
不同根际溶解氧质量浓度对生育中后期水稻根系和抗氧化酶活性的影响
2016-09-25 11:38:36王佳佳杜彦修李俊周孙红正赵全志
河南农业大学学报 2016年6期
王佳佳,彭 廷,张 静,刘 娟,陈 浩,杜彦修,李俊周,孙红正,赵全志
(河南农业大学农学院,河南 郑州 450002)
不同根际溶解氧质量浓度对生育中后期水稻根系和抗氧化酶活性的影响
王佳佳,彭 廷,张 静,刘 娟,陈 浩,杜彦修,李俊周,孙红正,赵全志
(河南农业大学农学院,河南 郑州 450002)
为探究溶解氧质量浓度对不同基因型水稻品种生育中后期根系生长发育的影响。以4种不同基因型水稻为试验材料,采用水培法,研究不同溶解氧质量浓度对水稻生育中后期根系形态、根系抗氧化酶活性及可溶性蛋白质含量的影响。结果表明,生育中后期高氧处理可显著增加水稻的根长和根条数,低氧处理对水稻根系的伸长影响不大,根条数有所增加,均未达到显著水平;高氧处理可显著增加抗氧化酶活性及可溶性蛋白质含量,说明水稻生育中后期增加根际溶解氧质量浓度有利于水稻根系的生长发育,有效延缓根系衰老。
水稻;根系;溶解氧质量浓度;抗氧化酶
水稻生长发育中后期是产量形成的关键时期,而此时水稻根系逐渐衰老,叶片的功能及子粒的灌浆也随之受到影响。因此,如何延缓生育中后期根系的衰老对水稻产量的提升具有重要意义。在中国传统水稻栽培中,常通过水旱轮作、中耕耘田、排水晒田等调控措施来增加土壤的通透性,改善根际的氧环境[1, 2]。近年来,通过干湿交替灌溉、增施氧肥(过氧化尿素、过氧化钙)、超微气泡灌溉系统及其他物理增氧措施,研究氧对水稻根系生长、氮代谢及产量影响[3]已取得较大进展。已经证明在不同的增氧模式(追施氧肥、干湿交替)下,灌浆期水稻地上地下部衰老均有所减缓[4];超微气泡水增氧灌溉技术不仅能提高杂交水稻的抗氧化系统的活性,增强水稻生育后期的根系功能,还能延缓生育后期水稻根系和叶片的衰老[5];增氧和铵硝混合营养对促进水稻根系生长和氮素积累具有正互作效应[6];连续增氧使水稻根比表面积增加、根系氧化强度增强[7]。前人研究成果中,有关根际溶解氧质量浓度对水稻根系生长发育影响的研究主要集中在移栽缓苗后,而针对生育中后期溶解氧质量浓度对根系生长的影响研究较少。因此,本试验以不同基因型的水稻品种为材料,通过物理措施创造水稻根际高氧和低氧环境,研究根际溶解氧质量浓度对生育中后期水稻根系生长发育的影响,为水稻高产高效栽培提供理论依据。
1 材料与方法
1.1试验设计1.1.1 试验材料 选取新稻18(粳稻,Xindao18),金农丝苗(籼稻,Jingnongsimiao),Y两优2号(杂交稻,Y liangyou 2)以及IRAT 109(旱稻)为试验材料。
1.1.2 秧苗培育 2014年在河南农业大学水稻工程技术研究中心实验室和河南农业大学郑州科教园区进行试验。培养方法如表1。2014-05-05在河南农业大学科教园区采用塑料软盘单粒播种,每盘288孔,湿润育秧。六叶期后采用营养液水培法,将秧苗单本移栽至漂浮于水面上的泡沫板,每板75孔。预水培期水培池规格:1.24 m×6.10 m×0.26 m,处理期水培池规格:5.20 m×2.50 m×1.00 m,移栽行株距25.00 cm×12.70 cm。营养液组成采用国际水稻研究所(IRRI)配方,其中大量元素N,P,K,Ca,Mg所用试剂略有改变,分别由CO(NH2)2,KH2PO4,Ca(NO3)2·4H2O,MgSO4·7H2O提供,Si及其他微量元素不变。水培营养液采用HCl和NaOH调pH值至5.0~6.0,每3 d调1次。
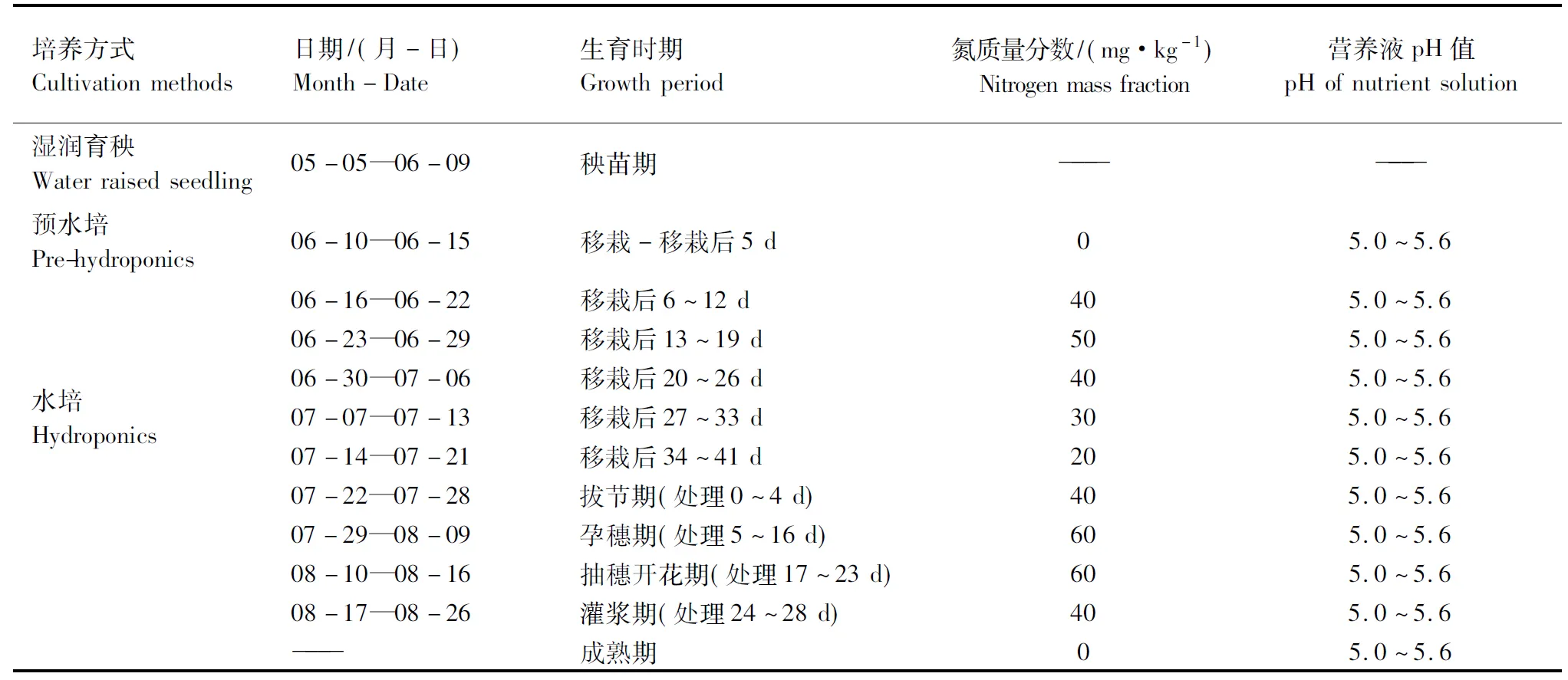
表1 全生育期的培养方法Table 1 The cultivation methods during full growth period
1.1.3 试验设计
水稻生育中后期设置3个根际溶解氧质量浓度处理,分别为低氧、对照和高氧。低氧利用溶氧仪实时监测溶解氧质量浓度,并通过与氮气罐、电磁阀、空气泵连接实现自动调控;对照和高氧处理,将溶氧仪与氧气罐、电磁阀连接,根据溶氧仪检测到的溶解氧质量浓度使电磁阀打开或关闭,从而实现自动充气或断气。低氧、对照和高氧处理溶氧仪设定的溶解氧质量浓度分别为2,5和10 mg·L-1。
1.2测定项目
1.2.1 根系形态 分别于处理后7 d(孕穗期)、18 d(抽穗开花期)和28 d(灌浆期)取样。每处理5次重复。根系经自来水洗净后,分别测量单株根条数、最长根长。
1.2.2 超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性及可溶性蛋白质含量测定 分别于处理后7,18和28 d取样,每处理5次重复,植株根系经自来水洗净后,液氮速冻后存于-40 ℃冰箱,取0.5 g根系样品于预冷研钵中,加1 mL 0.05 mol·L-1PBS(pH值 7.8)在冰浴下研磨成匀浆,定容为6 mL,4 ℃,10 000 r·min-1离心20 min,上清液即为酶粗提液。该酶粗提液用于超氧化物歧化酶(SOD)、过氧化氢酶(CAT)及可溶性蛋白质含量的测定。具体操作参照赵世杰等[8]的方法。
1.3数据处理
选取妇科常见的宫颈癌和子宫内膜癌患者作为本次研究对象。整理收集2017年1—11月我院妇科治疗的30例宫颈鳞癌患者的宫颈病理组织作为研究组1的研究对象,同时相应选取30例经宫颈液基细胞学证实无异常的宫颈病理标本作为对照组1的研究对象。另选取30例子宫内膜腺癌患者的子宫内膜病理标本作为研究组2的研究对象,同时选取30例子宫平滑肌瘤患者正常子宫内膜病理标本作为对照组2。所有患者术前均未接受放、化疗等治疗;患者均为已婚已育妇女。
利用Microsoft Excel 2007进行数据整理,SPSS 19进行方差分析和多重比较分析(Duncan法,α=0.05)。
2 结果与分析
2.1不同根际溶解氧质量浓度对水稻最长根长的影响
由表2可知,不同根际溶解氧质量浓度对水稻最长根长的影响随溶解氧质量浓度的提高逐渐增大,总体表现为低氧抑制根系的伸长,高氧促进根系的伸长,且4个品种表现一致。处理28 d时,4个品种高氧处理下最长根长均显著高于对照和低
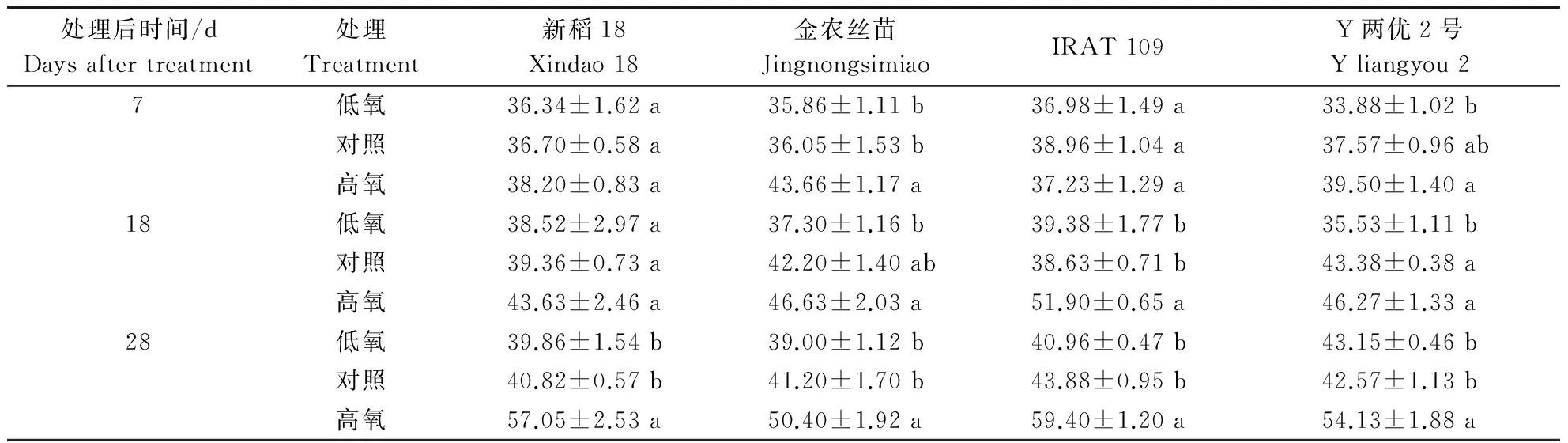
表2 不同根际溶解氧质量浓度对不同基因型水稻品种最长根长的影响Table 2 Effects of different dissolved oxygen mass concentration on the longest root length of different cultivars cm
注:数据为5个重复的平均值±标准误。不同字母表示多重比较Duncan检验达到5%的显著水平。下同。
Note: The data represent the mean ±SE(n=5). Different letters show significant difference according to Duncan test (0.05). The same as below.
氧处理。与对照相比,高氧处理增幅分别为39.76%(新稻18),22.33%(金农丝苗),35.38%(IRAT 109),27.15%(Y两优2号);与对照相比,低氧处理最长根长有所降低,但未达到显著水平。2.2不同根际溶解氧质量浓度对水稻总根条数的影响
由表3可知,随着处理质量浓度的提高,4个水稻品种的单株根条数均呈逐渐增加的变化趋势,且存在明显的基因型差异.在整个处理过程中,除低氧处理28 d外,均表现为新稻18﹥Y两优2号﹥金农丝苗﹥IRAT 109。在处理后18和28 d时,高氧处理下,4个水稻品种的单株根条数均有所提高,且新稻18和Y两优2号在处理后28 d,单株根条数显著高于对照,分别较对照增加48.11%和71.58%;处理后7,18和28 d时,低氧处理下,4个水稻品种的单株根条数均高于对照,除处理后28 d旱稻IRAT 109在低氧条件下单株根条数显著高于对照外,其他品种均未达到显著水平。
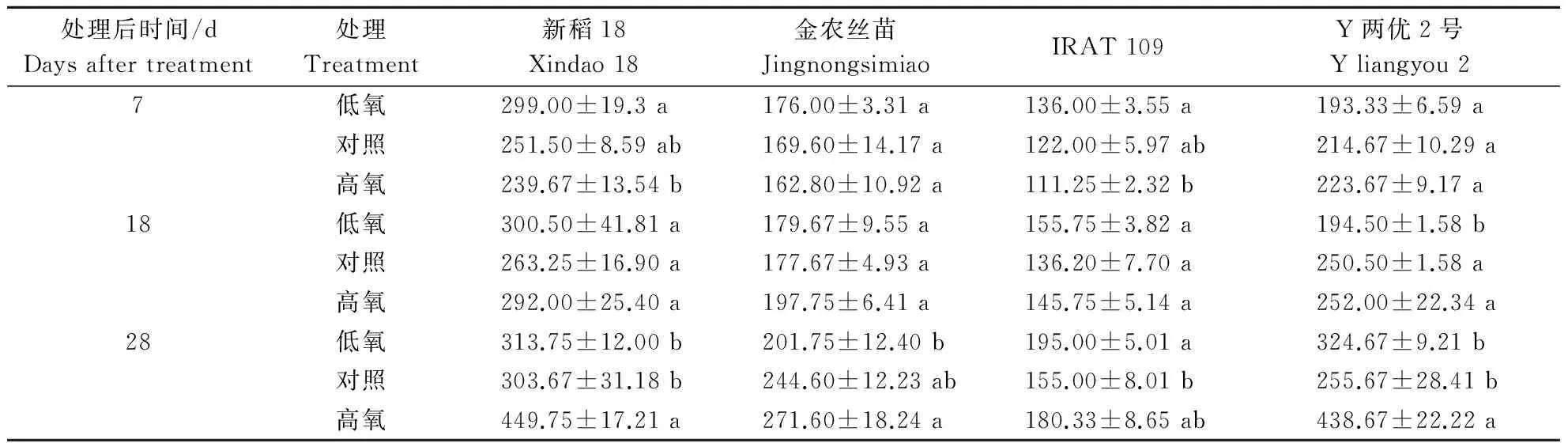
表3 不同根际溶解氧质量浓度对水稻总根条数的影响Table 3 Effects of different dissolved oxygen mass concentrations on total root numbers of rice
2.3不同根际溶解氧质量浓度对水稻根系超氧化物歧化酶(SOD)活性的影响
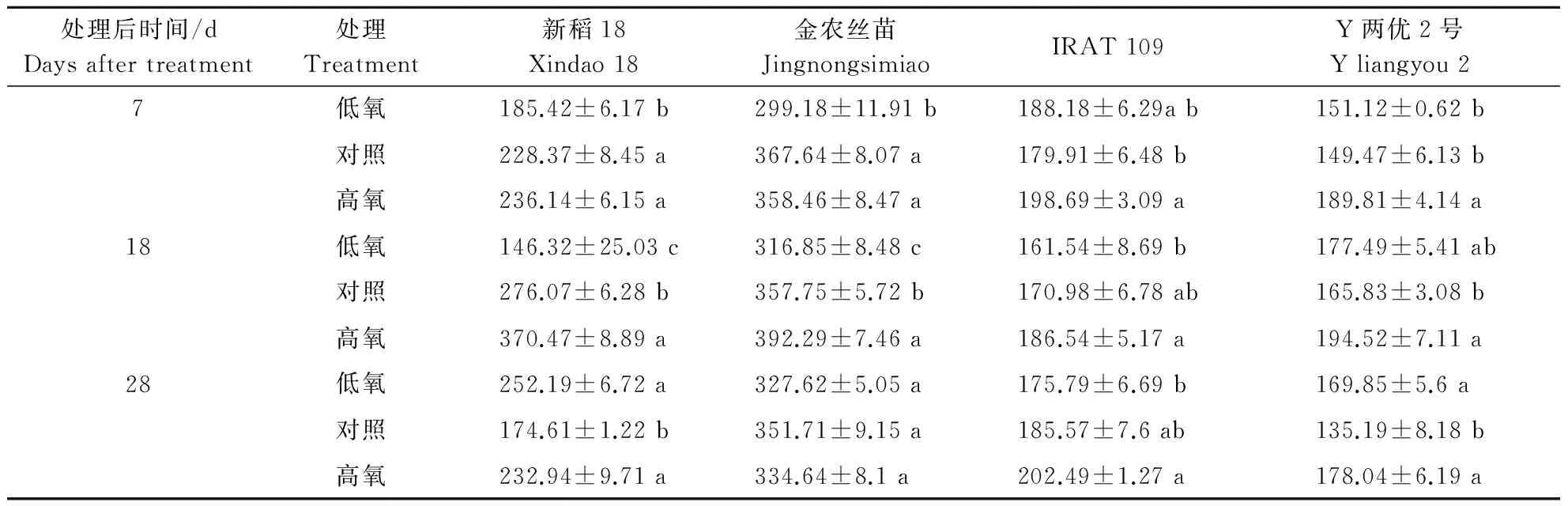
表4 不同根际溶解氧质量浓度对不同基因型水稻品种根系超氧化物歧化酶活性的影响Table 4 Effects of different dissolved oxygen mass concentration on SOD activity in root of different cultivars U·g-1
2.4不同根际溶解氧质量浓度对水稻根系过氧化氢酶(CAT)活性的影响
由表5可知,4个品种根系CAT活性同样具有明显的基因型差异,金农丝苗和Y两优2号根系CAT活性显著高于新稻18和旱稻IRAT 109。新稻18、旱稻IRAT 109和Y两优2号在处理末期均表现为高氧﹥低氧﹥对照,且高氧处理与对照、低氧间存在显著差异。新稻18和Y两优2号根系CAT活性在对照条件下随处理的质量浓度提高呈缓慢下降的变化趋势,在高氧处理下呈先增加后减少的变化趋势,而低氧处理下变化不大;Y两优2号根系CAT活性在低氧和高氧处理下均显著高于对照;金农丝苗和旱稻IRAT 109根系CAT活性随处理的质量浓度提高呈逐渐下降的变化趋势。
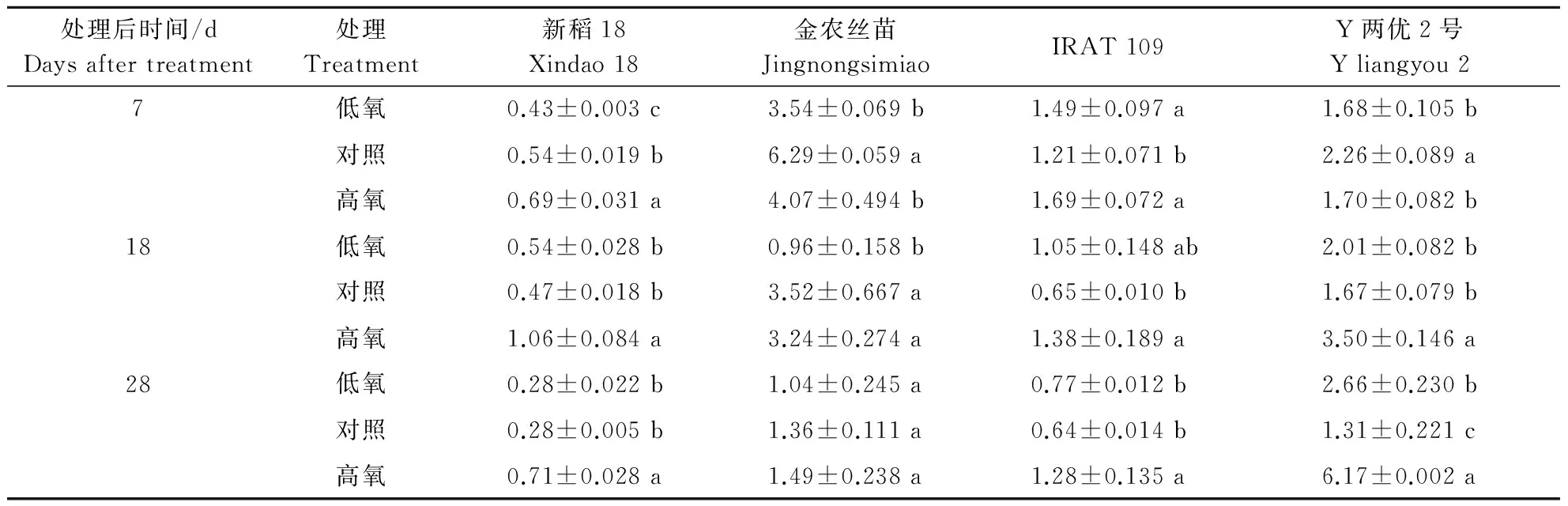
表5 不同根际溶解氧质量浓度对不同基因型水稻品种根系过氧化氢酶活性的影响Table 5 Effects of different dissolved oxygen mass concentration on the CAT activity in root of different cultivars A240·g-1·min-1
2.5不同根际溶解氧质量浓度对水稻根系可溶性蛋白质含量的影响
由表6可知,新稻18在处理期间,根际溶解氧质量浓度越高,根系可溶性蛋白质含量越高,且在处理初期,3个处理间差异达到显著水平,其中高氧处理下根系可溶性蛋白质含量为对照的1.31倍,对照是低氧处理下的2.14倍;金农丝苗在处理初期,低氧和高氧处理根系可溶性蛋白质含量均显著低于对照,而在处理后期,溶解氧质量浓度越高,根系可溶性蛋白质含量也越高;旱稻IRAT 109在处理初期,溶解氧质量浓度越高,其根系可溶性蛋白质含量越高,而在处理后期,则表现为对照﹥低氧﹥高氧;Y两优2号在处理后期则明显表现为高氧﹥低氧﹥对照。
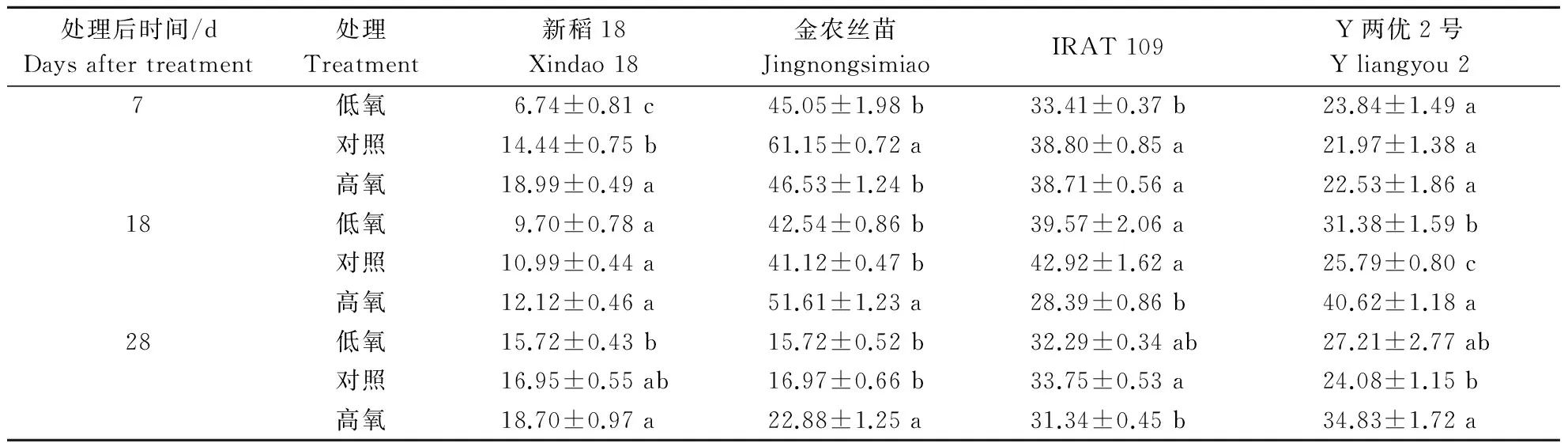
表6 不同根际溶解氧质量浓度对不同基因型水稻品种根系可溶性蛋白质含量的影响Table 6 Effects of different dissolved oxygen mass concentration on the protein contents in root of different cultivars mg·g-1
3 结语与讨论
水稻植株具有依据环境调节根系数量的能力[1]。根际增氧处理后植物根长增长、白根增多[6, 10];根际缺氧处理后植物根系伸长受到抑制,根长变短,黑根增多,根系弱小[11],但显著提高单株不定根数[12]。赵霞等[13]研究表明,适度增氧会促进地上分蘖、叶面积形成,使株高增高,根体积增大,根条数增多,根系吸收功能增强,然而过度增氧会减弱增氧对水稻生长的促进作用,最长根长随溶解氧质量浓度升高而增长。王丹英等[1]研究表明,水稻根条数在缺氧条件下增多,在高氧处理条件下整体上有减少的趋势,下层根系所占比例增加。赵锋等[7]发现,连续增氧后水稻的不定根数目显著减少。KHATOON等[14]通过对大豆幼苗的缺氧胁迫研究发现,在低氧和淹涝胁迫3和6 d后,两处理条件下大豆幼苗根系的生长均受到抑制。本试验研究结果表明,水稻生育中后期较高的根际溶解氧质量浓度会促进根系的伸长和根条数的增加。随着根际溶解氧质量浓度的提高,其促进效果更显著;低氧处理对水稻根系的伸长影响不大,根条数有所增加,但未达到显著水平。
超氧化物歧化酶(SOD)、过氧化氢酶(CAT)及多种过氧化物酶(POX)和抗坏血酸过氧化物酶(APX)等是构成防止细胞过氧化的重要清除酶,其活性高低对维持植物体内活性氧的正常水平,保护细胞膜免受氧化伤害具有重要意义[15]。低氧胁迫下植物体内抗氧化酶活性增加是植物对低氧胁迫的一种适应机制[16]。黄瓜幼苗可通过提高自身抗氧化酶活性来适应低氧胁迫,且低氧胁迫下耐低氧品种比低氧敏感品种抗氧化酶活性要高,但长期低氧胁迫会破坏抗氧化系统,使抗氧化酶活性降低[17],同时低氧胁迫下根系生长明显受到抑制,根系可溶性蛋白质含量降低[18-20],低氧胁迫下甜瓜幼苗根系和叶片抗氧化酶活性和同工酶表达均高于对照[21]。从本研究结果可以看出,经不同溶解氧质量浓度处理后,不同基因型水稻品种根系SOD,CAT酶活性均有不同程度的变化。本试验结果表明,高氧处理显著提高了水稻生长中后期根系SOD和CAT活性;低氧处理显著提高新稻18和Y两优2号处理后期根系SOD活性及Y两优2号根系CAT活性,且旱稻IRAT 109处理前期根系CAT活性也得到显著提高,这可能是品种间生育时期的差异造成的。以上结果表明,与短期低氧胁迫适应性的提高抗氧化酶活性不同。在水稻生长中后期,根系本身已处于逐渐衰老状态,抗氧化酶活性也在逐渐下降,而在高氧处理下,根长伸长,同时根条数也显著增加,进而增强根系的生理功能,使整个根系的抗氧化酶活性处于较高的水平,有利于超氧阴离子的清除,降低膜脂过氧化水平,从而延缓根系衰老,使根系能处于一个相对较好的生长状态。
综上所述,在水稻生育中后期根际高氧处理促进水稻根系的伸长和根条数的增加,低氧处理条件下最长根长变化不大,根条数有所增加,但未达到显著水平。根际高氧处理能在一定程度上增加生育中后期水稻根系抗氧化酶活性及可溶性蛋白质含量,有利于及时清除根系衰老过程中产生的活性氧,从而使植物在生育中后期抗逆性增强,延缓根系衰老,增加根系和地上部的功能期。
[1] 王丹英,韩勃,章秀福,等. 水稻根际含氧量对根系生长的影响[J]. 作物学报,2008,34 (5):803-808.
[2] 张静,刘娟,陈浩,等. 干湿交替条件下稻田土壤氧气和水分变化规律研究[J]. 中国生态农业学报,2014,22 (4):408-413.
[3] 赵锋,王丹英,徐春梅,等. 水稻氧营养的生理、生态机制及环境效应研究进展[J]. 中国水稻科学,2009,23 (4):335-341.
[4] 赵锋,张卫建,章秀福,等. 稻田增氧模式对水稻子粒灌浆的影响[J]. 中国水稻科学,2011,25 (6):605-612.
[5] 朱练峰,刘学,禹盛苗,等. 增氧灌溉对水稻生理特性和后期衰老的影响[J]. 中国水稻科学,2010,24 (3):257-263.
[6] 赵锋,徐春梅,张卫建,等. 根际溶氧量与氮素形态对水稻根系特征及氮素积累的影响[J]. 中国水稻科学,2011,25 (2):195-200.
[7] 赵锋,张卫建,章秀福,等. 连续增氧对不同基因型水稻分蘖期生长和氮代谢酶活性的影响[J]. 作物学报,2012,38 (2):344-351.
[8] 赵世杰,史国安,董新纯. 植物生理实验技术[M]. 北京: 中国农业科学技术出版社,2002.
[10] JACKSON M, COLMER T. Response and adaptation by plants to flooding stress[J]. Annals of Botany, 2005,96(4):501-505.
[11] 韩勃. 增氧条件下水稻根系及地上部生长特性研究[D]: 扬州: 扬州大学, 2007.
[12] 邓丹,吴可为,邓泓. 根区通氧状况对水稻幼苗生长及吸收镉的影响[J]. 生态学报,2009,29 (5):2520-2526.
[13] 赵霞,徐春梅,王丹英,等. 根际溶氧量对分蘖期水稻生长特性及其氮素代谢的影响[J]. 中国农业科学,2015,48 (18):3733-3742.
[14] KHATOON A, REHMAN S, OH M W, et al. Analysis of response mechanism in soybean under low oxygen and flooding stresses using gel-base proteomics technique[J]. Molecular Biology Reports, 2012,39(12):10581-10594.[15] PRASAD TK, ANDERSON M D, MARTIN B A, et al. Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide[J]. The Plant Cell, 1994,6(1):65-74.
[16] USHIMARU T, KANEMATSU S, KATAYAMA M, et al. Antioxidative enzymes in seedlings ofNelumbonuciferagerminated under water[J]. Physiologia Plantarum, 2001,112(1):39-46.
[17] 胡晓辉,郭世荣,王素平,等. 外源亚精胺对根际低氧胁迫下黄瓜幼苗根系多胺含量和抗氧化酶活性的影响[J]. 植物生态学报,2006,30 (1):118-123.
[18] 陆晓民,孙锦,郭世荣,等. 外源 24-表油菜素内酯对低氧胁迫下黄瓜幼苗生长和可溶性蛋白表达的影响[J]. 南京农业大学学报,2011,34 (6):31-35.
[19] 何立中,郭世荣,陆晓民,等. 钙对根际低氧胁迫下黄瓜幼苗可溶性蛋白表达的影响[J]. 南京农业大学学报,2012,35 (1):21-25.
[20] 宋锁玲,李敬蕊,高洪波,等. γ-氨基丁酸对低氧胁迫下甜瓜幼苗氮代谢及矿质元素含量的影响[J]. 园艺学报,2012,39 (4):695-704.
[21] 李敬蕊,杨丽文,王春燕,等. γ-氨基丁酸对低氧胁迫下甜瓜幼苗抗氧化酶活性及表达的影响[J]. 东北农业大学学报,2014,45(11):28-36.
(责任编辑:常思敏)
Effectsofdifferentrhizospheredissolvedoxygenmassconcentrationonricerootgrowthandantioxidantenzymeactivitiesatmiddleandlategrowthstage
WANG Jiajia, PENG Ting, ZHANG Jing, LIU Juan, CHEN Hao, DU Yanxiu, LI Junzhou, SUN Hongzheng, ZHAO Quanzhi
(College of Agronomy, Henan Agricultural University, Zhengzhou 450002, China)
In order to explore the influence of dissolved oxygen mass concentration on the root growth of different rice cultivars at middle and late growth stage, four rice cultivars were cultured in nutrient solution and the dynamic change of root morphology, root antioxidant enzyme, and soluble protein content were measured. The results showed that root length and number were significantly increased under high oxygen mass concentration of rhizosphere, while low oxygen mass concentration inhibited root elongation and increased the root number per plant, but the difference was not significant. In addition, high oxygen mass concentration could significantly increase the antioxidant enzyme activity and soluble protein content. It suggests that the increasing of dissolved oxygen mass concentration is beneficial to rice root growth, thus slowing down the senescence of root at middle and late growth stage of rice.
rice; root; dissolved oxygen mass concentration; antioxidant enzyme
2015-05-09
国家自然科学基金项目(31271651)
王佳佳( 1989-),女,河南舞钢人,硕士研究生,主要从事水稻逆境生理研究。
赵全志( 1968-),男,河南平舆人,教授,博士。
1000-2340(2016)06-0720-06
S 511
:A
猜你喜欢
食品安全导刊(2022年2期)2022-03-18 00:35:00
食品工业(2021年12期)2021-12-31 02:57:08
天津医科大学学报(2021年4期)2021-08-21 02:14:50
科学与信息化(2020年11期)2020-06-19 08:50:42
中国医药导报(2018年36期)2018-03-04 06:50:42
数学小灵通(1-2年级)(2017年12期)2018-01-23 03:37:05
计算机测量与控制(2017年6期)2017-07-01 16:24:28
水利科技与经济(2017年6期)2017-04-28 08:30:14
非公有制企业党建(2016年1期)2016-07-19 13:02:52
开心素质教育(2016年2期)2016-04-20 05:07:48
