残基相同,表明这些蛋白的功能存在一致性,证明这些蛋白序列都具有催化合成超长链脂肪酸的功能。这与Yu等[24]的研究结果相符,表明不同植物的KCS基因都存在着极端保守的序列,这是KCS基因发挥功能所必须的。 TaKCS6基因在酵母中的表达结果表明,TaKCS6蛋白能够催化合成C24以上的超长链脂肪酸,这与Tresch等[26]在酵母中对AtKCS6表达产物的研究结果一致。
Xiao等[27]研究表明,赤霉素能够使得部分棉花的KCS基因上调表达,进而产生更多的超长链脂肪酸,影响乙烯的合成最终影响纤维的生长。Qin等[20]研究表明,超长链脂肪酸能激活乙烯的合成途径,影响棉花纤维和拟南芥茎秆的伸长。而在小麦中,Hu等[28]发现ABA与PEG诱导KCS基因的表达,表明小麦KCS基因的表达受到激素的诱导,参与植物的逆境响应。在拟南芥21个KCS基因中,有6个KCS基因[10,13-18]参与蜡质的合成,蜡质对植物的保水抗旱性有着重要的影响[8]。小麦中叶片表皮蜡质的主要成分为C28脂肪醇[29],推测TaKCS6可能参与小麦表皮蜡质的合成。
以上研究结果表明,虽然小麦 TaKCS6基因与拟南芥 AtKCS6基因的编码蛋白序列一致性不是很高,但是,在两个重要的功能结构区和活性位点相对应的氨基酸残基一致性较好,表明TaKCS6蛋白能够正常的发挥催化作用。在酵母表达中,也证明了该蛋白具有催化合成C24以上链长的超长链脂肪酸的功能。因此,TaKCS6的克隆与酵母表达对揭示小麦KCS基因的功能具有重要意义。本研究为进一步研究小麦KCS的功能及其表达模式奠定了基础。
[1]MILLAR A A,KUNST L.Very-long-chain fatty acid biosynthesis is controlled through the expression and specificity of the condensing enzyme [J].PlantJournal,1997,12(1):121-131.
[2]KUNST L,SAMUELS A L.Biosynthesis and secretion of plant cuticular wax [J].ProgressinLipidResearch,2003,42(1):51-80.
[4]BACH L,FAURE J D.Role of very-long-chain fatty acids in plant development,when chain length does matter [J].ComptesRendusBiologies,2010,333(4):361-370.
[5]VANHERCKE T,WOOD C C,STYMNE S,etal.Metabolic engineering of plant oils and waxes for use as industrial feedstocks [J].PlantBiotechnologyJournal,2013,11(2):197-210.
[6]LEONARD A E,PEREIRA S L,SPRECHER H,etal.Elongation of long-chain fatty acids [J].ProgressinLipidResearch,2004,43(1):36-54.
[7]RAFFAELE S,LEGER A,ROBY D.Very long chain fatty acid and lipid signaling in the response of plants to pathogens [J].PlantSignaling&Behavior,2009,4(2):94-99.
[8]SEO P J,LEE S B,SUH M C,etal.The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions inArabidopsis[J].PlantCell,2011,23(3):1138-1152.
[9]HASLAM T M,KUNST L.Extending the story of very-long-chain fatty acid elongation [J].PlantScience,2013,210:93-107.
[10]JAMES D W,LIM E,KELLER J,etal.Directed tagging of theArabidopsisFATTY ACID ELONGATION1 (FAE1) gene with the maize transposon activator [J].PlantCell,1995,7(3):309-319.
[11]DENIC V,WEISSMAN J S.A molecular caliper mechanism for determining very long-chain fatty acid length [J].Cell,2007,130(4):663-677.
[12]KUNST L,TAYLOR D C,Underhill E W.Fatty-acid elongation in developing seeds ofArabidopsisthaliana[J].PlantPhysiologyandBiochemistry,1992,30(4):425-434.
[13]LEE S B,SUH M C.Recent advances in cuticular wax biosynthesis and its regulation inArabidopsis[J].MolecularPlant,2013,6(2):246-249.
[14]LEE S B,JUNG S J,GO Y S,etal.TwoArabidopsis3-ketoacyl CoA synthase genes, KCS20 and KCS2/DAISY,are functionally redundant in cuticular wax and root suberin biosynthesis,but differentially controlled by osmotic stress [J].ThePlantJournal,2009,60(3):462-475.
[15]FRANKE R,HÖFER R,BRIESEN I,etal.TheDAISYgene fromArabidopsisencodes a fatty acid elongase condensing enzyme involved in the biosynthesis of aliphatic suberin in roots and the chalaza-micropyle region of seeds [J].ThePlantJournal,2009,57(1):80-95.
[16]PRUITT R E,VIELLE-CALZADA J P,PLOENSE S E,etal.FIDDLEHEAD,a gene required to suppress epidermal cell interactions inArabidopsis,encodes a putative lipid biosynthetic enzyme [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2000,97(3):1311-1316.
[17]HOOKER T S,MILLAR A A,KUNST L.Significance of the expression of the CER6 condensing enzyme for cuticular wax production inArabidopsis[J].PlantPhysiology,2002,129(4):1568-1580.
[18]MILLAR A A,CLEMENS S,ZACHGO S,etal. CUT1,anArabidopsisgene required for cuticular wax biosynthesis and pollen fertility,encodes a very-long-chain fatty acid condensing enzyme [J].PlantCell,1999,11(5):825-838.
[19]SMIRNOVA A,LEIDE J,RIEDERER M.Deficiency in a very-long-chain fatty acid beta-ketoacyl-coenzyme a synthase of tomato impairs microgametogenesis and causes floral organ fusion [J].PlantPhysiology,2013,161(1):196-209.
[20]QIN Y M,HU C Y,PANG Y,etal.Saturated very-long-chain fatty acids promote cotton fiber andArabidopsiscell elongation by activating ethylene biosynthesis [J].PlantCell,2007,19(11):3692-3704.
[21]黄 玲,张正斌,崔玉亭,等.小麦叶片蜡质含量与水分利用效率和产量的关系[J].麦类作物学报,2003,23(3):41-44.
HUANG L,ZHANG Z B,CUI Y T,etal.Relationship between wax content and water use efficiency of leaf and yield in wheat [J].JournalofTriticeaeCrops,2003,23(3):41-44.
[22]AYA K,UEGUCHI-TANAKA M,KONDO M,etal.Gibberellin modulates anther development in rice via the transcriptional regulation of GAMYB [J].PlantCell,2009,21(5):1453-1472.
[23]SCHNEITER R,BRUGGER B,AMANN C M,etal.Identification and biophysical characterization of a very-long-chain-fatty-acid-substituted phosphatidylinositol in yeast subcellular membranes [J].BiochemicalJournal,2004,381(3):941-949.
[24]YU D,RANATHUNGE K,HUANG H,etal.WaxCrystal-SparseLeaf1 encodes a β-ketoacyl CoA synthase involved in biosynthesis of cuticular waxes on rice leaf [J].Planta,2008,228(4):675-685.
[25]JUNG K H,HAN M J,LEE D,etal.Wax-deficientanther1 is involved in cuticle and wax production in rice anther walls and is required for pollen development [J].PlantCell,2006,18(11):3015-3032.
[26]TRESCH S,HEILMANN M,CHRISTIANSEN N,etal.Inhibition of saturated very-long-chain fatty acid biosynthesis by mefluidide and perfluidone,selective inhibitors of 3-ketoacyl-CoA synthases [J].Phytochemistry,2012,76:162-171.
[27]XIAO G H,WANG K,HUANG G,etal.Genome-scale analysis of the cottonKCSgene family revealed a binary mode of action for gibberellin A regulated fiber growth [J].JournalofIntegrativePlantBiology,2016,58(6):577-589.
[28]HU X,ZHANG Z,FU Z,etal.Significance of a β-ketoacyl-CoA synthase gene expression for wheat tolerance to adverse environments [J].BiologiaPlantarum,2010,54(3):575-578.
[29]张芸芸,李婷婷,孙瑜琳,等.小麦叶片表皮蜡质成分及含量分析[J].麦类作物学报,2014,34(7):963-968.
ZHANG Y Y,LI T T,SUN Y L,etal.Analysis on composition and content of leaf cuticular waxes of wheat(Triticumaestivum)detected by GC-MS [J].JournalofTriticeaeCrops,2014,34(7):963-968.
Cloning and Yeast Expression of 3-ketoacyl-coenzyme Synthase (KCS) Gene in Wheat(TriticumaestivumL.)
XIA Lingfeng, SHI Xue, YANG Haohong, LI Chunlian, WANG Zhonghua, QUAN Li
(College of Agronomy, Northwest A&F University/State Key Laboratory of Crop Stress Biology for Arid Areas, Yangling, Shaanxi 712100, China)
Very-long-chain fatty acids (VLCFAs) are the synthetic substrates for many important substances in the living body. TheKCSgene encodes β-ketoacyl CoA synthase, a rate-limiting enzyme in the synthesis of VLCFAs, catalyzing the condensation reaction of VLCFAs elongation process. In order to identify the function ofKCSgene in biosynthesis of VLCFAs, the homologous cloning strategy was used to cloneKCSgene from wheat(TriticumaestivumL.), the coding sequence was analysed by a series of bioinformatics software and expressed in yeast(Saccharomycescerevisiae).The result showed that the open reading frame of TaKCS6 gene is 1 287 bp, which encodes 428 amino acid residues. Functional domain analysis predicted that TaKCS6 protein belonging to KCSs protein family, contains FAE1_CUT1_RppA and ACP_syn_III_C domains. The amino acid sequences of TaKCS6, AtKCS6 and KCS6 from other plants are conservative in two functional domains and activity sites. Yeast expression of TaKCS6 showed that encoded protein is involved in the elongation of VLCFAs more than 24 carbons.
TriticumaestivumL.; Very-long-chain fatty acids;KCSgene;Yeast expression
2016-05-12
2016-08-15
国家自然科学基金项目(31471568);陕西省重点科技创新团队项目(2014KCT-25);西北农林科技大学唐仲英育种项目;西北农林科技大学基本科研业务费一般项目(QN2013006)
E-mail:laughingxia@126.com
王中华(E-mail:zhonghuawang@nwsuaf.edu.cn);权 力(E-mail:lquan@nwafu.edu.cn)
S512.1;S330
A
1009-1041(2016)09-1121-09
网络出版时间:2016-08-31
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160831.1648.002.html
猜你喜欢
国际放射医学核医学杂志(2020年4期)2020-07-27
广州大学学报(自然科学版)(2019年1期)2019-05-07
上海农业学报(2017年3期)2017-04-10
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年5期)2016-08-20
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
中国科技信息(2015年2期)2015-11-16
现代检验医学杂志(2015年2期)2015-02-06
天然产物研究与开发(2014年6期)2014-04-27
