
小型浅水湖泊表层沉积物氨氧化古菌群落结构特征
2016-10-20 05:25燕文明赵倩维杨艳青
河海大学学报(自然科学版) 2016年5期
燕文明,刘 凌,赵倩维,杨艳青
(1. 河海大学水文水资源与水利工程科学国家重点试验室,江苏 南京 210098;2. 淮阴工学院交通工程学院,江苏 淮安 223300)
小型浅水湖泊表层沉积物氨氧化古菌群落结构特征
燕文明1,刘凌1,赵倩维2,杨艳青1
(1. 河海大学水文水资源与水利工程科学国家重点试验室,江苏 南京210098;2. 淮阴工学院交通工程学院,江苏 淮安223300)
为更深入地了解氨氧化古菌(AOA)在浅水湖泊表层沉积物中的分布特征,以淮河流域下游里下河地区浅水湖泊为研究对象,采用分子生物学和微生物学方法研究了该区域4个不同功能类型的浅水湖泊中的AOA群落结构。结果表明,里下河地区的浅水湖泊中存在AOA,为AOA的全球地理分布生境提供了重要补充。大纵湖湖心处表层沉积物中AOA的多样性和丰富度最高,AOA主要聚集于Group 1.1b和Group 1.1a,其中71.3%的序列属于Group 1.1b,28.7%的序列属于Group 1.1a。沉水植物的存在明显增加了Group 1.1b的丰富度,丰富的有机质有利于促进AOA的生长。得胜湖岸周边的工厂、采沙场、渔业养殖以及九龙口上游家畜类养殖影响了表层沉积物中的群落特征。
表层沉积物;氨氧化古菌;群落结构;污染物来源;多样性指数;丰富度指数;浅水湖泊;里下河地区
2008年,Herrmann等[1]在淡水湖泊水生植物根际中发现了AOA,随后人们开始了对AOA的广泛研究,研究还发现具有氨氧化能力的氨氧化古菌(AOA)有amoA[2]。Prosser等[3]将AOA分为3类:海洋、大多数水体以及沉积物中的海洋环境硝化古菌群落(Group 1.1a),土壤及其他陆源环境中的土壤环境硝化古菌群落(Group 1.1b),嗜高温环境硝化古菌群落(Group 1.1c)。Pester等[4]将AOA序列划分为5个类群:Nitrosocaldus类群、Nitrosopumilus类群、Nitrosotalea类群、Nitrososphaerasister类群、Nitrososphaera类群。Lliros等[5]指出在溶解氧含量较高的环境中,系统中的AOA多聚类于Group 1.1b和Group 1.1a。Wu 等[6]对太湖底泥的研究得出,该区域内的AOA多属于Group 1.1b,有少量属于Group 1.1a。浅水湖泊表层沉积物物理化学特性存在一定的差异,导致了其中的细菌群落结构不同[7-8],AOA的数量、多样性及其生存环境范围等多于AOB[9]。AOA的多样性受到外界环境因素的制约,土壤类型[10]、施肥处理[11]、铵浓度[12]等的变化都可能会引起AOA的丰富度和分布的改变。AOA在氮循环中的作用和功能都已得到深入研究,而关于同一地区不同污染来源的沉积物AOA丰富度和群落分布的研究较少。笔者以里下河地区小型浅水湖泊表层沉积物为研究对象,应用微生物技术研究不同类型浅水湖泊表层沉积物中AOA的多样性及丰富度其群落分布特征,旨在更好地认识AOA的群落结构组成,为进一步研究AOA在浅水湖泊氮循环中的作用奠定基础。
1 试验材料与方法
1.1采样点的布设与概况
在淮河下游里下河地区的得胜湖、九龙口、大纵湖布设4个采样点,编号分别为DS、JLK、DZ、DZ5,如图1所示。每个采样点同时采集3个表层沉积物样品混合。各采样点受人类扰动作用和生物作用强度不同,沉水植物密度、围网养殖程度、工业废水以及面源污染的注入对湖泊的环境影响不同。DS位于得胜湖中心,水深约3.5 m,得胜湖为河道型湖泊,受岸边化肥厂和采沙场影响较大,湖面多被围隔养殖。JLK位于九龙口入湖口,刚经过疏浚,水深1.5 m,上游存在渔业养殖和家畜类养殖现象。DZ位于大纵湖北部,水深约2.1 m,围网养殖现象严重。DZ5位于大纵湖湖心,水深约1 m,水体清澈,菹草和金鱼藻等水生植物密布。
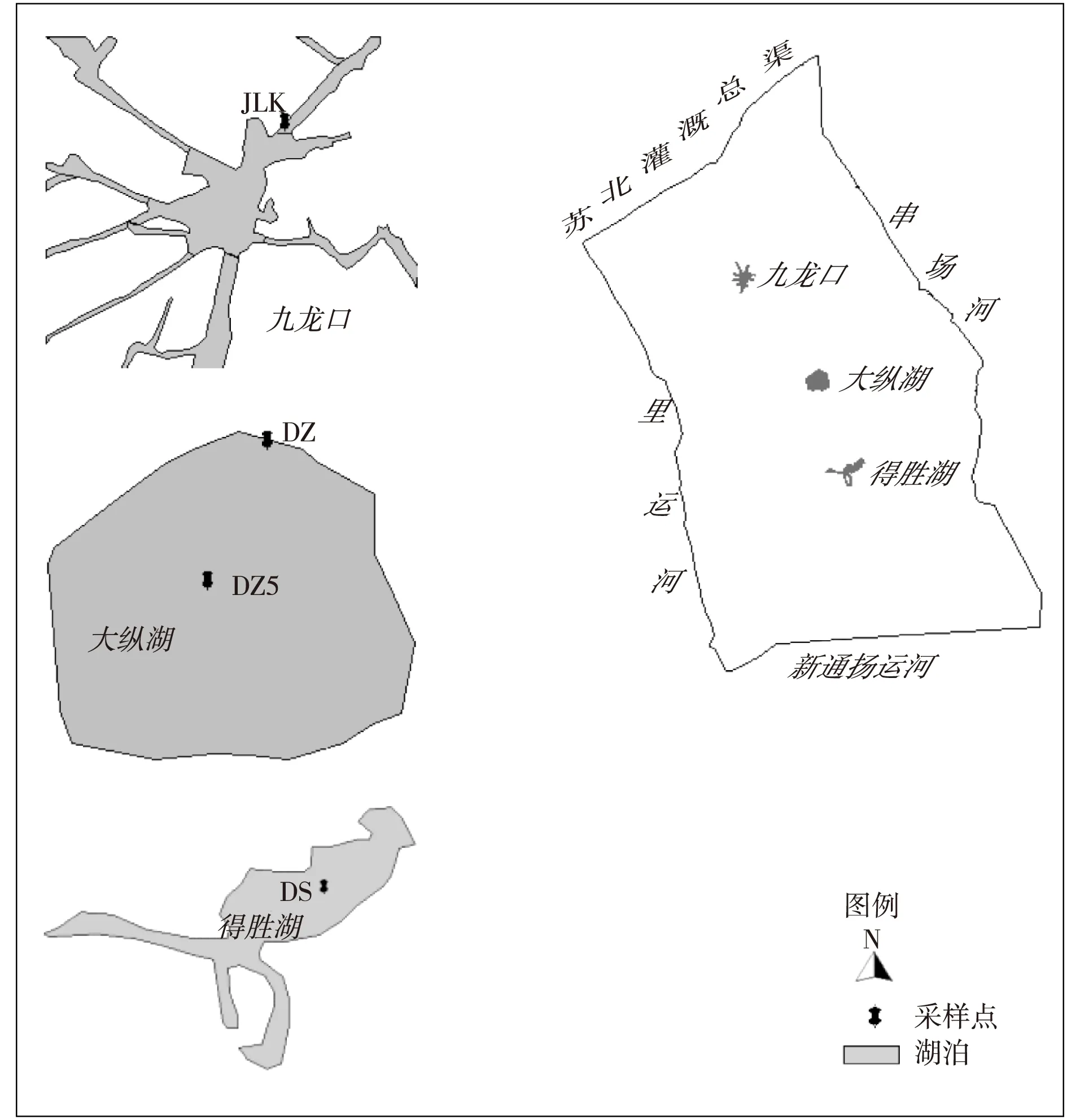
图1 研究区典型型湖泊位置与采样点位置Fig. 1 Locations of typical lakes and sampling points in study area
1.2沉积物DNA的提取和PCR的扩增
称取冻干过筛后的沉积物样品0.5 g,提取和纯化后,用预冷的70%乙醇洗涤沉淀2次,重悬于灭菌的TE溶液中,终体积为50 μL,-20℃保存。每个沉积物样品重复提取3次然后合并。将总DNA 进行10倍稀释后作为模板。AOA的amoA基因采用的特异性引物为amoAF(5′-STAATGGTCTGGCTTAGACG-3′) 和amoAR(5′-GCGGCCATCCATCTGTATGT-3′)[1],以提取纯化后的DNA作为模板,进行25 μL体系扩增。
1.3克隆文库构建
(a)纯化。使用PromegaAgaroseGelDNA 纯化试剂盒对PCR扩增产物进行切胶纯化。(b)连接与转化。将PCR 回收产物与pGEMTeasy载体进行连接后转入JM109 感受态细胞。(c)筛选。进行蓝白筛选。(d)挑菌。AOA选取40个白色克隆。(e)验证。将通用引物T7、SP6作为载体,PCR 扩增鉴定阳性克隆。(f)测序。挑取代表菌株进行测序(3 500,ABI,USA)。(g)比对。使用DOTUR软件获得分类操作单元(Operational Taxonomic Units,OTUs)后,每个OTU类型选取一个代表序列在GenBank上进行BLASTn相似性比对,挑选相似性最高的序列为参比序列。获得的AOA核酸序列登录号为:KM516351-KM516472。
1.4系统发育分析
应用DOTUR软件对每个文库单独计算操作分类单元(OTUs),选取每个OTU中最具代表性的序列,通过BLAST搜索GenBank中最相似序列进行比对,并参照有关文献,选取参照序列,再用ClustalX 1.8进行多序列比对。应用Jukes-Cantor参数模型计算进化距离[13],运用MEGA5.0软件的Kimura 2-parameter 校正模型检验不同的进化距离,并进行两两比对,通过Kimura 2邻接法构建系统发育树[14],进化树的可靠性采用自举法进行评估,自举值设为1 000次。通过DOTUR软件计算Shannon-Weiner指数、Simpson多样性指数、SACE丰富度指数和SChao1丰富度指数等。克隆库的覆盖率由公式C=[1-(n1/N)]×100%计算,其中n1是OTUs的数目,N为总克隆数[15]。
2 结果与讨论
2.1水环境特征
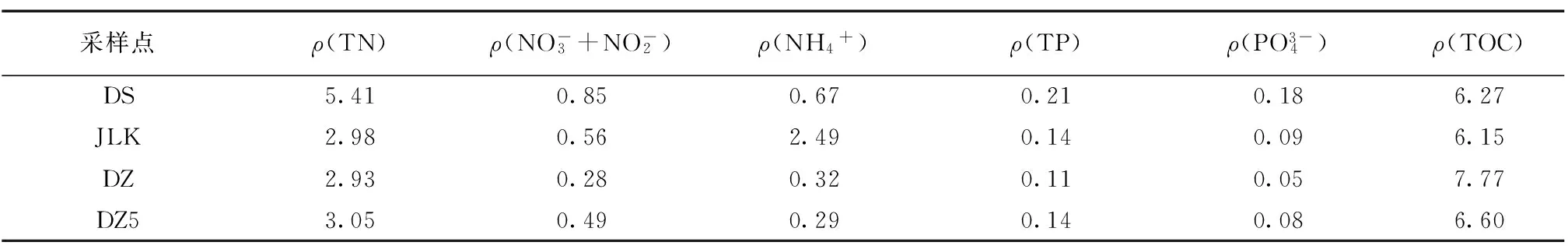
表1 水中营养盐和有机质含量Table 1 Concentration of nutrients and organic matter in water mg/L
2.2氨氧化古菌amoA基因的多样性分析
研究中所选取的里下河地区浅水湖泊表层沉积物中微生物总 DNA 提取较完整,进行PCR扩增,经胶回收纯化和克隆测序分析,序列长度为629 bp。BLAST比对发现PCR产物序列与其他不可培养AOA的amoA基因同源性达到99%,证明了里下河地区得胜湖、九龙口和大纵湖表层沉积物中存在AOA。在对AOA进行扩增后,各40个克隆进行测序,得到目的序列克隆数分别为27(DS)、40(DZ)、39(DZ5)和16(JLK)。由于JLK表层沉积物的质粒较为复杂,得到的目的序列较少,只有16个,无法使用DOTUR软件计算其多样性指数。本文仅对DS、DZ和DZ5中的AOA按99%的同源性划分为8个、9个和14个OTUs,并计算每一个OTU的SACE丰富度指数、SChao1丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数。研究中DS、DZ和DZ5中的AOA克隆文库覆盖率较高(表2),均大于65%,表明文库包括了大多数AOA的amoA基因类型。通过表2可知DZ5中AOA的Shannon-Wiener多样性指数、Simpson多样性指数最大。DZ的Shannon-Wiener多样性指数和Simpson多样性指数最小。比较分析可知,DZ5的AOA多样性和丰富度最高,而DZ和DS多样性和丰富度较低。
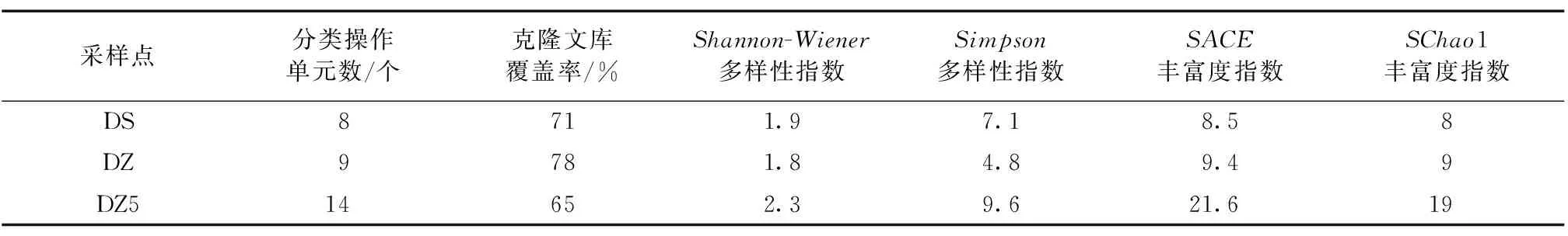
表2 氨氧化古菌amoA的多样性Table 2 Diversity of ammonia-oxidizing archaea amoA
2.3amoA基因的系统发育分析
按99%同源性将4个采样点的AOA划分为16个OTUs,BLAST寻找有相似来源的参比序列,用Mega5软件构建系统发育树。研究中的AOA主要聚集于Group 1.1b和Group 1.1a。在122个目的序列中,有87个属于Group 1.1b,占71.3%。这87个序列里有25个是从DS中分离得到的,有4个是从JLK中分离得到的,从DZ和DZ5中分离得到的序列分别为21个和37个。有35个序列属于Group 1.1a,占28.7%。DS、DZ、DZ5和JLK中AOA属于Group 1.1a的目的序列分别有2个、19个、2个和12个。可见,DS和DZ5中的AOA以Group 1.1b为主,DZ中AOA的Group 1.1a和Group 1.1b所占比例相差不大,JLK以Group 1.1a为主。Group 1.1b的87个序列中有85个被聚类为111clone5D11GenBank(EU671952)和222cloneCFSAOA-24GenBank(GQ906649)这2个亚类。其中,被聚类为前者的序列有13个,来自于DS、DZ和DZ5的分别有5个、2个和6个;被聚类为后者的序列有72个,来自于DS、DZ、DZ5和JLK的分别有20个、19个、30个和3个。16个OTUs中,只有属于Nitrososphaera类群的OTU1 在所有样品中都存在,占21.3%,数量最多,与在长江口底泥样品发现的AOA序列高度相似。Group 1.1b的87个序列被聚为11个OTUs,参考序列有来自废水处理系统的KJ496929,洞庭湖沉积物KC108808,河流底泥KF857068,长江口底泥KC735552、KC735379和KC735383,京杭大运河底泥KF537016,巢湖出口处的底泥JQ698526.1,红树林土壤KF169744。Nitrosopumilus类群的35个序列被分为5个OTUs,参考序列分别是从热带河口沉积物中分离得到的FJ601570,高山湖泊的KF898586和KC735494,长江口的KJ005026和KF413247。16个OTU的代表序列与各自参比序列的基因片段在核苷酸水平上的同源性均为99%,有高相似性。
里下河地区AOA的amoA基因序列与来源于土壤的Nitrososphaeraviennensis(FR773159)和UnculturedcrenarchaeoteForsmid54d9(AJ627422)的相似性良好,达到了97%以上,与热泉中富集的NitrososphaeragargensiscloneRHGCaA18c(EU281319)的相似性良好[6],达到了93%;与从西雅图海洋水族馆中成功分离的海洋AOA菌株NitrosopumilusmaritimusSCM1的相似性为71%[16],相似性较差。在122个目的序列中,有71.3%序列属于Group 1.1b,28.7%的序列属于Group 1.1a。Schleper[17]指出海洋、大多数水体以及沉积物来源的AOA聚类于Group 1.1a;土壤及其他陆源环境来源的AOA聚类于Group 1.1b。可见,里下河地区的AOA以土壤和其他陆源环境来源为主。4个采样点中,DS的AOA以Group 1.1b为主,占92.6%,结合得胜湖是河道型湖泊,受周边环境影响较大,可知该采样点AOA以土壤及其他陆源环境来源为主;而位于大纵湖湖心的DZ5的Group 1.1b也较多,占94.9%,由于该点位于湖心,其氨AOA来源于陆源环境的可能较小,Group 1.1b类群丰度较大可能与该点丰富的沉水植物有关。
通过系统发育分析可知, DZ5中AOA的多样性和丰富度最高,而DZ和DS较低。DZ5水生植被较为旺盛,死亡后腐烂沉积于湖底,沉积物中TN很高,是DS、DZ和JLK的2倍以上,结合DZ5的实际情况,沉水植物明显增加了Group 1.1b的丰富度。沉水植物的存在也导致了DZ5中AOA的多样性和丰富度明显高于DS和DZ。可见,表层沉积物中丰富的有机质含量有利于促进AOA的生长。也有研究指出沉积物中的有机质含量较高可能会对AOA起到抑制作用[16],这一点与本文的结论相反。DZ的Group 1.1b和Group 1.1a差别较小,说明该采样点AOA的来源以陆源环境以及自身的水体和沉积物为主。
在对DS和JLK所挑取的 40个代表菌株进行测序后,发现测序结果分别只有27个和16个是AOA的序列,其他序列均能找到AOA基因的扩增引物,但序列比对结果是质粒序列,这可能是样品的质粒上有引物结合位点所致。DS和JLK所处的环境与DZ和DZ5差异较大。DS采自河道型湖泊,受岸边的工厂、采沙场以及湖面围网养殖的影响较大。JLK位于九龙口南部入湖口,所在的湖区刚经过底泥疏浚,上游区域主要有渔业养殖和家畜类养殖。这些都有可能会对表层沉积物中的群落结构分布造成较大影响。所以需要寻找并添加新的特异引物对用于克隆前的PCR扩增,使得克隆文库具有足够的覆盖率,进而更为准确地反应沉积环境中AOA的群落结构特征。而DZ位于大纵湖的出湖口,DZ5位于湖心,这2个采用点的表层沉积物中AOA的群落分布结构特征与一般的浅水湖泊或者河口有一定的相似性,用通用的特异性引物对基本可以使得克隆序列具有良好的覆盖率。因此,要想更为细致准确地研究DS和JLK中的AOA,还需要寻找合适的特异性引物以满足构建克隆文库需要。
3 结 论
a. 淮河流域下游的里下河地区浅水湖泊表层沉积物中存在AOA,为AOA的全球地理分布生境的提供了重要补充。
b. 大纵湖、九龙口和得胜湖的AOA主要聚集于Group 1.1b和Group 1.1a,有71.3%的序列属于Group 1.1b。
c. 大纵湖湖心表层沉积物中的AOA多样性和丰富度最高。沉水植物的存在明显增加了Group 1.1b的丰富度,丰富的有机质有利于促进AOA的生长。
[1] HERRMANN M, SAUNDERS A M, SCHRAMM A. Archaea dominate the ammonia-oxidizing community in the rhizosphere of the freshwater macrophyte Littorella uniflora[J]. Applied and Environmental Microbiology, 2008, 74(10): 3279-3283.
[2] FRANCIS C A, ROBERTS K J, BEMAN J M, et al. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean [J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(41): 14683-14688.
[3] PROSSER J I, NICOL G W. Relative contributions of archaea and bacteria to aerobic ammonia oxidation in the environment [J]. Environmental Microbiology, 2008, 10(11): 2931-2941.
[4] PESTER M, RATTEI T, FLECHL S, et al. AmoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions [J]. Environmental Microbiology, 2012, 14(2):525-539.
[5] LLIROS M, GICH F, PLASENCIA A, et al. Vertical distribution of ammonia-oxidizing crenarchaeota and methanogens in the epipelagic waters of Lake Kivu (Rwanda-Democratic Republic of the Congo)[J]. Applied and Environmental Microbiology, 2010, 76(20):6853-6863.
[6] WU Yucheng, XIANG Yan, WANG Jianjun, et al. Heterogeneity of archaeal and bacterial ammonia-oxidizing communities in Lake Taihu, China[J]. Environmental Microbiology Reports, 2010, 2(4):569-576.
[7] 侯俊, 王超, 王沛芳, 等. 太湖表层沉积物粒度组成时空分布特征及分类命名[J]. 河海大学学报(自然科学版), 2013, 41(2):114-119. (HOU Jun, WANG Chao, WANG Peifang, et al. Temporal variability and spatial distribution of granulometric composition of surface sediments and classification in Taihu Lake[J]. Journal of Hohai University (Natural Sciences), 2013, 41(2):114-119. (in Chinese))
[8] 赵大勇,孙一萌,方超,等. 太湖梅梁湾不同深度沉积物中细菌群落结构组成[J]. 河海大学学报(自然科学版), 2013, 41(4): 283-287.(ZHAO Dayong, SUN Yimeng, FANG Chao, et al. Vertical distribution of bacterial community composition in sediments of Meiliang Bay in Taihu Lake[J]. Journal of Hohai University (Natural Sciences), 2013, 41(4): 283-287. (in Chinese))
[9] CHEN Xueping, ZHU Yongguan, XIA Yue, et al. Ammonia-oxidizing archaea: important players in paddy rhizosphere soil [J]. Environmental Microbiology, 2008, 10 (8):1978-1987.
[10] 张秋芳, 徐继荣, 苏建强, 等. 河口生态系统氨氧化菌生态学研究进展[J]. 生态学报, 2012(18): 5878-5888. (ZHANG Qiufang, XU Jirong, SU Jianqiang, et al. Research progress on ammonia-oxidizing microorganisms in estuarine ecosystem[J]. Acta Ecologica Sinica, 2012(18): 5878-5888.(in Chinese))
[11] LEININGER S, URICH T, SCHLOTER M, et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils [J]. Nature, 2006, 442:806-809.
[12] YANG J B, LI X C, XU L, et al. Influence of the nitrification inhibitor DMPP on the community composition of ammonia-oxidizing bacteria at microsites with increasing distance from the fertilizer zone [J]. Biology and Fertility of Soils, 2013, 49 (1):23-30.
[13] LI X R, XIAO Y P, REN W W, et al. Abundance and composition of ammonia-oxidizing bacteria and archaea in different types of soil in the Yangtze River estuary [J]. Journal of Zhejiang University-Science B, 2012, 13 (10):769-782.
[14] KUHNER M K, FELSENSTRIN J. A simulation comparison of phylogeny algorithms under equal and unequal evolutionary rates [J]. Molecular biology and evolution, 1994, 11 (3):459-468.
[15] TAMURA K, PETERSON D, PETERSON N, et al. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods [J]. Molecular Biology and Evolution, 2011, 28 (10):2731-2739.
[16] ZENG J, BIAN Y Q, XING P, et al. Macrophyte species drive the variation of bacterioplankton community composition in a shallow freshwater lake [J]. Applied and Environmental Microbiology, 2012, 78(1):177-184.
[17] SCHLEPER C, JURGENS G, JONUSCHEIT M. Genomic studies of uncultivated archaea [J]. Nature Reviews Microbiology, 2005, 3 (6):479-488.
Characteristics of community structure of AOA in surface sediments of small shallow lakes
YAN Wenming1, LIU Ling1, ZHAO Qianwei2, YANG Yanqing1
(1.StateKeyLaboratoryofHydrology-WaterResourcesandHydraulicEngineering,Nanjing210098,China;2.FacultyofTransportationEngineering,HuaiyinInstituteofTechnology,Huaian223300,China)
To investigate the distribution characteristics of ammonia-oxidizing archaea (AOA) in sediments of shallow lakes, we chose the shallow lakes in the Lixia River area, which is in the lower Huaihe River Basin, as the research object and studied the community structure of AOA in four shallow lakes with different functional types in the region using the methods of molecular biology and microbiology. The results show that there exists AOA in this region and this provides an important supplement to the global geographic distribution and habitat of AOA. In the surface sediments of this region, the diversity and richness of AOA is highest in the center of Dazong Lake. AOA is mainly accumulated in the groups 1.1b and 1.1a, in which, 71.3% of the sequences belong to Group 1.1b and 28.7% of the sequences belong to Group 1.1a. The presence of submerged plants significantly improves the richness of Group 1.1b, and abundant organic matter is conducive to promoting the growth of AOA. The surrounding factories, sandpits, fisheries, and the livestock breeding upstream of Jiulongkou affect the characteristics of the community in the surface sediments.
surface sediments; AOA; community structure; pollution sources; diversity index; richness index; shallow lake; Lixia River area
10.3876/j.issn.1000-1980.2016.05.012
2015-04-03
国家自然科学基金(41301531,51279060)
燕文明(1982—),女,江苏沛县人,高级实验师,博士,主要从事环境水文及水环境保护研究。E-mail:ywm0815@163.com
X524
A
1000-1980(2016)05-0449-05
猜你喜欢
舰船科学技术(2022年20期)2022-11-28
土壤学报(2022年3期)2022-08-26
蜜蜂杂志(2022年5期)2022-07-20
大自然探索(2022年5期)2022-07-11
知识就是力量(2022年6期)2022-06-16
蜜蜂杂志(2022年3期)2022-05-23
蜜蜂杂志(2022年2期)2022-04-15
蜜蜂杂志(2021年11期)2021-02-18
农产品市场周刊(2018年15期)2018-05-16
火控雷达技术(2016年1期)2016-02-06
