
不同混生地被竹光合和荧光特征比较
2016-10-26 09:42:53高贵宾吴志庄吴良如潘雁红田新立
福建农林大学学报(自然科学版) 2016年5期
高贵宾, 钟 浩, 吴志庄, 吴良如, 潘雁红, 田新立
(1.国家林业局竹子研究开发中心, 浙江 杭州 310012;2. 浙江杭嘉湖平原森林生态系统国家定位观测研究站,浙江 杭州 310012;3.浙江省竹子高效加工重点实验室,浙江 杭州 310012)
不同混生地被竹光合和荧光特征比较
高贵宾1,2, 钟浩1,2, 吴志庄1,2, 吴良如1,3, 潘雁红1,3, 田新立1,3
(1.国家林业局竹子研究开发中心, 浙江 杭州 310012;2. 浙江杭嘉湖平原森林生态系统国家定位观测研究站,浙江 杭州 310012;3.浙江省竹子高效加工重点实验室,浙江 杭州 310012)
选取11种优良混生地被竹,比较不同竹种在同质环境条件下的光合和荧光特征差异,以层次分析法评价高光效竹种,为进一步研究竹子光合生理生态适应机制及优良竹种多指标、多方位的综合评价提供依据.结果表明:(1)黄条金刚竹、铺地竹、美丽箬竹和白缟椎谷笹的净光合速率(Pn)均在14.00 μmol·m-2·s-1以上,白纹阴阳竹和倭竹的Pn仅为8.10 μmol·m-2·s-1左右.(2)美丽箬竹、白缟椎谷笹和黄条金刚竹等的最大光化学效率、PSⅡ实际光化学效率、光化学猝灭系数和相对电子传递速率相对较高,菲黄竹、白纹阴阳竹和倭竹等相对较低.(3)黄条金刚竹的光补偿点最低,光饱和点最高;倭竹的光补偿点最高;白纹阴阳竹的光饱和点最低;白纹阴阳竹、菲白竹和倭竹的非光化学猝灭系数和非调节性能量耗散量子产量相对较大,美丽箬竹、白缟椎谷笹、黄条金刚竹和阔叶箬竹相对较小.可见,在同质环境条件下,不同竹种的光合生产能力、光合作用效率、光利用和光适应各有差别.综合评价显示,黄条金刚竹、美丽箬竹和白缟椎谷笹等为高光效竹种,但竹种的光合生理差异还可能与长期环境选择、遗传进化及竹子生态适应性有关.
混生竹; 地被竹; 光合特征; 荧光特征; 综合评价
竹类植物为当前无性系植物与环境相互作用关系研究的热点[1],按地下茎构型差异分为3种类型:合轴丛生型、复轴混生型和单轴散生型.混生地被竹属于以无性繁殖为主,地下茎为复轴型的地被类竹种,在竹子系统进化过程中处于合轴丛生竹向单轴散生竹演化的过渡位置,对生态环境因子的敏感性及生理、生长可塑性很高[2];同时,地被竹无性繁殖能力十分强大,是优良的观赏经济竹种[3],拥有良好的水土保持功能和观赏价值,将来可能是替代草坪类植物的良好材料.植物本身面临的生存环境是复杂多变的,在众多环境因子中,光是植物生长的能源,是对植物光合作用最重要和影响最大的环境因素,植物只有通过光合作用才能完成有机物质合成、能量贮存与转化,是高光效竹种选育的关键指标[4].目前,关于混生地被竹光合生理的研究相对较少,主要集中在两个方面:一是环境胁迫(包括臭氧、干旱、酸雨、盐、寒冷、Pb和Cd 等)对混生地被竹光合生理的影响研究[5-7];二是混生地被竹光合生理对外源物质(包括外源纳米TiO2、纳米SiO2、NO和La等)的响应研究[8-9],缺乏不同竹种光合能力比较及综合评价方面的研究.本试验选取11种优良混生地被竹种,在同质环境条件下进行光合性状、光响应特征和荧光参数等测试比较,试图了解竹子光合生理及其对环境的适应情况;通过层次分析方法,综合评价竹子对光照的适应和利用效率,旨在完善竹子良种的综合评价体系,为优质竹种选育和高效栽培技术提供依据.
1 材料与方法
1.1试验地概况
试验地设在位于“竹子之乡”的浙江省临安市(29°56′~30°23′N,118°51′~119°72′E)太湖源镇太湖源观赏竹种园苗圃.试验地属亚热带季风气候,温暖湿润,四季分明,年降水量1 250~1 600 mm,年平均气温15.4 ℃,1 月份平均气温3.2 ℃,7月份平均气温29.9 ℃,极端低温-13.3 ℃,极端高温40.2 ℃,全年大于10 ℃的平均活动积温5 100 ℃,年均无霜期235 d,年日照时数1 850~1 950 h.土壤为红壤,土层厚度大于60 cm,土质肥沃,结构疏松,十分适宜竹类植物生长.
1.2材料
供试材料为翠竹Sasapygmaea(Miq.) E. G. Camus(原产日本,秆高20~40 cm,地径0.1~0.2 cm)、菲黄竹SasaauricomaE. G. Camus(原产日本,秆高20~80 cm,地径0.1~0.2 cm)、菲白竹Sasafortunei(Van Houtte) Fiori(原产日本,秆高15~40 cm,地径0.1~0.2 cm)、阔叶箬竹Indocalamuslatifolius(Keng) McClure(产自山东、江苏、安徽、浙江、江西、福建、湖北、湖南、广东和四川等地,秆高150~200 cm,地径0.5~1.5 cm)、白缟椎谷笹Sasaelleaglabraf.albo-striataMuroi(原产日本,秆高20~60 cm,地径0.1~0.2 cm)、鹅毛竹ShibataeachinensisNakai(分布于江苏、浙江、江西、福建和安徽等地,秆高60~100 cm,地径0.2~0.3 cm)、倭竹Shibataeakumasasa(Zoll. ex Steud.) Makino(产自我国东南沿海地区,秆高100~200 cm,地径0.3~0.4 cm)、铺地竹SasaargenteistriatusE. G. Camus(主要分布于浙江和江苏等地,秆高30~50 cm,地径0.2~0.3 cm)、黄条金刚竹Pleioblastuskongosanensisf.aureostriaus(分布于浙江和江苏等地,秆高50~100 cm,地径0.2~0.3 cm)、美丽箬竹IndocalamusdecorusQ. H. Dai(长江以南地区广为分布,秆高30~90 cm,地径0.2~0.4 cm)和白纹阴阳竹Hibanobambustranguillansf.shiroshimaH. Okamura(原产日本,秆高150~200 cm,地径1~2 cm)等11种优良竹种.苗圃中的各竹种于2004年引种至竹种园,2008年按竹种分区连片栽种,竹种栽培采用相同质量的基质和肥料,经营管理方式一致,没有发生病虫害,虽然竹子整体生长较为致密(各竹种平均密度为1~3株·cm-2),但生长势均良好.
1.3方法
试验于2015年5月正值1年生混生地被竹抽枝展叶期间,在混生地被竹苗圃中测定11种优良竹种的光合性状、光响应曲线和荧光参数等.
1.3.1光合参数的测定参考文献[10]的方法测定.分别选取各竹种9~12株生长势良好的1年生竹株,要求竹株在苗圃地中分布合理,能够基本代表整个竹种.选择晴朗无风天气,在上午9:00~11:00,采用Li-6400光合测定仪,以1年生竹株顶端东南方向充分伸展且生长状况较为一致的竹叶作为测试样本,测定各竹种的净光合速率(Pn)、气孔导度、胞间CO2浓度和蒸腾速率,Pn与蒸腾速率之比为水分利用效率.测定时,待光合测定仪读数基本稳定时,连续记录3组数据,以便在数据统计时确认该组数据的稳定性.光响应特征测定时,选用仪器Auto Lightcurve程序,以1年生竹株顶端东南方向的健康竹叶为样本,从高到低设置光合有效辐射强度为2 000、1 600、1 200、800、400、200、100、60、20和0 μmol·m-2·s-1等10个梯度,最大和最小的等待时间分别设置为280和150 s,温度设置为28 ℃,参比室的CO2浓度稳定在400 μmol·mol-1.
1.3.2荧光参数的测定以测定光合参数的各竹种竹株叶片为测试样本,选择晴朗无风天气,采用PAM-2500调制叶绿素荧光仪,在上午9:00~11:00测定荧光参数.在光适应条件下,测定实时荧光值(Ft)、光下最小荧光值(F0′)和光下最大荧光值(Fm′);对叶片进行原位暗适应处理30 min,测定初始荧光值(F0)、最大荧光值(Fm)和最大光化学效率[(Fm-F0)/Fm].根据测得的参数计算如下指标.PSⅡ实际光化学效率[Y(Ⅱ)]=(Fm′-Ft)/Fm′;光化学猝灭系数qP=(Fm′-Ft)/(Fm′-F0′),qL=qP×F0′/Ft;非光化学猝灭系数qN=1-(Fm′-F0′)/(Fm-F0),NPQ=Fm/Fm′-1;非调节性能量耗散量子产量[Y(NO)]=1/[NPQ+1+qL×(Fm/F0-1)];相对电子传递速率(ETR) =光合有效辐射强度(PAR)×0.84×0.5×Y(Ⅱ).
1.3.3数据分析根据Bassman et al[11]的方法,以Pn=Pmax×(1-C0×e-α×PAR/Pmax)来拟合光响应曲线,并计算主要光响应特征参数.式中:α为表观量子效率(AQY),Pmax为潜在最大净光合速率,C0为度量弱光下Pn趋近于零的指标.如适合性检验方程拟合效果良好,则光补偿点=Pmax×ln(C0)/α,光饱和点则以达到90%Pmax时的光强计算,即光饱和点=Pmax×ln(90C0)/α.光合和荧光参数的平均值、标准差及各指标相关性分析等,均采用Excel和SPSS 17.0软件进行统计处理和作图,采用单因素随机区组方差分析和SNK检验方法(即q检验)进行多重比较.
选取Pn、光补偿点、光饱和点、最大光化学效率、PSⅡ实际光化学效率、非光化学猝灭系数和非调节性能量耗散量子产量等主要光合和荧光参数进行层次分析.在Yaahp V0.5.3层次分析软件中,以优良混生地被竹种为决策目标,以7个光合和荧光参数指标为中间层要素,以11个竹种为备选方案,构建层次结构模型.根据参数指标大小设置各竹种的重要性,建立一致性的判断矩阵,所得数据在Excel软件中按权重大小完成竹种排序并作图.
2 结果与分析
2.1不同混生地被竹光合特征比较
2.1.1不同竹种的光合性状从表1可见:11种混生地被竹的Pn以黄条金刚竹最高,其次分别为铺地竹、美丽箬竹和白缟椎谷笹,4种竹种Pn均在14.00 μmol·m-2·s-1以上,相互间无显著差异,这说明与其他混生地被竹相比,这4种竹种具有较高的光合生产能力;白纹阴阳竹和倭竹的Pn仅为8.10 μmol·m-2·s-1左右,竹种光合生产能力相对较低;在各竹种中,美丽箬竹的气孔导度和胞间CO2浓度最大,铺地竹的蒸腾速率最大,黄条金刚竹的水分利用效率最大,菲白竹、翠竹、白纹阴阳竹和倭竹的光合参数则相对较小.
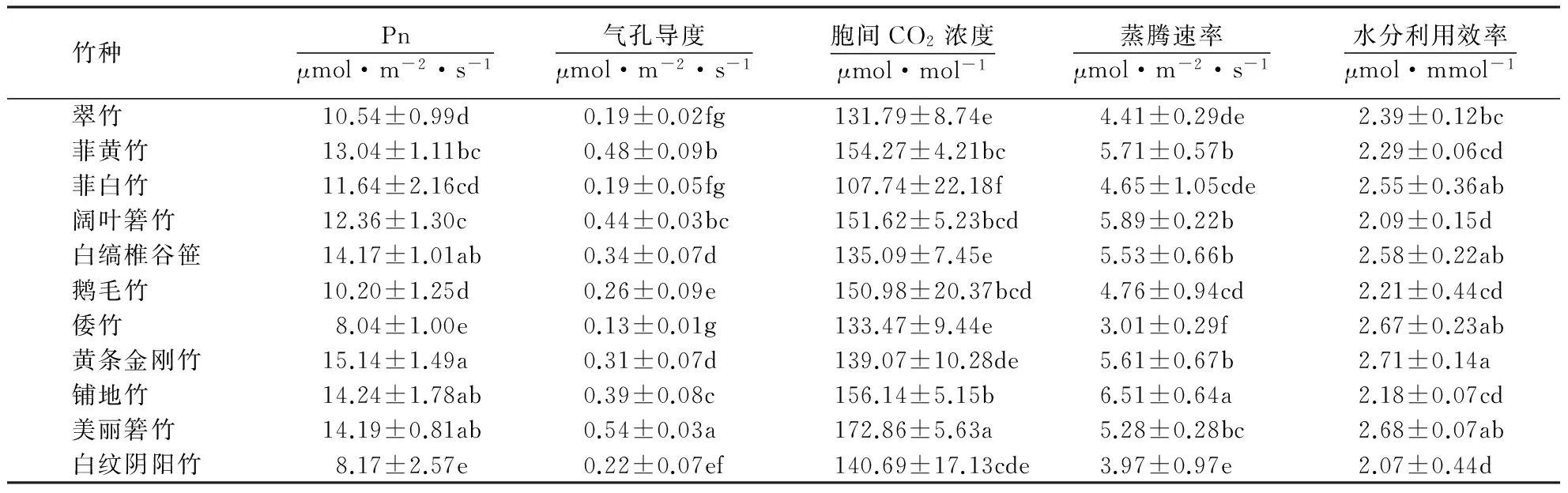
表1 竹种的光合性状1)Table 1 Photosynthesis characteristics of different bamboo
1)同列数据后附不同小写字母者表示差异显著(P<0.05),a→g表示差异显著性逐渐增强;附相同字母者表示差异不显著(P>0.05).
对竹种光合性状进行相关性分析的结果(表2)显示:竹种的Pn与气孔导度呈显著正相关,与蒸腾速率呈极显著正相关;气孔导度与胞间CO2浓度、蒸腾速率呈极显著正相关;水分利用效率与Pn呈正相关,与气孔导度、胞间CO2浓度、蒸腾速率呈负相关,但相关均不显著.这说明气孔导度和蒸腾速率是Pn的重要影响因子,对竹林光合作用具有显著的促进作用,这可能与竹种气孔导度增大,能显著提高胞间CO2浓度和蒸腾速率有关.
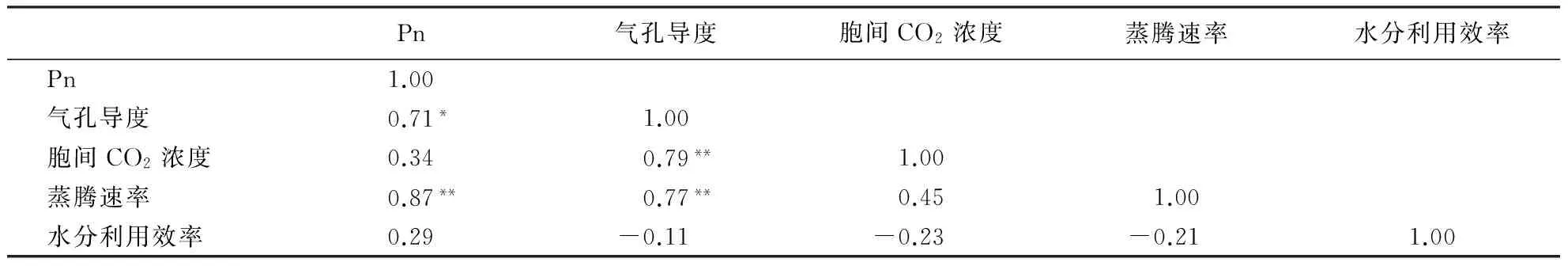
表2 竹种光合性状的相关性1)Table 2 Correlations between photosynthetic parameters
1)*表示相关显著(P<0.05),**表示相关极显著(P<0.01).
2.1.2不同竹种的光响应特征各竹种光响应曲线(图1)显示:光合有效辐射强度从0 μmol·m-2·s-1升高至800 μmol·m-2·s-1时,各竹种的Pn逐渐增大,变化相对平缓.光合有效辐射强度为800 μmol·m-2·s-1时,翠竹、阔叶箬竹和白纹阴阳竹的光强逐渐达到饱和;光合有效辐射强度为1 200 μmol·m-2·s-1时,菲黄竹、菲白竹、白缟椎谷笹、鹅毛竹和倭竹的光强逐渐达到饱和;光合有效辐射强度为1 600 μmol·m-2·s-1时,黄条金刚竹、铺地竹和美丽箬竹的光强逐渐达到饱和.光合有效辐射强度升高至1 600 μmol·m-2·s-1后,各竹林的Pn以铺地竹最高,倭竹最低.说明长期环境选择造成不同竹种之间的光响应生理产生了较大差异.

图1 竹种的光响应曲线
从各竹种光响应特征参数(表3)可以看出:铺地竹的潜在最大净光合速率最大,倭竹最小,仅为7.25 μmol·m-2·s-1;黄条金刚竹的光补偿点最低,光饱和点最高;倭竹的光补偿点最高;白纹阴阳竹的光饱和点最低.各竹种对光照的利用区间值(光饱和点—光补偿点)以黄条金刚竹最大,达1 695.72 μmol·m-2·s-1,以白纹阴阳竹最小,仅为823.25 μmol·m-2·s-1.光补偿点和光饱和点分别反映了植物对弱光、强光的利用和适应能力[12].可见,黄条金刚竹、美丽箬竹和铺地竹等具有较高的光合潜力,阔叶箬竹、翠竹和倭竹等的光合潜力相对较低.各竹种的趋零参数和表观量子效率等特征参数变化相对较小.
2.2不同混生地被竹荧光特征比较
通过植物叶绿素荧光动力学可以快速、灵敏、可靠、无损伤地了解植物光合作用生理状况及其与环境的关系[13].从表4可以看出,竹种相互间的最大光化学效率、PSⅡ实际光化学效率、非调节性能量耗散量子产量和相对电子传递速率存在显著差异,光化学猝灭系数和非光化学猝灭系数无显著差异.美丽箬竹、白缟椎谷笹和黄条金刚竹等的最大光化学效率、PSⅡ实际光化学效率、光化学猝灭系数和相对电子传递速率相对较高,菲黄竹、白纹阴阳竹和倭竹等相对较低,说明美丽箬竹等具有较高的光合作用效率,而菲黄竹等的光合作用效率则相对较低.非光化学猝灭系数和非调节性能量耗散量子产量是植物光保护和光损伤的重要指标参数[14-15].白纹阴阳竹、菲白竹和倭竹的非光化学猝灭系数和非调节性能量耗散量子产量相对较大,美丽箬竹、白缟椎谷笹、黄条金刚竹和阔叶箬竹相对较小,说明白纹阴阳竹等不耐强光照,美丽箬竹等对强光照具有较高的适应性.
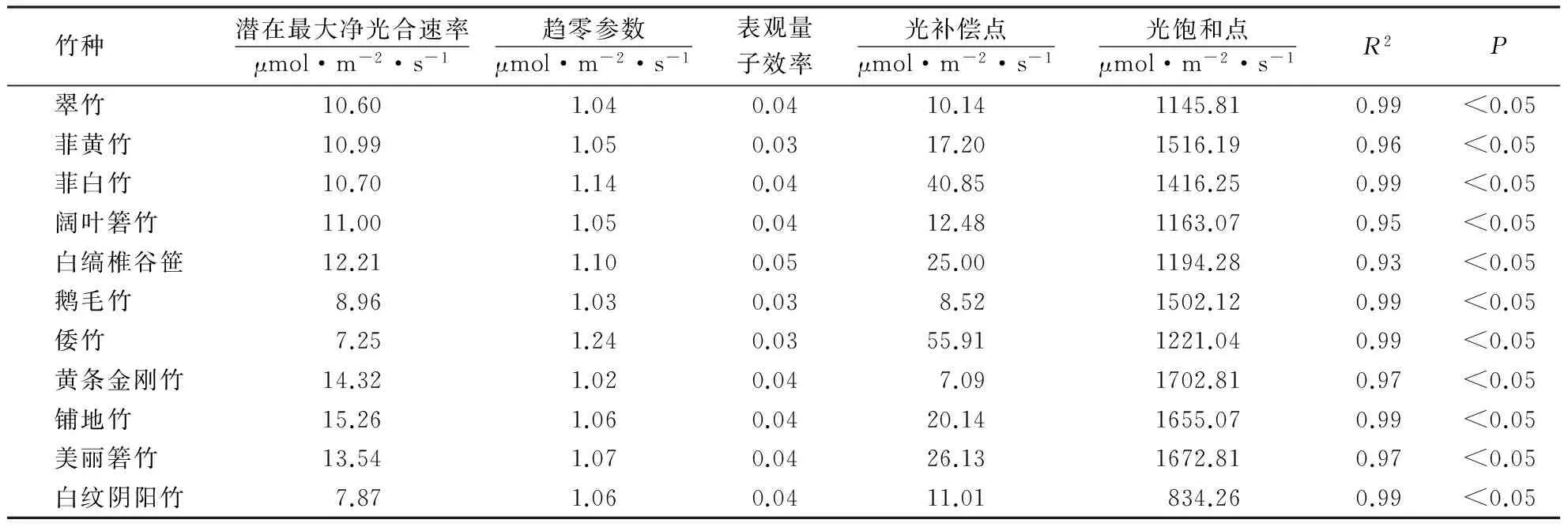
表3 竹种的光响应特征参数Table 3 Photoresponse characteristic parameters of different bamboo
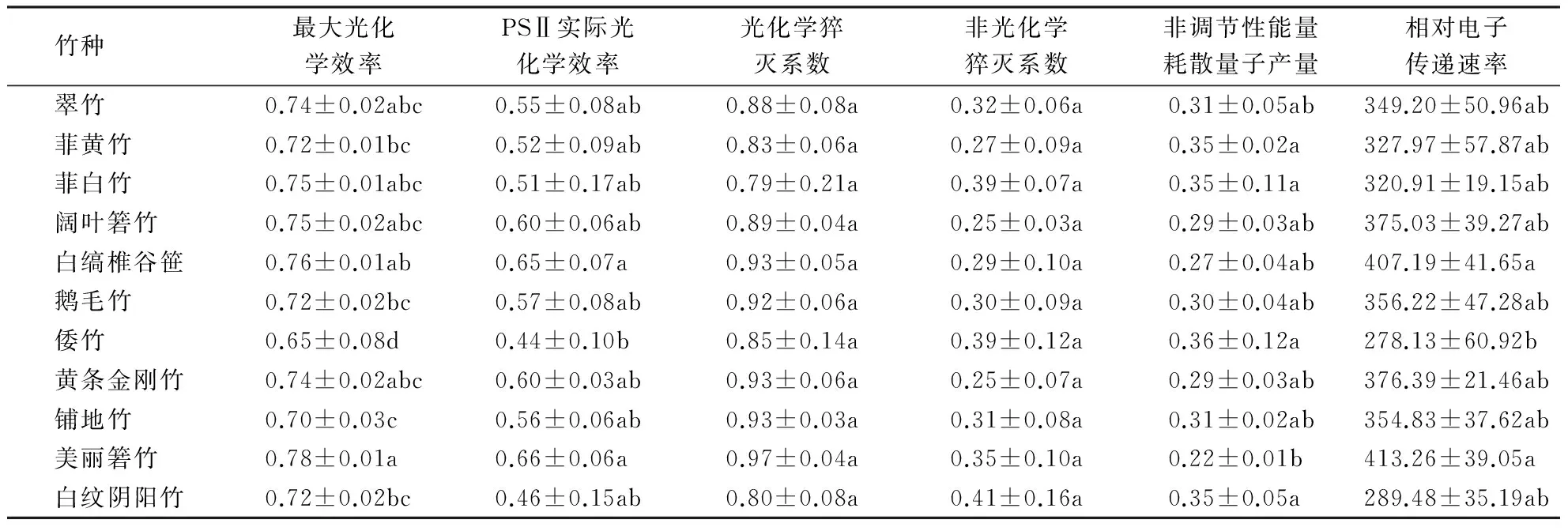
表4 竹种的荧光参数1)Table 4 Fluorescence parameters of different bamboo
1)同列数据后附不同小写字母者表示差异显著(P<0.05),a→g表示差异显著性逐渐增强;附相同字母者表示差异不显著(P>0.05).
竹种荧光参数相关性分析结果(表5)显示:最大光化学效率与PSⅡ实际光化学效率、相对电子传递速率呈极显著正相关,与非调节性能量耗散量子产量呈显著负相关,与光化学猝灭系数呈正相关,与非光化学猝灭系数呈负相关,相关不显著;PSⅡ实际光化学效率与光化学猝灭系数、相对电子传递速率呈极显著正相关,与非光化学猝灭系数、非调节性能量耗散量子产量呈显著正相关;光化学猝灭系数与相对电子传递速率呈极显著正相关,与非调节性能量耗散量子产量呈极显著负相关,与非光化学猝灭系数呈负相关,相关不显著;非光化学猝灭系数与相对电子传递速率呈显著负相关,与非调节性能量耗散量子产量呈正相关,相关不显著;非调节性能量耗散量子产量与相对电子传递速率呈极显著负相关.这说明最大光化学效率、PSⅡ实际光化学效率、光化学猝灭系数和相对电子传递速率较高,非光化学猝灭系数和非调节性能量耗散量子产量较低,竹种的光合性能较好.
2.3不同混生地被竹光合和荧光参数的层次分析
对不同混生地被竹光合和荧光主要参数进行层次分析.结果(图2)显示:以光合生产能力大、光合作用效率强和强光照适应效果好为目标,各竹种按权重由高到低排序依次为:黄条金刚竹、美丽箬竹、白缟椎谷笹、阔叶箬竹、铺地竹、鹅毛竹、菲黄竹、翠竹、菲白竹、白纹阴阳竹、倭竹.
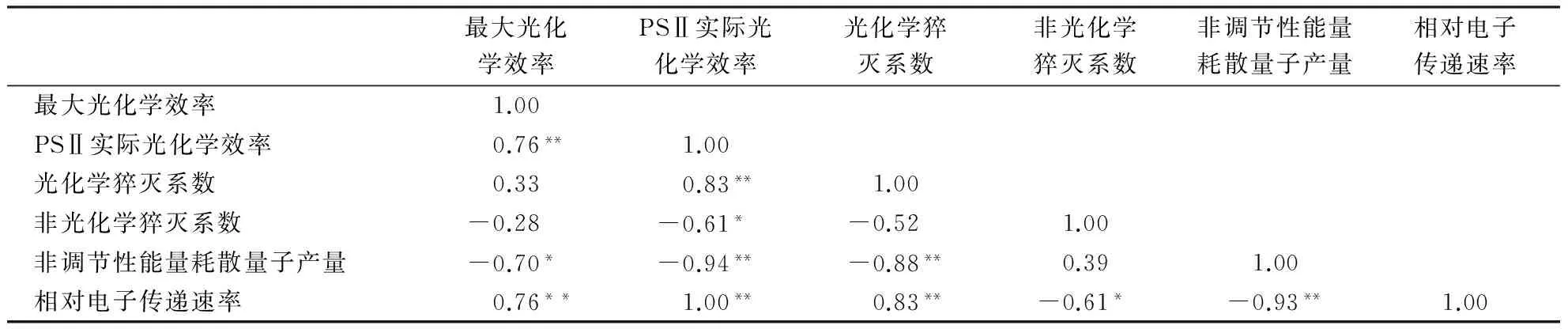
表5 竹种荧光参数的相关性1)Table 5 Correlation between fluorescence parameters of bamboo
1)*表示相关显著(P<0.05),**表示相关极显著(P<0.01).
3 结论与讨论
3.1不同竹种光合生产能力和光合作用效率比较

图2 竹种的层次分析结果Fig.2 Analytic hierarchy process among different bamboo
Pn是影响光合生产能力的重要因素,反映了植物同化物生产能力的强弱[16].研究发现,限制Pn 的因素主要分为两大类:一类是气孔因素阻止了CO2的供应,主要受气孔数量、气孔孔径和气孔开度等的影响;另一类为非气孔因素,主要受细胞内部的酶活力和光合组分控制[17-18].本试验中,竹种的Pn与气孔导度呈显著正相关,与蒸腾速率呈极显著正相关,而气孔导度与胞间CO2浓度、蒸腾速率呈极显著正相关.植物可以通过改变气孔的开闭程度来控制与外界CO2和水汽的交换,从而调节光合速率和蒸腾速率[19],这说明在本试验中,竹种叶片通过提高胞间CO2流通量,加速水分运转,对Pn产生了促进作用.通过比较,11种混生地被竹按光合生产能力的强弱由高到低排序依次为:黄条金刚竹、铺地竹、美丽箬竹、白缟椎谷笹、菲黄竹、阔叶箬竹、菲白竹、翠竹、鹅毛竹、白纹阴阳竹、倭竹.研究表明:最大光化学效率、PSⅡ实际光化学效率、光化学猝灭系数和相对电子传递速率等参数已经被公认为植物叶片光合作用效率的重要依据[20].本试验结果表明,竹种的最大光化学效率、PSⅡ实际光化学效率、光化学猝灭系数和相对电子传递速率等参数间基本呈显著正相关,这说明其相互间具有明显的协同促进作用.比较发现,美丽箬竹、白缟椎谷笹和黄条金刚竹等具有较高的光合作用效率,而菲黄竹、白纹阴阳竹和倭竹等的光合作用效率则相对较低.
3.2不同竹种光利用和光适应比较
光饱和点和光补偿点是植物两个重要的光合生理指标,反映了植物对光照的利用和适应能力,是判断植物耐阴性的重要指标[21].光补偿点低且光饱和点相应也低的植物具有很强的耐阴性;光补偿点低,而光饱和点较高的植物,能适应多种光照环境;光补偿点较高,而光饱和点较低的植物,应栽植于侧方遮阴或部分时段荫蔽的环境;光饱和点和光补偿点均较高的植物则为喜光的阳生植物.研究表明,白纹阴阳竹和翠竹属于相对耐阴的竹种;黄条金刚竹和鹅毛竹的光照利用率相对较高,倭竹的光照利用率则相对较低;菲白竹、美丽箬竹和铺地竹等属于相对喜阳竹种.这说明不同竹种在同质环境中的光响应特征存在较大差异,植物对同质环境的趋同进化是一个缓慢的过程.
本试验结果显示,在同样光照环境条件下,白纹阴阳竹、菲白竹和倭竹等的非光化学猝灭系数和非调节性能量耗散量子产量相对较高,表明这些竹种启动了PSⅡ热耗散及非调节性能量耗散功能,以减少因过多吸收光能向PSⅡ反应中心传递而引起PSⅡ的光抑制或光破坏,这与孙启祥[22]的研究结果相近.本试验结果还显示:黄条金刚竹、美丽箬竹和铺地竹等具有较高的光合潜力,阔叶箬竹、翠竹和倭竹等的光合潜力相对较低;白纹阴阳竹、菲白竹和倭竹等不耐强光照,而美丽箬竹、白缟椎谷笹、黄条金刚竹和阔叶箬竹等则对强光照具有较高的适应性.
3.3不同竹种光合能力的综合评价
近年来,育种学家已普遍关注植物光合作用效率品种间的差异,积极寻找在遗传特性方面具有高光合作用效率的品种,虽然取得了一些进展,但始终没有重大突破,其根本原因在于光合作用是一个非常复杂的理化过程.光合作用主要由原初反应、光合磷酸化、电子传递、碳同化以及光合产物的合成等几个主要环节组成,各个环节互相配合、互相制约,而各个环节在不同情况下的制约作用大小是不同的[23].本试验通过比较各竹种光合和荧光特征发现,在竹子光合生产能力、光合作用效率、光利用和光适应等方面,不同竹种间各有差异和优劣势.有的竹种光合生产能力很高,但光合作用效率却相对较低;有的竹种光适应能力很强,但光合生产能力却相对较低.有研究认为,环境异质性是自然界的普遍属性[24-25],由此可以推测,自然界光照环境的异质性,可能会造成竹种间,乃至同一竹种的光合生产能力、光合作用效率和光适应等生理过程发生复杂多样的变化,仅通过某一方面的光合参数很难全面了解竹种的光合生理机制,也不能对其光利用和光适应进行科学评价.因此,本试验采用层次分析法综合分析了不同混生地被竹光合和荧光的主要参数,以光合生产能力高、光合作用效率强和强光照适应效果好为目标,11种竹种光合能力由高到低排序依次为:黄条金刚竹、美丽箬竹、白缟椎谷笹、阔叶箬竹、铺地竹、鹅毛竹、菲黄竹、翠竹、菲白竹、白纹阴阳竹、倭竹.但同质环境条件下竹种的光合生理差异,还可能与竹种长期环境选择、遗传进化以及竹子的生态适应性有关,其详细机制尚待进一步深入研究.
[1] 施建敏,叶学华,陈伏生,等.竹类植物对异质生境的适应——表型可塑性[J].生态学报,2014,34(20):5 687-5 695.
[2] 江泽慧.世界竹藤[M].沈阳:辽宁科学技术出版社,2002.
[3] 刘国华,王福升,丁雨龙,等.4种地被竹光合作用日变化及光合光响应曲线[J].福建林学院学报,2009,29(3):258-263.
[4] 吴志庄,高贵宾,熊德礼,等.9个主要丛生竹种光响应曲线的研究[J].竹子研究汇刊,2013,32(2):16-20.
[5] 李应,陈双林,谢双喜,等.3种地被竹对大气臭氧胁迫的光合生理响应[J].热带亚热带植物学报,2012,20(3):263-269.
[6] 李娟,彭镇华,高健,等.干旱胁迫下黄条金刚竹的光合和叶绿素荧光特性[J].应用生态学报,2011,22(6):1 395-1 402.
[7] 谢寅峰,杨万红,陆美蓉,等.模拟酸雨胁迫下硅对髯毛箬竹光合特性的影响[J].应用生态学报,2008,19(6):1 179-1 184.
[8] 谢寅峰,李博,张千千,等.纳米TiO2对髯毛箬竹光合特性的影响[J].东北林业大学学报,2011,39(3):22-25.
[9] 谢寅峰,杨万红,杨阳,等.外源一氧化氮对模拟酸雨胁迫下箬竹(Indocalamusbarbatus)光合特性的影响[J].生态学报,2007,27(12):5 193-5 201.
[10] 吴志庄,杜旭华,熊德礼,等.不同类型竹种光合特性的比较研究[J].生态环境学报,2013,22(9):1 523-1 527.
[11] BASSMAN J H, ZWIER J C. Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltoidsandPopulustrichocarpa×P.deltoidsclones [J]. Tree Physiology, 1991,8(2):145-149.
[12] 徐克章.植物生理学[M].北京:中国农业出版社,2007:163-164.
[13] 蔡楚雄,邓雄,曹洪麟,等.6个芒果品种的叶绿素荧光日变化研究初报[J].广东农业科学,2003(2):17-19.
[14] 郑蓉,黄耀华,连巧霞,等.刚竹属13个竹种叶绿素荧光特性比较[J].江西农业大学学报,2008,30(2):263-267.
[15] KRAMER D M, JOHNSON G, KIIRATS O, et al. New fluorescence parameters for the determination of q (a) redox state and excitation energy fluxes [J]. Photosynthesis Research, 2004,79(2):201-218.
[16] 潘瑞炽.植物生理学[M].6版.北京:高等教育出版社,2008:56-103.
[17] QUICK W P, CHAVES M M, WENDLER R, et al. The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions [J]. Plant, Cell and Environment, 1992,15(1):25-35.
[18] LAL A M, KU S B, EDWARDS G E. Analysis of inhibition of photosynthesis due to water stress in the C3speciesHordeumvulgareandViciafaba: electron transport, CO2fixation and carboxylation capacity [J]. Photosynthesis Research, 1996,49(1):57-69.
[19] 刘娟,马媛,廖康,等.新疆主栽杏品种的光响应曲线[J].经济林研究,2012,30(1):45-50.
[20] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002:29-35.
[21] 马兰涛,陈双林.Guaduaamplexifolia光合季节动态[J].广西植物,2013,33(4):475-481.
[22] 孙启祥.竹子光抑制特征研究[D].北京:中国林业科学研究院,2004:41-50.
[23] 张蕾,金松恒,方伟.4种珍稀观赏竹光合作用特性研究[J].浙江大学学报(农业与生命科学版),2009,35(2):165-172.
[24] HE W M, ALPERT P, YU F H, et al. Reciprocal and coincident patchiness of multiple resources differentially affect benefits of clonal integration in two perennial plants [J]. Journal of Ecology, 2011,99(5):1 202-1 210.
[25] 王晗生.黄土高原环境异质性与植被的恢复与重建[J].生态学报,2009,29(5):2 445-2 455.
(责任编辑:施晓棠)
Comparative study on photosynthetic and fluorescence characteristics of different amphipodial dwarf bamboo
GAO Guibin1,2, ZHONG Hao1,2, WU Zhizhuang1,2, WU Liangru1,3, PAN Yanhong1,3, TIAN Xinli1,3
(1.China National Bamboo Research Center, Hangzhou, Zhejiang 310012, China; 2.National Long-term Observation and Research Station for Forest Ecosystem in Hangzhou-Jiaxing-Huzhou Plain, Hangzhou, Zhejiang 310012; 3.Key Laboratory of High Efficient Processing of Bamboo of Zhejiang Province, Hangzhou, Zhejiang 310012, China)
To elucidate adaptive mechanism of dwarf bamboo on photosynthetic and fluorescence characteristics, and establish parameters of comprehensive evaluation for fine varieties, 11 types of amphipodial dwarf bamboo which were grown in the same condition were selected for photosynthetic and fluorescence analysis. Results showed that: (1) all the net photosynthetic rates (Pn) ofPleioblastuskongosanensisf.aureostriaus,Sasaargenteistriatus,IndocalamusdecorusandSasaelleaglabraf.albo-striatawere over 14.00 μmol·m-2·s-1, but Pn rates ofHibanobambustranguillansf.shiroshimaandShibataeakumasasawere around 8.10 μmol·m-2·s-1. (2) Maximum photochemical quantum yield of photosystem II (PSII), effective photochemical quantum yield of PSII, coefficient of photochemical fluorescence quenching and relative electron transport rate (ETR) of IndocalamusI.decorus,S.glabraf.albo-striata,P.kongosanensisf.aureostriaus(in a descending order) were higher thanSasaauricoma,H.tranguillansf.shiroshimaandS.kumasasa; (3)P.kongosanensisf.aureostriaushad the lowest light compensation point and the highest light saturation point whileS.kumasasahad the highest light compensation point andH.tranguillansf.shiroshimahad the lowest light saturation point. Coefficient of photochemical fluorescence quenching, quantum yield of non-light induced non-photochemical fluorescence quenching ofH.tranguillansf.shiroshima,SasafortuneiandS.kumasasawere relatively high but those ofI.decorus,S.glabraf.albo-striata,P.kongosanensisf.aureostriausandIndocalamuslatifoliuswere relatively low. To summarize, photosynthetic productivity and efficiency, light utility and adaptation varied significantly among species even when they were grown in the same environment.P.kongosanensisf.aureostriaus,I.decorusandS.glabraf.albo-striatawere species of high photosynthetic efficiency. Differences in photosynthetic characteristics of bamboo species were results of long-term environment selection, genetic evolution and ecological adaptation. Therefore, further study might be required for thorough investigation.
amphipodial bamboo; dwarf bamboo; photosynthesis characteristic; fluorescence characteristic; comprehensive evaluation
2015-10-08
2015-11-30
中央级公益性科研院所基本科研业务费专项资金资助项目(CAFYBB2014QA038);浙江省自然科学基金资助项目(LY14C030008);浙江省科技计划项目(2014F10047).
高贵宾(1982-),男,助理研究员,硕士.研究方向:竹类植物生态.Email:anshu998@163.com.
S718.51+2.2
A
1671-5470(2016)05-0515-07
10.13323/j.cnki.j.fafu(nat.sci.).2016.05.007
猜你喜欢
世界竹藤通讯(2023年2期)2023-05-12 09:27:08
世界竹藤通讯(2022年4期)2022-11-28 09:55:26
食品工业科技(2022年9期)2022-04-26 10:33:28
江苏农业科学(2020年12期)2020-07-20 03:26:38
西安科技大学学报(社会科学版)(2019年1期)2019-09-10 07:22:44
世界竹藤通讯(2015年4期)2015-12-29 10:46:38
世界竹藤通讯(2015年4期)2015-12-29 10:46:36
东北林业大学学报(2015年2期)2015-03-08 05:44:46
新课程·中旬(2014年10期)2014-12-26 04:47:30
世界竹藤通讯(2010年2期)2010-08-15 00:50:49
