
谷子扩展蛋白与品种抗旱性关系的研究
2016-10-21 01:07苏彦冰马芳芳温思钰刘锐禾璐刘龙龙李红英韩渊怀
山西农业大学学报(自然科学版) 2016年10期
苏彦冰,马芳芳,温思钰,刘锐,禾璐,刘龙龙,李红英,韩渊怀,2,*
(1.山西农业大学 农学院,山西 太谷 030801; 2.农业部黄土高原作物基因资源与种质创制重点实验室,山西 太原 030031;3.杂粮种质资源发掘与遗传改良山西省重点实验室,山西 太谷 030801)
谷子扩展蛋白与品种抗旱性关系的研究
苏彦冰1,马芳芳1,温思钰1,刘锐1,禾璐1,刘龙龙3,李红英1,韩渊怀1,2,3*
(1.山西农业大学 农学院,山西 太谷 030801; 2.农业部黄土高原作物基因资源与种质创制重点实验室,山西 太原 030031;3.杂粮种质资源发掘与遗传改良山西省重点实验室,山西 太谷 030801)
[目的]扩展蛋白是植物细胞壁的重要组成部分,普遍存在于植物中,除了具有增加细胞壁柔韧性的作用外,在植物的生长发育及抵御生物和非生物胁迫等方面都起着重要的作用。为发掘谷子抗旱基因。[方法]本文采用生物信息学手段结合表达验证,分析了谷子扩展蛋白基因家族中的16个成员。[结果]发现它们分属于EXPA、EXPB、EXPLA和EXPLB 4个亚家族;对谷子扩展蛋白基因进行启动子分析,发现了与干旱胁迫相关的响应元件,包括干旱响应MYB结合位点MBS、脱落酸响应元件ABRE等;通过对两个谷子品种勾勾母鸡咀(GG,耐干旱品种)和晋汾16(JF16,干旱敏感品种)扩展蛋白基因表达的分析比较,发现干旱胁迫下不同谷子品种中同一基因的表达水平存在显著差异。[结论]结合亲缘关系图及启动子顺式调控元件的预测结果,并未发现基因亲缘关系与扩展蛋白基因表达间的规律;也未发现胁迫相关的调控元件与扩展蛋白表达之间的关联,初步了解了扩展蛋白基因与谷子抗旱的关系,为发掘谷子抗旱基因提供基础。
谷子;扩展蛋白;干旱胁迫;抗旱性
谷子(Setariaitalica),禾本科狗尾草属,是我国北方重要的杂粮作物,主要产区分布在河北省、山西省、陕西省和内蒙古自治区[1]。随着人们饮食结构的变化,对小米的需求不断增加,市场缺口逐渐扩大,但由于华北地区干旱情况十分严重,当谷子遇到干旱胁迫时其生理生化都会发生变化以适应干旱环境,导致低产等状况[2]。所以发掘谷子抗旱相关基因,提高谷子抗旱性从而增加谷子产量,成为当前对谷子抗旱研究亟待解决的重要问题之一。
扩展蛋白(Expansin)是一类存在于植物细胞壁中的松弛蛋白,具有松弛和修饰细胞壁的功能,广泛存在于植物中,是植物细胞壁的重要组成部分。又称膨胀素[3]。1989年Cosgrove等人从黄瓜(Cucumissativus)的根尖细胞壁中提取出扩展蛋白[4];1992年McQueen-Mason等人将该类蛋白分离提纯[5,6]。扩展蛋白一直以来备受关注,现已证明,扩展蛋白在植物生长发育及植物抗逆性过程中[7~9]具有重要作用。如,扩展蛋白基因的表达对水分胁迫产生响应,植物在干旱胁迫下通过提高扩展蛋白基因的表达水平,提高扩展蛋白含量,以增加细胞壁的韧性,从而舒缓水分胁迫对植物细胞造成的压力[10]。
目前关于拟南芥[11]、水稻[12]和商陆[8]等物种扩展蛋白的研究已经出现,但谷子中关于其抗旱的分子机制和抗旱基因的研究相对较少,尤其对于谷子扩展蛋白基因家族的研究鲜有报道。本文拟通过对谷子扩展蛋白基因进行生物信息学分析,并对勾勾母鸡咀(GG,耐干旱品种)和晋汾16(JF16,干旱敏感品种)[13]两个谷子品种在PEG模拟干旱胁迫下扩展蛋白基因的表达情况进行分析,旨在为后续该蛋白的功能鉴定和研究奠定理论基础。
1 材料与方法
1.1研究材料
本文研究材料勾勾母鸡咀和晋汾16由山西农业大学农业生物工程研究所提供[13]。
1.2研究方法
1.2.1谷子扩展蛋白基因的生物信息学分析
利用Phytozome v 11.0中豫谷1号基因组数据库(https://phytozome.jgi.doe.gov/pz/portal.html)进行关键词搜索,进一步结合NCBI、GenBank等数据库获取谷子扩展蛋白基因的信息,并下载整理。
使用phytozome v 11.0中JBrowse功能下载豫谷1号扩展蛋白基因在染色体上的位置信息。
使用MEGA 7.0软件中的Neibor-joining算法对下载的谷子扩展蛋白氨基酸序列构建系统发育树。
利用DNAMAN 6.0预测蛋白质分子量和等电点等理化性质。
通过PlantCARE网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线分析启动子元件。
1.2.2干旱胁迫下谷子扩展蛋白基因的表达情况分析
GG和JF16在人工气候室中培养,采用营养土∶蛭石= 3∶1的混合土种植,培养于14 h光照28 ℃/10 h黑暗23 ℃的条件下,出苗后两天浇一次蒸馏水,培养21 d后,釆用20% PEG-6000模拟干旱,处理材料0.5 h,对照用蒸馏水处理相同时间,每个处理取3株用于RNA的提取,进行表达谱测序。
2 结果与分析
2.1谷子扩展蛋白基因的基本信息
本文研究的16个谷子扩展蛋白基因分别属于EXPA、EXPB、EXPLA、EXPLB 4个亚家族(表1),利用豫谷1号的氨基酸序列制作了16个扩展蛋白的亲缘关系分布图(图1)。进一步结合NCBI获取的信息发现,各亚家族成员会聚集到一起。其中扩展蛋白A亚家族基因7个;扩展蛋白B亚家族基因5个;扩展蛋白LB亚家族基因和LA亚家族基因各2个。

图1 谷子扩展蛋白基因家族系统进化树Fig.1 Phylogenetic tree of expansion family in foxtail millet
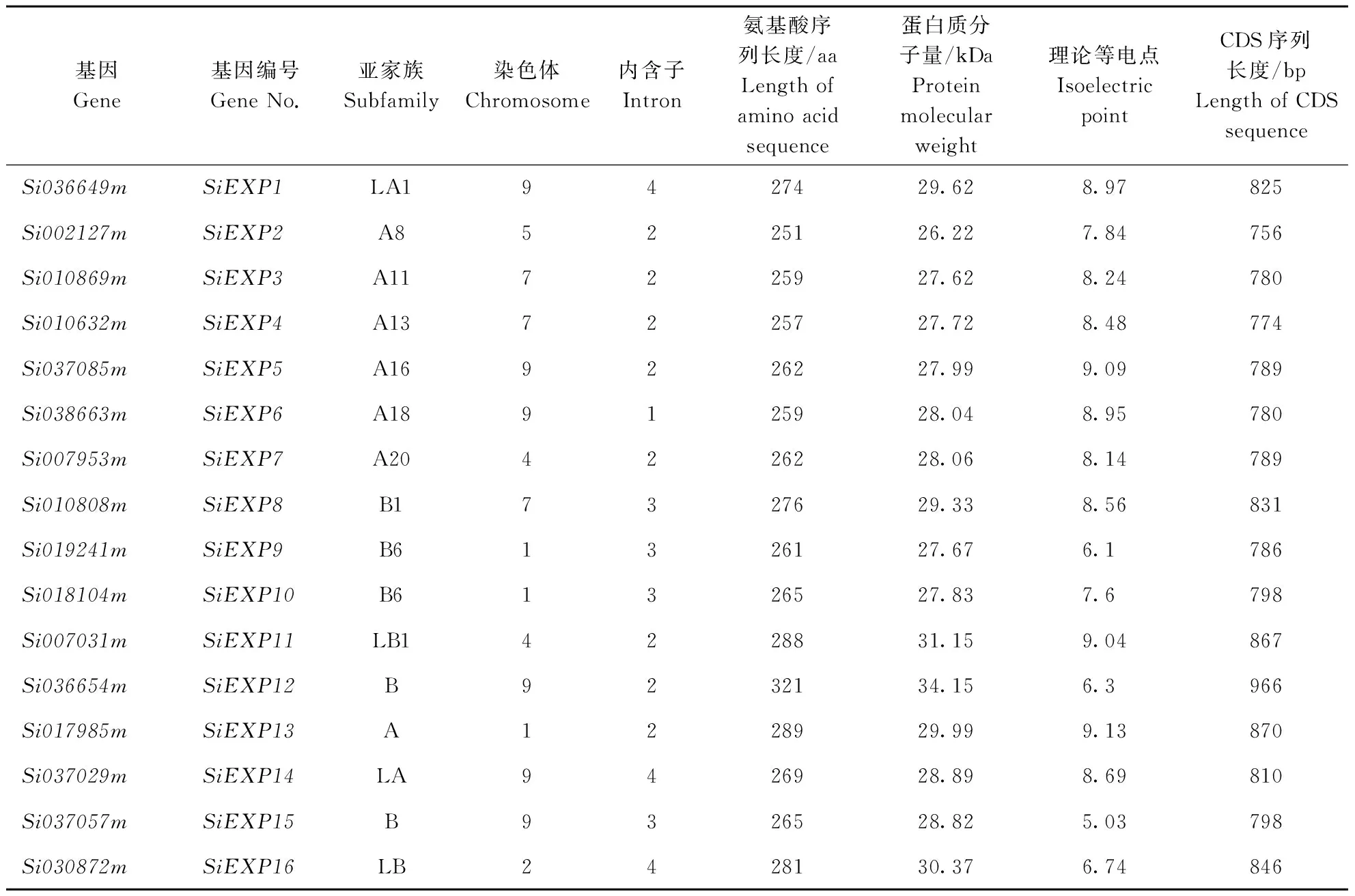
表1 谷子扩展蛋白基本信息及理化性质
由Phytozome V11.0中JBrowse下载的基因位置信息可知,9号染色体上包含6个扩展蛋白基因家族成员,最远相距56.9 Mb,其中Si037029m和Si038663m距离最近,约为343.1 kb。第1和7条染色体上包含3个家族成员,其中1号染色体上的Si019241m和Si018104m基因距离最近,约为2.3 kb。第4条染色体上包含2个家族成员,距离约为6.7 Mb。
谷子扩展蛋白基因家族中不同扩展蛋白基因在内含子数目及大小方面都存在差异。16个扩展蛋白基因中8(50%)个基因含有2个内含子,其中6个属于EXPA亚家族;4(25%)个基因含有3个内含子,都属于EXPB亚家族;3(18.75%)个基因含有4个内含子,其中2个属于EXPLA亚家族(表1)。
2.2谷子扩展蛋白理化性质
根据豫谷1号扩展蛋白基因的cDNA序列预测得到的氨基酸序列,发现谷子扩展蛋白的氨基酸序列长度处于251~321 aa之间,平均长度为271 aa(表1)。分子量处于26.22~34.15 kDa之间,A类扩展蛋白均值达到27.95 kDa,B类扩展蛋白均值达到29.56 kDa,总平均值为28.97 kDa。理论等电点处于5.03~9.13之间,平均值为7.93(表1)。
2.3谷子扩展蛋白基因家族启动子元件
根据豫谷1号全基因组序列,获取谷子扩展蛋白基因CDS区上游3 000 bp的核苷酸序列,用PlantCARE进行启动子在线分析,发现除了基本启动子元件TATA-box和CAAT-box外,谷子扩展蛋白基因还包含多种与胁迫响应相关的顺式作用元件(表2),包括乙烯响应元件ERE、水杨酸响应元件TCA-element、热胁迫响应元件HSE、低温胁迫响应元件LTR、植物激素响应元件TGA-element、赤霉素响应元件P-box、真菌诱导响应元件Box-Wl等。
利用PlantCARE在线分析16个谷子扩展蛋白基因起始密码子上游3 000 bp的顺式作用元件(表3),统计分析发现每个基因至少含有105个调控元件,最少的是Si037057m基因,含有106个顺式作用元件。调控Si037085m基因的顺式作用元件最多,高达165个,其中与病虫害胁迫相关的茉莉酸甲酯响应元件TGACG-motif多达14个,是家族成员中最多的。有15个扩展蛋白基因家族成员包含2~10个脱落酸响应元件ABRE,其中Si037029m基因含10个;Si010632m基因没有脱落酸响应元件ABRE。有14个家族成员包含1~5个干旱响应MYB结合位点MBS,其中Si007953m基因含有5个。16个谷子扩展蛋白基因家族成员中都包含20个以上的光响应调控元件,其中Si038663m基因最多,高达50个。
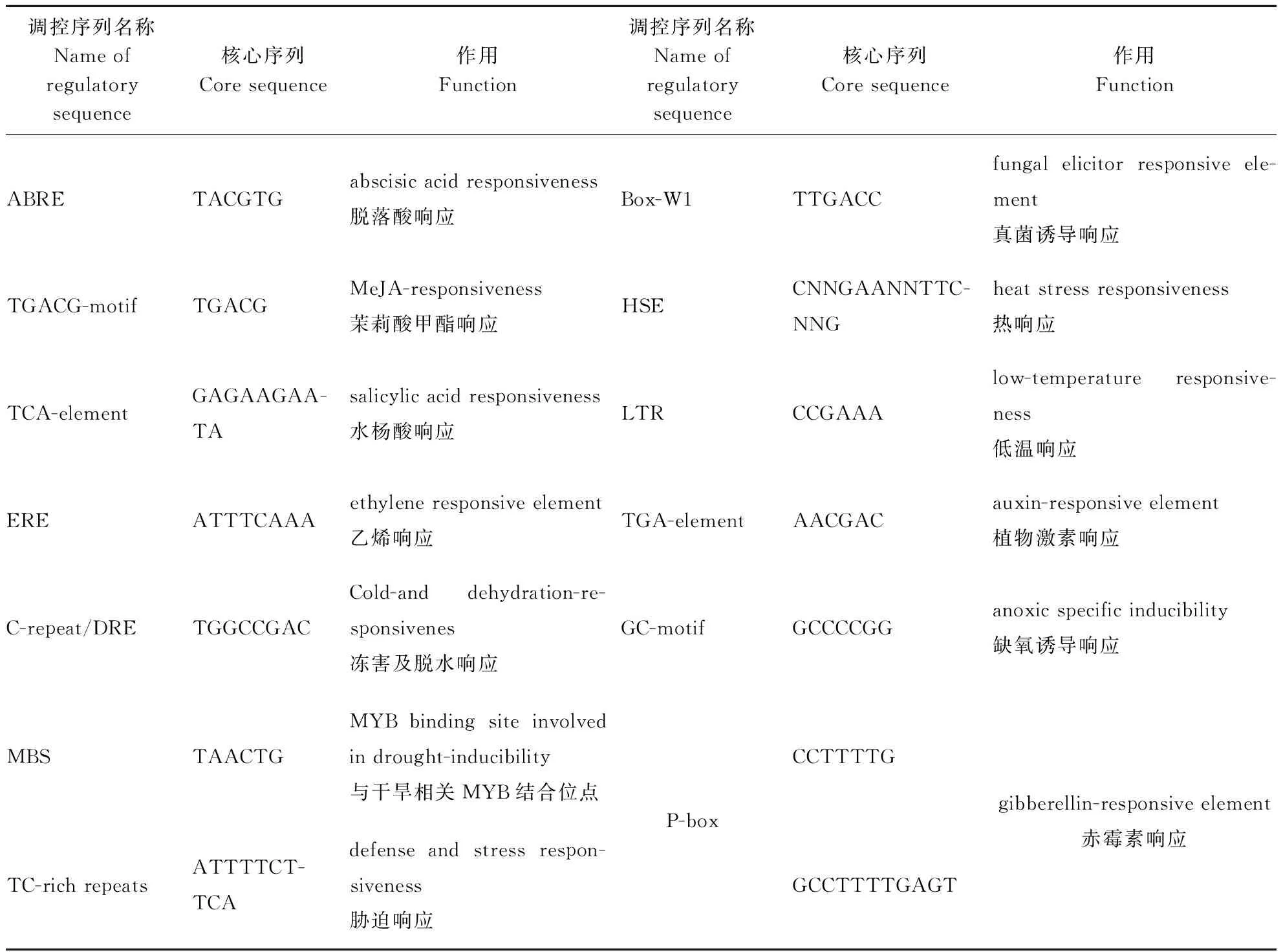
表2 谷子扩展蛋白基因启动子顺式作用元件
2.4谷子扩展蛋白基因在干旱胁迫下的表达模式
用20% PEG-6000模拟干旱胁迫,处理耐干旱品种勾勾母鸡咀(GG)和干旱敏感品种晋汾16(JF16),通过表达谱分析发现,在干旱胁迫下GG转录组中有4个扩展蛋白基因(SiEXP12,SiEXP14,SiEXP15, 和SiEXP16),JF16转录组中有5个扩展蛋白基因(SiEXP12,SiEXP13,SiEXP14,SiEXP15, 和SiEXP16)。
两个品种中SiEXP12基因仅在干旱胁迫下表达,且在JF16中表达水平较GG高(图2-a)。
SiEXP13基因仅在干旱敏感品种JF16中表达,且在干旱胁迫下表达量显著上调(图2-b)。
在正常浇水条件下,SiEXP14基因在JF16中的表达水平较GG高;干旱胁迫处理后,该基因在两个谷子品种中的表达水平均上调,且在JF16中上调幅度较大(图2-c)。
GG中,SiEXP15基因仅在干旱胁迫下表达;JF16中,该基因在干旱胁迫处理后表达水平显著下调(图2-d)。
SiEXP16基因在JF16品种中的表达水平较GG高。干旱胁迫处理后该基因在两个品种中表达水平均下调(图2-e)。

表3 谷子扩展蛋白基因家族启动子顺式元件预测
注:以上数据除去了基本启动子元件TATA-box和CAAT-box。
Note:The data does not include the basic promoter elements such as TATA-box and CAAT-box.
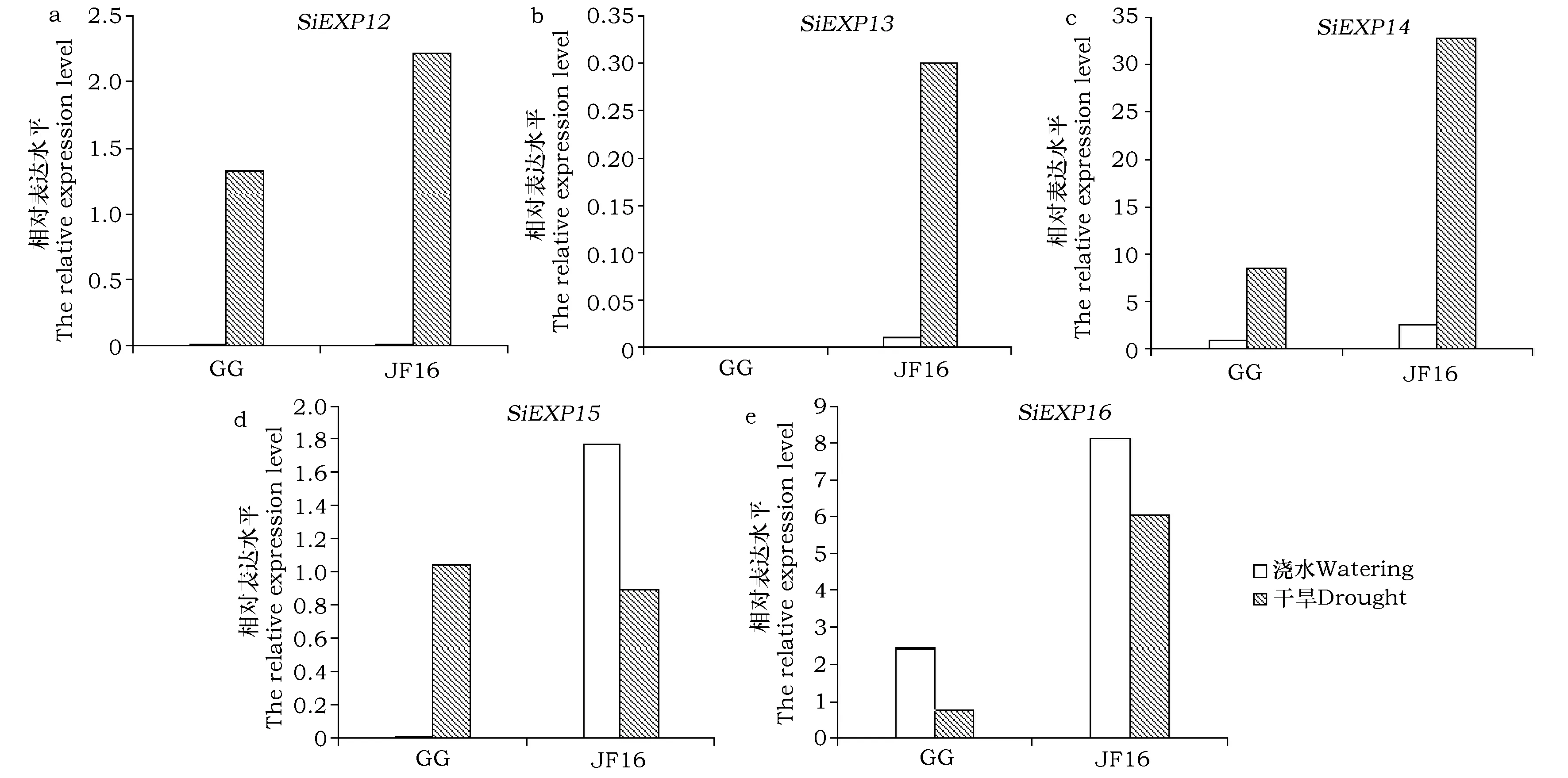
图2 GG和JF16扩展蛋白基因的表达模式Fig.2 Expression profiles of expansin genes in GG and JF16
3 讨论与结论
扩展蛋白是植物细胞壁重要的组成成分。通过对谷子扩展蛋白基因组的分析,有利于我们了解扩展蛋白基因基本特征及功能,对后续研究具有重要意义。
在已测序的单子叶植物水稻基因组中发现扩展蛋白家族成员为56个,其中EXPA和EXPB两种蛋白比例分别为59%和33%,谷子中两类蛋白比例分别为44%和31%,由此可见,这两类蛋白表现出一定的保守性[12]。在内含子数目方面,谷子扩展蛋白基因也表现出保守性,如85%以上的EXPA亚家族含有2个内含子,80%以上的EXPB亚家族含有3个内含子,对于EXPLA和EXPLB两个亚家族基本含有3个或4个内含子。
本研究对谷子扩展蛋白构建了系统发育树,结果表明,4个亚家族的成员会聚集到一起。该结论与李昊阳等对杨树扩展蛋白的研究结果类似[14]。
谷子是重要的粮食作物,然而高温、低温、干旱等不良环境因素会对谷子产量造成极大影响,因此,研究其抗逆机制显得尤为重要。本文研究发现,谷子扩展蛋白家族基因上游约3 000 bp的启动子序列中含有多个与逆境相关的顺式作用元件,包括低温响应元件LTR、热响应元件HSE、真菌诱导响应元件Box-W1、干旱响应MYB结合位点MBS和脱落酸响应元件ABRE,扩展蛋白基因家族可能通过以上顺式作用元件参与谷子对胁迫信号传导途径的应答,在植物响应胁迫过程中发挥作用[10]。
本研究通过分析谷子5个扩展蛋白基因的表达水平,发现在正常浇水条件下,SiEXP12基因在两个谷子品种中均不表达,但在干旱胁迫下表达量显著上调,说明该基因在耐干旱品种和干旱敏感品种中不具有表达特异性;SiEXP13基因仅在JF16中表达,且在干旱胁迫下表达水平显著上调,说明该基因对干旱胁迫有响应,但并未有效缓解植株干旱状况;SiEXP14和SiEXP16基因在两个品种受到干旱胁迫时表达水平分别显著上调和下调,说明这两个基因在耐干旱品种和干旱敏感品种中的作用是相同的,只是表达差异造成了两者抗旱性差异[13];干旱胁迫下,仅有SiEXP15基因在耐干旱品种GG中的表达水平较干旱敏感品种JF16高。
然而,结合亲缘关系图及启动子顺式调控元件的预测结果,并未发现基因亲缘关系与扩展蛋白基因表达间的规律;也未发现胁迫相关的调控元件与扩展蛋白表达之间的关联。我们推测可能有3个原因:第一,同一基因在不同组织中的表达水平不同,而扩展蛋白基因的表达水平可能与各组织细胞生长分裂状态呈正相关[15]。第二,扩展蛋白基因的表达非常复杂,会受其他外界因素的影响,如光,植物激素等的调控。本文研究的SiEXP12、SiEXP13、SiEXP14和SiEXP16等4个基因可能与其他胁迫相关。第三,在谷子的不同生育期,GG和JF16的抗旱性也有差别,如在孕穗期GG抗旱性较JF16强,但在灌浆期JF16抗旱性较GG强[13]。以上这些因素都可能导致干旱胁迫下谷子扩展蛋白基因的表达水平与品种抗旱性无关,且差异较大。由于对扩展蛋白的具体作用机制还不清楚,为解决上述疑惑,仍需要进一步对扩展蛋白在植物抗逆性中的作用机制、对逆境的响应机制等进行深入的研究。
综上所述,本研究通过对谷子扩展蛋白基因进行生物信息学分析,并对其中5个基因家族成员的表达水平进行分析,初步了解了扩展蛋白基因与谷子抗旱的关系,为进一步理解扩展蛋白基因在谷子响应干旱胁迫过程中的作用提供了依据,同时为后续深入研究谷子扩展蛋白基因的功能奠定了理论基础。
[1]He L,Zhang B,Wang X C,et al.Foxtail millet:nutritional and eating quality,and prospects for genetic improvement[J].Front Agr Sci Eng,2015,2(2):124-133.
[2]王永丽,王珏,杜金哲,等.不同时期干旱胁迫对谷子农艺性状的影响[J].华北农学报,2012,27(6):125-129.
[3]牛艳梅,沈文涛,周鹏.Expansin超级家族的进化与命名[J].广东农业科学,2007(8):133-135.
[4]Cosgrove D J.Charactereization of long term extension for isolated cell walls from growing cucumber hypocotyls[J].Planta,1989,177:121-130.
[5]Mcqueen-mason S,Durachko D M,Cosgrove D J.Two endogenous proteins that induce cell wall extension in plant[J].Plant Cell,1992,4(11):1425-1433.
[6]Mcqueen-mason S,Cosgrove D J.Disruption of hydrogen bonding between plant cell wall polymers by proteins that induce wall extension[J]. Proc Natl Acad Sci USA,1994,91(14):6574-6578.
[7]徐筱,黄炳如,徐吉臣.翦股颖AsEXP1基因的抗旱性分析[J].北京林业大学学报,2010,32(5):126-131.
[8]王辉,孙涛,赵会君,等.商陆扩展蛋白基因PaEXP1在逆境胁迫下的表达[J].中国科学院研究生院学报,2010,27(4):448-455.
[9]Han Y,Li A X,Li F,et al.Characterization of a wheat(TriticumaestivumL.) expansin gene,TaEXPB23,involved in the abiotic stress response and phytohormone regulation[J].Plant Physiol Biochem,2012,54:49-58.
[10]赵美荣,李永春,王玮.扩展蛋白与植物抗逆性关系研究进展[J].植物生理学报,2012,48(7):637-642.
[11]Xu P,Cai X T,Wang Y,et al.HDG11 upregulates cell-wall-loosening protein genes to promote root elongation in Arabidopsis[J].J Exp Bot,2014,65(15):4285-4295.
[12]施杨,徐筱,李昊阳,等.水稻扩展蛋白家族的生物信息学分析[J].遗传,2014,36(8):809-820.
[13]张雁明.不同谷子品种抗旱性比较及干旱相关基因表达分析[D].太谷:山西农业大学,2014.
[14]李昊阳,施杨,丁亚娜,等.杨树扩展蛋白基因家族的生物信息学分析[J].北京林业大学学报,2014,36(2):59-67.
[15]荆赞革,柳李旺,龚义勤,等.萝卜扩展蛋白基因RsEXPB1克隆与表达特征分析[J].分子植物育种,2009,7(4):801-805.
(编辑:邢国芳)
收稿日期:2016-07-24
作者简介:刘德磊(1982-),男(汉),山东菏泽人,讲师,博士,研究方向:智能计算与模式识别
*通讯作者:李富忠,教授,博士生导师。Tel:13734008985;E-mail:sxaulfz@126.com
基金项目:山西农业大学博士科研启动基金(2014YJ20)
The relationship between expansin gene family and drought stress in foxtail millet
Su Yanbing1, Ma Fangfang1, Wen Siyu1, Liu Rui1, He Lu1, Liu Longlong3, Li Hongying1, Han Yuanhuai1,2,3*
(1.CollegeofAgriculture,ShanxiAgriculturalUniversity,Taigu030801,China, 2.KeyLabaratoryofCropGeneResourcesandGermplamEnhancementonLoessPlateau,MinistryofAgriculture,Taiyuan030031,China, 3.ShanxiKeyLaboratoryofGeneticResourcesandGeneticImprovementofMinorCrops,Taigu030801,China)
[Objective]As an important part of plant cell wall, expansin exists widely in almost every plant genome. Expansin can increase the flexibility of the cell wall and plays an important role in physiological process of plant growth and stress. In order to explore drought genetic resources in foxtail millet. [Methods]16 expansin genes of foxtail millet were analyzed. [Results]They were classified into 4 subfamilies (EXPA, EXPB, EXPLA and EXPLB), through the promoter analysis, we found a number of responsive elements related to drought stress, including drought response MYB binding site MBS, ABA response element ABRE, and so on. The expression levels of expansin genes in two foxtail millet varieties GG (drought-resistant variety) and JF16 (drought-sensitive variety) were analyzed, the results showed that their expression levels were different significantly in different varieties under drought stress. [Conclusion]According tocis-acting elements of promoter and phylogenetic tree of expansin genes, we found out there are no significant relationship between drought stress and these results. But these results provide a foundation for further study on the relationship between expansin genes and stress resistance in foxtail millet.
Foxtail millet, Expansin, Drought stress, Drought tolerance
2016-04-18
2016-05-16
苏彦冰(1993-),女(汉),山西临汾人,硕士研究生,研究方向:谷子抗旱机理
韩渊怀,教授,博士生导师。Tel:0354-6287239;E-mail:swgctd@163.com
国家自然科学基金(31371693,31471556);教育部高等学校博士学科点专项科研基金(20131403110001);山西农业大学科技创新基金(2014022,2014YZ2-5)
S515; Q94
A
1671-8151(2016)10-0685-06
猜你喜欢
当代陕西(2021年21期)2022-01-19
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
生物学通报(2018年12期)2018-10-10
浙江农业学报(2017年1期)2017-05-17
西南农业学报(2016年4期)2016-05-17
现代农业(2015年5期)2015-02-28
草食家畜(2012年2期)2012-03-20
现代农业科技(2009年17期)2009-03-08
