
抗菌肽高效表达及生产优化研究进展
2016-10-13 13:20杨平袁奕豪杨晓莉钟玥廖于鑫高荣
生物技术通报 2016年3期
杨平袁奕豪杨晓莉钟玥廖于鑫高荣
(1. 四川大学生命科学学院,成都 610064;2. 四川大学高分子科学与工程学院,成都 610064)
抗菌肽高效表达及生产优化研究进展
杨平1袁奕豪2杨晓莉1钟玥1廖于鑫2高荣1
(1. 四川大学生命科学学院,成都 610064;2. 四川大学高分子科学与工程学院,成都 610064)
抗菌肽是基因编码、经外界诱导产生的一类多肽。在抗生素引发“耐药性”难题的时代,抗菌肽以广谱杀菌、不易产生耐药性、抗肿瘤、抗病毒等特点而拥有超越抗生素、造福人类的巨大潜能。然而,天然抗菌肽还需进行增强抗菌性和稳定性、降低细胞毒性等改造,并通过工业化改良实现大规模生产。因此现从抗菌肽改造设计、生产应用等最新研究做较为详细的综述。
抗菌肽;分子设计;表达策略;生产优化
抗菌肽因其拥有的广谱抗菌、不易产生耐药性、调节先天免疫等诸多优点而备受关注。目前市场上已有抗菌肽产品,并且市场效果良好。但是天然的抗菌肽仍然存在许多不足,如天然含量少、生产成本高、毒副作用大和工艺条件复杂等。因此其作为具潜力的生物制品之一,自发现以来被不断改造以改善抑菌活性、表达产量及溶血性等特性[1-4]。抗菌肽的优化改进一直是研发阶段的主要任务,也是决定整个生产的重要因素。而迄今为止,研究者对抗菌肽生产及应用过程中的改进也做了相应探讨。
1 研发设计
研发阶段可从根本上解决抗菌肽产量不高、溶血性和抗原性等问题,提高广谱抑菌性和表达产量。通过不断研发,人们在抗菌肽分子的设计、表达载体和表达系统的优化与选择、计算机模拟预测等方面取得了一些可喜成果。现逐一阐述如下。
1.1 抗菌肽分子设计
1.1.1 氨基酸组成与序列改变 抗菌肽的抑菌、抗癌及增强天然免疫等优点主要因为它的螺旋结构、两亲性、阳离子等特点。目前许多分子设计都是针对其氨基酸组成与序列进行改造。这类改造主要有两种,一是不改变氨基酸的种类而改变其顺序;二是对氨基酸的种类进行增减从而赋予其新性能。突变可导致氨基酸残基的移码、替换、插入或者删减等,是抗菌肽结构功能研究中的常用方法。抗菌肽经过改造修饰后能有显著优势变化,如抗菌活性提高、溶血性降低、抗癌能力增强、高表达量等。明飞平等[5]通过PCR敲除抗菌肽PR39的5'UTR多余核苷酸得到重组质粒,再构建已删除8个核苷酸的新型表达载体pPIC9K-PR39-D-E,产生的抗菌肽具有高表达量和抑菌活性。刘忠渊等[6]将cecropin-XJ的C末端赖氨酸突变成天冬酰胺,抑菌试验检测发现后者抑菌活性高很多。它们的电荷数相同、螺旋度与疏水性相似,在此情况下二者的抑菌活力不同,推测应该是由于天冬酰胺导致的结果。
1.1.2 去除多余结构并保留修饰活性部分 天然抗菌肽中不都是保证其活性的必需成分,而较大的肽链和蛋白质分子会对抗菌肽的稳定性、免疫原性等产生影响。因此,去除其多余结构、保持活性部分是重要改良措施之一。van Dijk等[7]将cathelicidin-2截短后,溶血性和毒性均得到明显改善。Jittikoon等[8]对Cc-CATH3的氨基酸残基进行研究发现,其N端的前4个氨基酸残基对执行抗菌肽的功能没有任何影响,而第5-8个残基则对抗菌肽活性起到至关重要的作用。
1.1.3 抗菌肽形态设计 不同形状抗菌肽的抑菌效果、稳定性等不同。于岚岚等[9]合成线性、环状、短链多肽发现,其与细菌细胞膜结合能力及杀菌活性顺序均为长链肽>环状肽>短链肽。稳定性是环状肽>长链肽>短链肽。而稳定性往往决定其大规模生产的可能性,因此环状肽对提高抗菌肽产品稳定性具有重要作用。超短肽也具有其特性,大部分抗菌肽可与细胞膜相互作用,而超短肽易将疏水性和正电性共同体现出来,所以抗菌抗病毒能力很强。另外,超短肽分子量较小,所以较易表达。Ahn等[10]用修饰后的组氨酸合成的超短拟肽具有较高抗菌活性,且对蛋白酶及盐均有较高稳定性。
1.1.4 抗菌肽的协同作用 抗菌肽具有杀菌抗病毒的强大功能,而这不仅和其自身的性能机制有关,也受其他物质的影响、调控,这些因素包括多种抗菌肽基因共表达、抗菌肽与抗生素共作用、抗菌肽基因与其他基因相互作用等。天蚕素抗菌肽B在大肠杆菌中的表达是高度有害的,但是Yu等[11]将一种天蚕素抗菌肽B突变体和绿色荧光蛋白融合在大肠杆菌中表达并制备其抗血清,最后得到的融合肽特异性抗血清效价大于1∶25 600。Wang等[12]通过实验探究发现抗菌肽PMAP-36和PRW4与抗生素gentamicin之间对大肠杆菌和金黄色葡萄球菌均存在协同作用,这对临床药物的开发都有着重要意义。
1.2 表达系统的优化与选择
常用的表达系统分为3类:原核表达系统、真核表达系统、杆状病毒表达系统。这3种表达系统各有特点,因此在表达不同抗菌肽时可根据抗菌肽性质与特点选择适当表达系统并对其改造修饰。
1.2.1 原核表达系统 此类常用的是大肠杆菌表达系统,其具有培养简单、繁殖迅速等特点,易进行工业化生产。许多真核基因如胰岛素基因都可在大肠杆菌中实现高效表达,但其仍有不具备真核生物蛋白加工系统、表达量有限等缺陷。对此,不少针对包涵体、杂合肽等内容的研究工作正在展开以使大肠杆菌表达系统不断优化。Wu等[13]研究的杂合蜂毒肽(1-13)-LL37(17-30)(M-L)是将蜂毒肽疏水性N末端(M)与抗菌肽LL37的核心片段(L)相连接,导入E.coli BL21中,在IPTG诱导下获得约165 mg高表达量的融合蛋白。随着DNA重组技术的发展,尤其这一发现——金黄色葡萄球菌带有抗性标志的质粒可作为枯草芽孢杆菌载体,为克服枯草芽孢杆菌只有隐秘性质粒的困难奠定了重要基础,使得枯草芽孢杆菌基因工程的研究迅速发展。迄今为止,枯草芽孢杆菌已成为最常用的原核表达系统之一,大量的原核和真核基因在枯草芽孢杆菌及其近缘种中得到克隆和表达,有的已应用于工业化生产,取得了不少成果[14-17]。Zobel等[14]在枯草芽孢杆菌中第一个表达编码小分子环状肽形成的真核生物的非核糖体肽合成酶基因,从而证明了枯草芽孢杆菌是适合异源表达真核非核糖体合成蛋白的重要表达系统。
1.2.2 真核表达系统 此类常用酵母表达系统,动植物细胞较其而言,培养条件复杂、生长较慢,并不宜大规模生产。目前动植物在抗菌肽的生产表达上主要有微藻表达系统和其他转基因动植物。微藻表达系统具有低成本、少污染、可再生、大生物量等特性,因此其是继大肠杆菌和酵母菌表达系统之后较有前景的表达平台。Chen等[18]将兔防御素NP-l基因转入小球藻,并且建立了高效的单细胞真核藻类表达系统进行表达。他们通过体外离体抑菌试验证明,NP-1基因已稳定整合到小球藻基因组中,并进行了正确转录和表达。此外,Niu等[19]用Phaeodactylum tricornutum的硝酸还原酶的启动子和终止子来优化Chlorella vulgaris,并以氯霉素乙酰转移酶(chloramphenic awtyltransferase,CAT)作为报告基因构建好载体后,用电穿孔方法转入小球藻中,最后发现转基因小球藻不仅能够在选择性培养基上生长,而且具有很高的CAT表达量。目前的实验来看,通过抗菌肽转基因改造的动植物能抗虫抗菌且稳定性强。Zeitler等[20]设计了几种抗菌肽将其插入病毒载体中,再将重组子导入烟草内,结果得到0.025 mg(肽)/ g(受感染叶生物量),且转基因烟草具有较好的生物活性。Li等[21]将CFG、抗菌肽Cecropin基因5'和3'端的片段等融合在同一载体上并将其转入蚕体内,得到了具有抗菌效果的转基因蚕茧。由此可见,转抗菌肽基因动植物不仅对抗菌肽和转基因的基础研究有着重要意义,而且也将使烟草、丝绸等行业具有巨大的商业潜力。
酵母表达系统有很多优势,如表达调控机理较清楚、可修饰真核生物蛋白质、易大规模发酵等。常用的酵母表达系统有酿酒酵母和毕赤酵母。酿酒酵母的生理过程会导致自身生长受限,从而降低了目的蛋白的产量,所以其在大规模生产中使用较少。目前最常用的是以甲醇为唯一碳源的毕赤酵母。其具有好氧生长和表达量高等特点,所以培养成本相对较低,可以支持高密度细胞培养。金小宝等[22]将分泌型重组表达质粒pPIc9K/Defen8in线性化后转入巴斯德毕赤酵母GSll5中,并用0.5%的甲醇在28℃条件下诱导。结果显示抗茵肽Defensin在毕赤酵母中得到了成功表达,且表达产物对大肠杆菌E. coli K12D31的生长有抑制作用。随着发酵方法的建立,毕赤酵母以其独特优势和潜力被广泛应用,但仍有不足的地方,如甲醇的去除和过糖基化等。如今科研工作者仍在不断研究,以使毕赤酵母更适用于大规模工业化生产。
1.2.3 杆状病毒表达系统 杆状病毒表达系统适用范围广且表达效率高。常用的Bac-To-Bac系统具有周期较短、易纯化、能大量重组等优点。Li等[23]将EG I基因转入到 Bac-to-Bac/BmNPV突变表达体系中发现,所得的蛋白活性和家蚕细胞中表达的蛋白相比提高了22.71%,与被未突变的Bac-to-Bac病毒侵染后的家蚕细胞中的蛋白相比提高了24.71%。与真原核表达系统相比,杆状病毒表达系统操作相对简便,昆虫细胞较易培养且成本低,外源基因表达量高,具有蛋白翻译后修饰功能。然而杆状病毒表达系统也具有一定缺陷。例如,虽然其糖基化位点与哺乳动物细胞一样,但寡糖链性质有所不同,无法产生复杂糖基侧链。因此在使用杆状病毒载体表达某些抗原时,无法保持其天然状态。另外用该系统表达的抗菌肽种类较少,这也可能与杆状病毒宿主范围狭窄有关。
1.3 表达载体的优化与选择
抗菌肽常用的表达载体有大肠杆菌表达载体、酵母表达载体以及两者都适合的穿梭表达载体等,优势各异。其主要的改造研究如下。
1.3.1 宿主菌偏好型密码子 有时人们使用的载体不一定完全适合表达系统。如人的载体在大肠杆菌中表达,就可能因宿主细胞对密码子偏好性不同而导致蛋白表达产量不佳。若构建表达载体时使用宿主菌偏好型密码子,便可使载体有效表达出抗菌肽。Ren等[24]根据已知的抗菌肽基因SMAP-29的氨基酸序列,参照毕赤酵母密码子偏好性,设计并合成SMAP-29的成熟基因片段,且在诱导表达的第2天检测到了预期的SMAP-29,并对金黄色葡萄球菌和白色念球菌都有明显的抑菌效果。
1.3.2 不需要酶切的表达载体 以往在构建融合肽或杂合肽时需对产物进行酶切,使抗菌肽得到纯化富集,但这些过程会对抗菌肽结构性质产生不良影响,使产物产量和质量受到损害。近年来,出现了一些不需酶切就可使抗菌肽表达后自身进行剪切或者剪接的载体。利用intein融合表达抗菌肽只需亲和层析及在柱切割,操作简便,成本低廉。谢永刚[25]构建OG2-intein2-CBD融合蛋白重组子,并且通过条件优化如降低诱导温度及诱导剂IPTG浓度,有效提高蛋白的可溶表达。2A自剪辑技术的应用可将多个抗菌肽融合表达,然后经过自身剪辑使其各自行使功能,有利于抗菌肽协同表达和作用[26-28]。
1.3.3 5'非翻译区序列改建 5'UTR对蛋白表达的影响主要表现在mRNA翻译水平上。如在巴氏德毕赤酵母中表达时,mRNA 5'非翻译区序列应与毕赤酵母AOXImRNA 5'非翻译区序列一致。Staley等[29]研究发现AOX 1的5' UTR中存在着复杂的正面和负面的顺式作用元件,其活性作用不仅仅与其长度有关,更与其改变的位置有关。明飞平等[5]在实验中将抗菌肽重组表达质粒经PCR敲除5'端多余核苷酸序列后再经PCR、酶切构建5' UTR已删除8个核苷酸的新型毕赤酵母表达载体,使两者mRNA 5'非翻译区序列相同,从而提高了抗菌肽PR39在毕赤酵母中的表达及其抑菌活性。
1.4 计算机模拟预测
随着计算机技术飞速发展,很多软件和计算机方法运用于科学研究和生产实践。越来越多的生物工作者采用计算机方法减轻工作量。
目前,其在两大方面对抗菌肽的工作有指导作用:一是其丰富而信息量宏大的数据库;二是各种软件技术的使用。一些常用的数据库,如表1所示。

表1 各类抗菌肽主要数据库
其能提供氨基酸组成、空间结构等信息,对科研工作有很大的指导作用。各种软件技术(如ProtParam、NNPREDICT等)的发展,利于人们对抗菌肽合成、结构等进行较合理的预测。Lata等[30]新研发了AntiBP2,旨在为快速增长的抗菌肽研究领域提供一个有效、快速而方便的分类和设计预测方法。Avram等[31]用计算机软件计算黄蜂毒素衍生物的疏水性、氢键等参数,建立了3D-QSAR模型并合成新的衍生物,部分衍生物比母体具有更好的抗菌活性。于岚岚等[9]在实验中将新型抗茵肽的设计、活性研究及与磷脂相互作用与计算模拟相结合,得到有较强抑菌活性的抗菌肽。抗菌肽的选择性和抗菌活性受到多种因素影响,可用计算机辅助手段分析,对抗菌肽进行修饰改造,得到大量结构全新的抗菌肽,并得出合理的使用方法和趋势,设计出有高选择性、高抗菌活性、低溶血性的抗菌肽,为化学和基因工程方法合成抗菌肽提供参考。
2 生产应用
抗菌肽经过几十年的研究发展,已有很多成品出现在市场上,给生产生活带来新的变化与发展。而将抗菌肽从上游研发阶段带入市场的关键就是发酵生产和分离纯化及检测过程。Li等[16]从物理参数和金属无机盐两方面探索了提高枯草芽孢杆菌BS501a拮抗代谢反应发酵的优化条件。他们通过单因素轮换和正交实验得到了发酵最佳物理参数应是初始pH为7.0,培养温度30℃和发酵时间48 h,并探索出在金属无机盐中硫酸镁、硫酸锰(尤其是硫酸镁)对发酵起着重要的影响。由此可见,一个完整的发酵过程受到很多方面的影响,而要在诸多因素中选出合适的条件,通常难以通过完整的发酵过程进行探索或验证。因此,必须借助计算机软件和数学模型对重要参数进行预测,比较优化选择。其一般步骤,见图1。
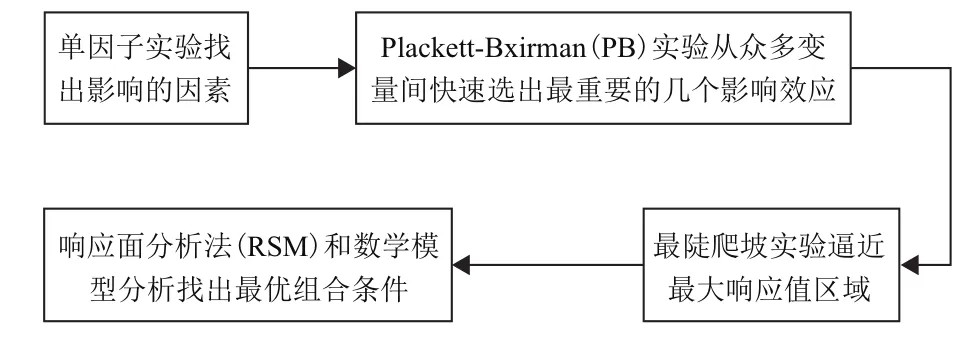
图1 发酵过程优化选择步骤
发酵方式主有固体发酵和液体发酵。液体发酵的主要特点是能够实现高密度发酵、机械化程度高。固体发酵主要是在低湿含量的固态培养基上(aw一般为0.4-0.90),在不腐烂和自然状态下进行的发酵过程。其主要有设备简单、操作简便、原料来源广等特点。选用不同的发酵方式对最终的发酵结果有很大影响。Zhu等[32]通过傅里叶变换红外光谱、光学成像技术、高效液相色谱等技术分析了Bacillus amyloliquefaciens XZ-173在固态发酵和液体发酵两种方式下发酵的脂肽组成、性质以及这两种条件下的转录水平差异。他们最终发现在这两种发酵方式下,脂肽的极性和结构没有太多差异,但是液态发酵有较高的氨基酸比例以及较好的乳化和拮抗活性。在固体发酵条件下,脂肽的合成基因srfAA和sfp有较高积累,并且ituD和lpa-14的转录水平在很长一段时间内不断升高。由此可见,固体发酵和液态发酵各自有其独特的优势,需要根据实际生产的需求不断探索,选择合适的发酵方式。
除了借助计算机软件和发酵方式外,菌的培养发酵设备也是值得改进的重要内容。陶德录等[33]报告了一套在上海实施了生产量达2 000 t/年抗菌肽的新技术、新工艺和新设备,为抗菌肽的规模化生产探索了一条新途径。李建华等[34]设计了一种新型的带有上下翻盖的托盘平动固态发酵机,他利用Matlab对发酵罐内托盘的3种排列方式下的各种情况进行了基于托盘运动不干涉的各机构参数的初步求解和精确求解,对喷雾接种时生物颗粒流体系统的两相流进行了分析,并采用 CFD 计算流体动力学软件仿真和实验验证的方法对喷头类型、喷雾参数等因素对喷雾接种效果的影响进行了系统的研究。最后实验和仿真结果表明,用喷雾接种实现纯种固态发酵过程的自动化、保证接种的均匀性是可行的,涡流扇形雾喷头 TT1102 在喷雾压力为 0.3 MPa 时的能量耗散率最低、菌种死亡率最小。这种设备通过发酵过程的自动控制,创造良好的生产条件,保证真正意义的纯种固态培养,可以得到生长好且质量均匀的产品,同时又便于工业规模的放大。
中游发酵完成之后,下游的分离纯化、包装检测等目前都是一些较为常规的技术,比如产品浓缩时,工业生产上多采用旋转蒸发、喷雾干燥、冷冻浓缩等方法将产品制成结晶、干粉等形式,便于产品保存和后期检验及再加工等操作。鉴定是否为目的抗菌肽常用方法有质谱分析、电泳、PCR等,对产品抑菌性质进行检测常用的方法是琼脂孔穴扩散法。当然,在产品形式设计与包装中也应该体现出科学性。现在生产生活中已有多种形式的抗菌肽制剂,如粉制剂、喷雾制剂、凝胶制剂等。这些形式都是根据公司产品特点和市场适应性设计包装的。而抗菌肽在应用时也应该探索合适的条件,注意一些影响因素。如Terova等[35]在研究中指出内源性抗菌多肽能直接快速杀灭病原体,但其mRNA的表达量受到急性胁迫压力的调控影响。这一发现将对鱼类的健康和成功养殖有着重大意义。Wang 等[36]通过实验发现带锌甲硫氨酸和抗菌肽具有协同作用,能够更好提高断龄猪仔的免疫力、提高生长性能。Nithya等[37]发现用来自Bacillus licheniformis Me1的部分纯化抗菌肽与LDPE和纤维素材料制成活性薄膜,其在食物的抗菌保鲜方面可有重要作用。
3 展望
在这个亟需倡导绿色环保的时代,抗生素的使用已经无法满足人们对更高质量生产生活的追求。抗菌肽应时而生,自被人类发现以来展现出来其独特的众多优势和潜力,作为抗生素的替代者而备受期待。在这30多年的发展中,抗菌肽的研发和生产取得了很大进步,已经能够通过工业生产出良好的商品进入人们生产生活的方方面面,如美容行业、饲料行业、医药业及食品添加剂等等。其中抗菌肽又在饲料行业应用颇多,只是其作为饲料在实际应用中也面临着许多的问题,如如何不影响动物的食欲、提高其免疫力、增强肉质等。而且如何大规模地生产优化的抗菌肽使其满足人们的生产生活也是一大难题。但随着科技的不断发展、经验的不断增加,人们对抗菌肽的分子改造和生产研发将有着越来越多的认识和改进,抗菌肽的大规模工业化生产也会日益成熟,而这将是“后抗生素时代”的一条全新出路。
[1]Li W, Tailhades J, O'Brien-Simpson NM, et al. Proline-rich antimicrobial peptides:potential therapeutics against antibioticresistant bacteria[J]. Amino Acids, 2014, 46(10):2287-2294.
[2]Lico C, Santi L, Twyman R, et al. The use of plants for the production of therapeutic human peptides[J]. Plant Cell Rep, 2012, 31(3):439-451.
[3]Di Luca M, Maccari G, Nifosi R. Treatment of microbial biofilms in the post-antibiotic era:prophylactic and therapeutic use of antimicrobial peptides and their design by bioinformatics tools[J]. Pathog Dis, 2014, 70(3):257-270.
[4]da Costa JP, Cova M, Ferreira R, et al. Antimicrobial peptides:an alternative for innovative medicines?[J]. Applied Microbiology and Biotechnology, 2015, 99(5):2023-2040.
[5]明飞平, 杨军, 朱进美, 等. 5'非翻译区序列改建提高抗菌肽PR39表达[J]. 生物技术, 2013, 33(12):86-91.
[6]刘忠渊, 毛新芳, 张兰廷, 等. 新疆家蚕抗菌肽基因突变及其抗菌活性的研究[J]. 生物技术通报, 2011(9):147-156.
[7]van Dijk A, Molhoek EM, Velduizen EJ, et al. Identification of chicken cathelicidin-2 core elements involved in antibacterial and immunomodulatory activities[J]. Mol Biosyst, 2012, 8(2):587-594.
[8]Jittikoon J, Ngamsaithong N, Pimthon J, et al. Effect of N-terminal truncation on antibacterial activity, cytotoxicity and membrane perturbation activity of Cc-CATH3[J]. Archives of Pharmacal Research, 2015, 38(10):1839-1849.
[9]于岚岚, 冉瑜, 白希希, 等. 新型抗茵肽的设计、活性研究及与磷脂相互作用的计算模拟[J]. 高等学校化学学报, 2012(33):268l-2687.
[10]Ahn M, Murugan RN, Jacob B, et al. Discovery of novel Histidine derivedlipo-aminoacid:Applied in the synthesis of ultrashort a ntimicrobial peptidomimetic having potent antimicrobial activity,salt resistance and proteasestability[J]. Eur J Med Chem, 2013,68:10-18.
[11]Yu FX, Wang JG, Zhang P, et al. Fusion expression of cecropin B-like antibacterial peptide in Escherichia coli and preparation of its antiserum[J]. Biotechnology Letters, 2010, 32:669-673.
[12]Wang Z, Zhang L, Wang J, et al. Synergistic interaction of PMAP-36 and PRW4 with aminoglycoside antibiotics and their antibacterial mechanism[J]. World J Microbiol Biotechnol, 2014, 30(12):3121-3128.
[13]Wu RJ, Wang Q, Zheng ZJ, et al. Design, characte-rization and expression of a novel hybrid peptides melittin(1-13)-LL37(17-30)[J]. Mol Biol Rep, 2014, 41:4163-4169.
[14]Zobel S, Kumpfmüller J, Süssmuth RD, et al. Bacillus subtilis as heterologous host for the secretory production of the non-ribosomal cyclodepsipeptide enniatin[J]. Appl Microbiol Biotechnol, 2015,99:681-691.
[15]Lee NK, Yeo IC, Park JW, et al. Growth inhibition and induction of stress protein, GroEL, of Bacillus cereus exposed to antibacterial peptide isolated from Bacillus subtilis SC-8[J]. Appl Biochem Biotechnol, 2011, 165:235-242.
[16]Li RF, Xu Y. Fermentation optimization to improve production of antagonistic metabolites by Bacillus subtilis strain BS50la[J]. Journal of Central South University of Technology(English Edition), 2011, 18(4):1047-1053.
[17]Liu L, Liu YF, Shin HD, et al. Developing Bacillus spp. as a cell factory for production of microbial enzymes and industrially important biochemicals in the context of systems and synthetic biology[J]. Appl Microbiol Biotechnol, 2013, 97:6113-6127.
[18]Chen Y, Wang YQ, Sun YG, et al. Highly efficient expression of rabbit neutrophil peptide-1 gene in Chlorella ellipsoidea cells[J]. Current Genetics, 2001, 39(5-6):365-370.
[19]Niu YF, Zhang MH, Xie WH, et al. A new inducible expressionsystem in a transformed green alga, Chlorella vulgaris[J]. Genetics and Molecular Research, 2011:3427-3434.
[20]Zeitler B, Bernhard A, Meyer H, et al. Production of a de-novo designed antimicrobial peptide in Nicotiana benthamiana[J]. Plant Molecular Biology, 2013, 81(3):259-272.
[21]Li Z, Jiang Y, Cao GL, et al. Construction of transgenic silkworm spinning antibacterial silk with fluorescence[J]. Mol Biol Rep,20154, 2:19-25.
[22]金小宝, 王婷婷, 朱家勇, 等. 家蝇抗菌肽Defensin在毕赤酵母中的表达及活性鉴定[J]. 生物技术通报, 2010(9):194-197.
[23]Li XH, Wang D, Zhou F, et al. Cloning and expression of a cellulase gene in the silkworm, Bombyx mori by improved Bac-to-Bac/ BmNPV baculovirus expression system[J]. Molecular Biology Reports, 2010, 37(8):3721-3728.
[24]Ren YJ, He EP, Wang XH, et al. Codon optimization of SMAP-29 gene and its expression in Pichia pastoris[J]. Agricultural Biotechnology, 2012, 1(4):49-53.
[25]谢永刚. 蛙抗菌肽Palustrin-OGl的分子改良及改良肽的生物学活性、抗菌机制和重组表达研究[D]. 杭州:浙江大学,2012.
[26]Tang WN, Ehrlich I, WolffSBE, et al. Faithful expression of multiple proteins via 2A-peptide self-processing:a versatile and reliable method for manipulating brain circuits[J]. The Journal of Neuroscience, 2009, 29(27):8621-8629.
[27]Mariano A, Li X, Han RZ. Highly efficient genome editing via 2A-coupled co-expression of two TALEN monomers[J]. BMC Research Notes, 2014, 7:628.
[28]Kim JH, Lee SR, Li LH, et al. High cleavage efficiency of a 2A peptide derived from porcine teschovirus-1 in human cell lines,zebrafish and mice[J]. PLoS One, 2011, 6(4):e18556.
[29]Staley CA, Huang A, Nattestad M, et al. Analysis of the 5' untranslated region(5'UTR)of the alcohol oxidase 1(AOX1) gene in recombinant protein expression in Pichia pastoris[J]. Gene, 2012, 496(2):118-127.
[30]Lata S, Mishra NK, Raghava GP. AntiBP2:improved version of antibacterial peptide prediction[J]. BMC Bioinformatics, 2010,11(Supp l):S19.
[31]Avram S, Buin C, Borcan F, et al. More effective antimicrobial mastoparan derivatives, generated by 3D-QSAR-almond and computational mutagenesis[J]. Mol Biosyst, 2012, 8(2):587-594.
[32] Zhu Z, Zhang JC, Wu YL, et al. Comparative study on the properties of lipopeptide products and expression of biosynthetic genes from Bacillus amyloliquefaciens XZ-173 in liquid fermentation and solidstate fermentation[J]. World J Microbiol Biotechnol, 2013, 29:2105-2114.
[33]陶德录, 李巧贤. 抗菌肽规模化生产新工艺新设备的研究和应用[J]. 饲料工业, 2010, 31(21):4-6.
[34] 李建华. 多功能纯种固态发酵设备及其喷雾接种技术研究[D]. 南京:南京林业大学, 2013.
[35]Terova G, Cattaneo AG, Preziosa E, et al. Impact of acute stress on antimicrobial polypeptides mRNA copy number in several tissues of marine sea bass(Dicentrarchus labrax)[J]. BMC Immunology,2011, 12:69.
[36]Wang JH, Wu CC, Feng J. Effect of dietary antibacterial peptide and zinc-methionine on performance and serum biochemical parameters in piglets[J]. Czech Journal of Animal Science,2011, 56(1):30-36.
[37]Nithya V, Murthy PSK, Halami PM. Development and application of active films for food packaging using antibacterial peptide of Bacillus licheniformis Me1[J]. Journal of Applied Microbiology,2013, 115(2):475-483.
(责任编辑 狄艳红)
Research Progress of Efficient Expression and Optimization of Production of Antibacterial Peptide
YANG Ping1YUAN Yi-hao2YANG Xiao-li1ZHONG Yue2LIAO Yu-xin2GAO Rong1
(1. College of Life Science,Sichuan University,Chengdu 610064;2. College of Polymer Science and Engineering,Sichuan University,Chengdu 610064)
Antimicrobial peptides(AMPs),a class of polypeptides induced by external conditions,are encoded by specific genes. In the era of facing the issue of “drug resistance” caused by antibiotics,AMPs own huge potential of benefits to mankind and replacing the antibiotics due to their characteristics of broad-spectrum antibacterial activity,scarce generation of drug-resistance,and antitumor virus,etc. However,the antibacterial activity and instability of natural AMPs should be enhanced,their toxicity should be reduced,and the large-scale production may be achieved by improving the industrial process. Therefore,the recent researches on the molecular modification and design,production and application of AMPS are summarized in detailed here.
antimicrobial peptides;molecular design;expression strategies;production optimization
10.13560/j.cnki.biotech.bull.1985.2016.03.005
2015-06-11
国家自然科学基金项目(30871855),科技部国际科技合作项目(2011DFA10101103),四川大学大学生创新基金资助(208240-4184011,2020404131001)
杨平,女,研究方向:分子免疫、生物技术;E-mail:523008626@qq.com
高荣,教授,研究方向:分子遗传和基因工程、分子免疫学、生物技术、微生物学和动物传染病等;E-mail:gaorong96@163.com
猜你喜欢
华人时刊(2022年9期)2022-09-06
现代畜牧科技(2021年9期)2021-10-13
华人时刊(2020年15期)2020-12-14
中国调味品(2017年2期)2017-03-20
广东饲料(2016年5期)2016-12-01
创新作文(小学版)(2016年16期)2016-11-11
现代检验医学杂志(2016年5期)2016-08-20
国外医药(抗生素分册)(2016年5期)2016-07-12
广州大学学报(自然科学版)(2015年4期)2015-12-23
中国科技信息(2015年2期)2015-11-16
