
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代胚胎发育及仔稚幼鱼形态观察
2016-09-08 04:52:56吴水清郑乐云黄种持仇登高许朝辉郑君飘邱峰岩吴精灵刘银华
渔业研究 2016年1期
吴水清,郑乐云*,黄种持,仇登高,许朝辉,郑君飘,邱峰岩,吴精灵,刘银华
(1.福建省水产研究所,福建省海洋生物增养殖与高值化利用重点实验室,福建 厦门 361013;2.厦门小嶝水产科技有限公司,福建 厦门 361000)
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代胚胎发育及仔稚幼鱼形态观察
吴水清1,郑乐云1*,黄种持1,仇登高1,许朝辉1,郑君飘2,邱峰岩2,吴精灵1,刘银华1
(1.福建省水产研究所,福建省海洋生物增养殖与高值化利用重点实验室,福建 厦门 361013;2.厦门小嶝水产科技有限公司,福建 厦门 361000)
对云纹石斑鱼(♀)与鞍带石斑鱼(♂)杂交子代胚胎及仔稚幼鱼的生长发育进行了观察与研究。在水温(22±1)℃、盐度33、pH7.8的海水中,杂交受精卵经过35h23min完成整个胚胎发育。胚胎发育可分为卵裂期、囊胚期、原肠期、神经胚期和器官形成期,其中在卵裂期、囊胚期和仔鱼出膜前后,杂交子代胚胎的死亡率较高。杂交石斑鱼的孵化率为91.5%,低于对照组的93.1%;畸形率为10.5%,高于对照组的5.3%。根据杂交子代卵黄囊消失与否、第二背鳍棘和腹鳍棘的生长与收缩、体色和鳞片的出现,可将杂交石斑鱼胎后发育分为:前期仔鱼(03日龄)、后期仔鱼(427日龄)、稚鱼期(2845日龄)、幼鱼期(46日龄以后);通过与其亲本早期发育进行比较发现,云纹石斑鱼(♀)与鞍带石斑鱼(♂)杂交子代具有一定的杂交优势。
云纹石斑鱼;鞍带石斑鱼;杂交子代;胚胎发育;形态观察
云纹石斑鱼(Epinephelus moara)和鞍带石斑鱼(E.lanceolatus)都隶属于鲈形目(Perciformes),鮨科(Serranidae)、石斑鱼亚科(Epinephelinae),石斑鱼属。云纹石斑鱼俗称草斑或真油斑,主要分布在日本、韩国和中国沿岸[1]。鞍带石斑鱼俗称龙趸、龙胆石斑鱼,是石斑鱼类群中体型最大的鱼类,主要产于东南亚、大洋洲海域,在中国的南海也可捕捞到,但是数量稀少[2-5]。云纹石斑鱼和鞍带石斑鱼都是肉食性鱼类,是具有较高营养价值的名贵海产鱼类,也是中国华南沿海地区重要的海水养殖种类之一[6-7]。
近年来杂交育种作为海水鱼类新品种选育的有效方法,也被广泛地用于石斑鱼新品种培育。目前国内外科研工作者已进行人工杂交的石斑鱼种类有:云纹石斑鱼(♀)×七带石斑鱼(E.septem faciatus)(♂)[8]、 斜带石斑鱼(E.coioides)(♀)×赤点石斑鱼(E.akaara)(♂)[9]、 巨石斑鱼(E.tauvina)(♀)×赤点石斑鱼(♂)[10]、 地中海石斑鱼(E.costae)(♀)×东大西洋石斑鱼(E.marginatus)(♂)[11]、 棕点石斑鱼(E.fuscoguttatus)(♀)×鞍带石斑鱼(♂)[12]、 斜带石斑鱼(♀)×鞍带石斑鱼(♂)[12-13]、 云纹石斑鱼(♀)×赤点石斑鱼(♂)[7,14]。
本研究对云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代早期胚胎发育及仔稚幼鱼发育过程中的形态变化特征进行较为系统的观察。旨在为石斑鱼新品种的选育提供参考,丰富杂交石斑鱼基础生物学文库,并为实现杂交石斑鱼规模化养殖提供科学依据。
1 材料与方法
1.1实验材料
实验于2015年4月至6月在厦门小嶝水产科技有限公司进行,实验所用的云纹石斑鱼和鞍带石斑鱼均取自经驯养的野生石斑鱼亲鱼,驯养时间为2~3年。亲鱼经注射促黄体激素释放激素(LHRH-A3)和人类绒毛膜促性腺激素(HCG)促熟、催产,对成熟的雌性云纹石斑鱼与雄性鞍带石斑鱼进行人工授精(干受精法),得到的受精卵(云纹石斑鱼♀×鞍带石斑鱼♂)作为实验组;对成熟的云纹石斑鱼进行人工授精(干受精法),所得受精卵(云纹石斑鱼♀×云纹石斑鱼♂)作为对照组。
1.2杂交受精卵的孵化及仔稚幼鱼的培育
将收集到的受精卵放入圆形孵化桶中孵化直至仔鱼孵出,期间要定时换水(8h换水一次,每次换水量为50%),孵化时水温为(22±1)℃,盐度33,pH7.8;待仔鱼孵出后,将仔鱼移入幼苗培育池中培育,水温(24±1)℃,盐度33,pH7.8。仔鱼开口后投喂牡蛎受精卵,连续投喂3d,之后开始投喂轮虫,17d后补充投喂卤虫无节幼体,20d后补充投喂桡足类和大卤虫,30d后以投喂桡足类和大卤虫为主,并根据各阶段幼体生长状况、存活数量和育苗池中生物饵料的存量,调整桡足类和大卤虫的投喂量和投饵次数,培育水体中每天补充小球藻。
1.3胚胎发育和仔稚幼鱼的形态观察
在孵化桶中每次随机取50cell受精卵,用NikonE200显微镜进行胚胎发育的连续观测和显微拍照,并详细记录受精卵发育过程的时间和发育特征,实验组和对照组均采用相同的操作方法。25日龄之前每天取样,26~50日龄每隔2天取样一次,50日龄之后每隔5天取样一次,每次均随机取样30尾。在NikonE200显微镜和OlympusSZX7解剖镜下观察杂交斑早期的形态发育,并拍照。
1.4数据处理
杂交斑胚胎和仔稚幼鱼的可量性状数据均用Image-Proexpress5.1 软件采集分析,所得数据进行单因素方差分析,数据以平均值±标准差(x±SD)表示。
2 结果
2.1胚胎发育
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交受精卵呈透明的球形(图版 Ⅰ-1),油球明显,卵径为(0.861±0.012)mm,油球径为(0.210±0.008)mm,在水温(22±1)℃、盐度33的海水中孵化,经过35h23min完成整个胚胎发育过程,早期胚胎发育各个阶段的形态如图版Ⅰ所示。胚胎发育可分为卵裂期、囊胚期、原肠期、神经胚期和器官形成期,胚胎发育时序如表1所示。

表1 云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代受精卵胚胎发育过程时序
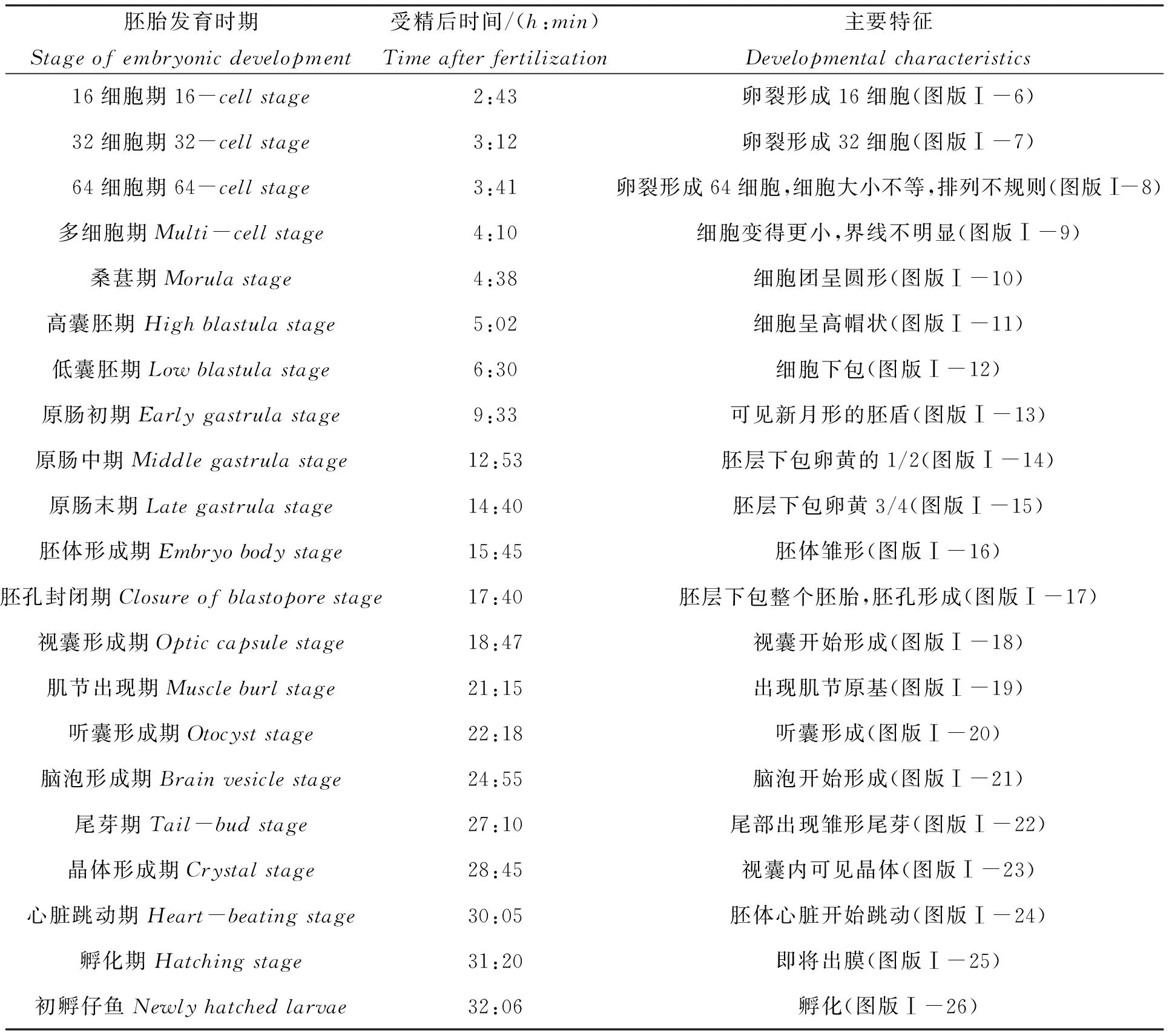
续表1
注:受精卵孵化温度(22±1)℃,盐度33。
Note:Hatchingtemperatureandsalinitywere(22±1)℃and33respectively.
2.1.1卵裂期
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交受精卵卵裂方式属于盘状卵裂,受精后32min可见胚盘隆起(图版Ⅰ-2);受精后1h1min开始第一次卵裂,此时受精卵一分为二,产生2个对等的细胞,进入2细胞期(图版Ⅰ-3);受精后1h35min,受精卵进行第二次分裂,进入4细胞期(图版Ⅰ-4);受精后2h5min开始第三次分裂,进入8细胞期(图版Ⅰ-5);受精后2h43min开始第四次分裂,进入16细胞期(图版Ⅰ-6);而后随着细胞的等分裂,细胞数量不断增加,经32细胞期(图版Ⅰ-7)、64细胞期,此时分裂的细胞大小不等,排列不规则(图版Ⅰ-8)和多细胞期(图版Ⅰ-9),受精4h38min后进入桑葚期(图版Ⅰ-10),此时的细胞体积变得更小,细胞间界限不明显,整个细胞团呈圆形。
2.1.2囊胚期
随着细胞分裂的进行,细胞的层数和数量不断增加,此时在胚盘与卵黄之间形成囊胚腔,囊胚中部呈高帽状突起,胚胎开始进入高囊胚期(图版Ⅰ-11);受精后6h30min,囊胚隆起开始变低,细胞逐渐下包,胚胎发育进入低囊胚期(图版Ⅰ-12)。
2.1.3原肠胚期
随着细胞分裂的进行,胚胎动物极细胞向植物极迁移和下包,受精后9h33min,胚层下包卵黄的四分之一,此时通过侧面观察可见一个新月形的胚盾,胚胎发育进入原肠初期(图版Ⅰ-13);受精后12h53min,胚层下包卵黄的二分之一,胚环位于胚胎中间位置,胚胎发育进入原肠中期(图版Ⅰ-14);受精后14h40min,胚层下包卵黄的四分之三,胚胎发育进入原肠末期(图版Ⅰ-15)。
2.1.4神经胚期
原肠期结束后,随着细胞分裂的进行,开始进入神经胚期,受精后15h45min,胚体形成神经板,其中央可见圆柱形脊索,胚体雏形,开始进入胚体形成期(图版Ⅰ-16);随后胚环逐渐下包收缩成小圆孔(即胚孔),受精后17h40min,胚层下包整个胚胎,胚孔闭合,此时胚胎发育进入胚孔封闭期(图版Ⅰ-17)。
2.1.5器官形成期
受精后18h47min,在胚体头部的前两侧分化出眼原基,形成视囊,胚体进入视囊形成期(图版Ⅰ-18);受精后21h15min,脊索两侧出现肌节原基,胚体进入肌节出现期(图版Ⅰ-19);受精后22h18min,胚体肌节变多,在视囊后方两侧出现1对听囊,胚体进入听囊形成期(图版Ⅰ-20);受精后24h55min,胚体的视囊之间出现脑泡,呈椭圆形板状,此时的脑泡尚未分室,胚体进入脑泡形成期(图版Ⅰ-21);受精后27h10min,胚体逐渐形成鳍褶,尾部逐渐与卵黄囊分离,胚体发育至尾芽期(图版Ⅰ-22);受精后28h45min,此时视囊内可见折光性强的透明状晶体,胚体抽动明显,胚体发育进入晶体形成期(图版Ⅰ-23);受精后30h5min,可见胚体心脏开始跳动,胚体进入心脏跳动期(图版Ⅰ-24);受精后31h20min,仔鱼孵化时胚体的头部先破膜伸出,尾部仍在膜内剧烈摆动(图版Ⅰ-25)。受精后32h06min,大部分的仔鱼孵出,仔鱼浮于水面,无游泳能力(图版Ⅰ-26)。
2.2杂交受精卵胚胎发育孵化率及畸形
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交受精卵呈透明的球形,油球明显,卵径为(0.861±0.012)mm,油球径为(0.210±0.008)mm。云纹石斑鱼的卵子和鞍带石斑鱼的精子具有较好的配子相容性,可受精产生正常发育的杂交子代。杂交组合的孵化率为91.5%,略低于对照组的93.1%,影响杂交受精卵孵化率的是在胚胎发育的卵裂期和囊胚期(图版Ⅰ-27)。杂交组合的畸形率为10.5%,也较云纹石斑鱼(♀)×云纹石斑鱼(♂)的受精卵高(畸形率5.3%),畸形主要分布在仔鱼出膜前后:(1)仔鱼在卵膜中就已经是畸形造成不能正常出膜(图版Ⅰ-28);(2)仔鱼出膜之后出现畸形不能正常发育(图版Ⅰ-29)。
2.3仔稚幼鱼形态发育
2.3.1前期仔鱼
仔鱼初孵至卵黄囊消失为前期仔鱼阶段。
初孵仔鱼:仔鱼浮于水面,无游泳能力,卵黄囊呈椭圆形,鳍褶起自头部后方,绕过尾部,终止于肛门后部,仔鱼脊索细长,身体透明,初孵仔鱼全长:(2.215±0.091)mm(图版Ⅰ-26)。
1日龄仔鱼:卵黄囊较前一天明显收缩(图版Ⅱ-1)。
2日龄仔鱼:卵黄囊即将消失,油球明显,仔鱼口裂形成,但未摄食,仍为内源性营养阶段(图版Ⅱ-2)。
3日龄仔鱼:眼部黑色素增加,视觉形成。鱼体出现2处黑色素沉积,一处在消化道中央上方位置,另一处在肛门与尾端之间,胸鳍呈叶状,游泳能力较之前明显增强,仔鱼口裂明显,消化道中还未见生物饵料(图版Ⅱ-3)。
2.3.2后期仔鱼
卵黄囊消失至第二背鳍棘和腹鳍棘达到仔稚鱼阶段的最大值。
4日龄仔鱼:消化系统的发育更加完善,腹部的黑色素几乎包围整个消化道,仔鱼已摄食,消化道中可见生物饵料,腹鳍棘开始出现,胸鳍进一步生长、呈透明的扇形,仔鱼开始有集群行为(图版Ⅱ-4)。
7日龄仔鱼:腹鳍棘明显,背鳍棘原基开始出现,鳃盖骨明显,胸鳍更加发达,活力更强,仔鱼游动速度加快,集群在池边和池角活动(图版Ⅱ-5)。
10日龄仔鱼:鱼体肌节呈“V”型,第二背鳍棘已明显可见,第二背鳍棘末端开始出现少量的黑色素,腹鳍棘进一步增长,腹鳍棘比第二背鳍棘长,尾部鳍膜开始出现黑色的鳍条丝(图版Ⅱ-6)。
15日龄仔鱼:第一背鳍棘出现,第二背鳍生长迅速,长度开始超过腹鳍棘,长棘仍呈锯齿状,尾柄中部的黑色素减少,尾椎开始向上弯曲,尾鳍呈辐射状扇形,肌节呈“W”型(图版Ⅱ-7)。
27日龄仔鱼:仔鱼各鳍基本形成,尾鳍已由圆形转变为截形尾,尾椎骨出现黑色素,第二背鳍棘和腹鳍棘长度达到仔稚鱼阶段的最大值,第二背鳍棘长度超过体长的一半,此后第二背鳍棘和腹鳍棘开始收缩(图版Ⅱ-8)。
2.3.3稚鱼期
第二背鳍棘和腹鳍棘开始收缩至稚幼鱼阶段的最小值,鳞片和体色开始出现。
30日龄稚鱼:第二背鳍棘和腹鳍棘收缩明显,鳍棘上面的小刺开始收回,鳍棘末端的黑色素消失,臀鳍明显,眼睛后缘出现黑色素群(图版Ⅱ-9)。
38日龄稚鱼:尾椎骨上的黑色素明显,第二背鳍棘和腹鳍棘较之前已明显收缩,但是第二背鳍棘的长度仍是鳍棘中最长的,鳞片在背鳍基部的两侧以及身体的背面开始出现(图版Ⅱ-10)。
2.3.4幼鱼期
体色与成鱼相似,全身覆盖鳞片。
46日龄幼鱼:幼鱼各鳍的形态与成鱼相似,幼鱼条带和体色明显,全身覆盖鳞片,第二背鳍棘和腹鳍棘收缩到稚幼鱼阶段最小值之后又开始生长(图版Ⅱ-11)。
3 讨论
3.1杂交子代与其亲本胚胎发育的比较
云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交受精卵的胚胎发育,与其亲本具有共同的特性,同时也存在差异:杂交受精卵的卵径为(0.861±0.012)mm,大于鞍带石斑鱼受精卵的0.790~0.850mm[15],与对照组云纹石斑鱼的受精卵接近;在相同孵化条件下,杂交受精卵与云纹石斑鱼受精卵两者所经历的各个时期及各时期的形态特征无明显差别,但是仔鱼初孵的时间比对照组(云纹石斑鱼♀×云纹石斑鱼♂)快1h;杂交受精卵的胚胎发育的大部分时期都晚于其父本鞍带石斑鱼的胚胎发育,但是初孵仔鱼全长(2.215±0.091)mm与云纹石斑鱼仔鱼全长接近,较鞍带石斑鱼仔鱼长(1.660±0.060)mm[2]。3.2杂交子代与其亲本仔稚幼鱼生长的比较
在石斑鱼早期发育阶段的划分中,大多数学者将背鳍棘、腹鳍棘和软鳍鳍条的发育程度作为各个阶段划分的标准。邹记兴等[16]将点带石斑鱼仔鱼的腹、背鳍棘开始收缩,鳞片长出前为后期仔鱼结束的标志。张海发等[17]将斜带石斑鱼各鳍基本形成、腹鳍棘和第二背鳍棘开始收缩以及鳍棘上小刺数目急剧减少、鳞片开始长出作为仔、稚鱼分界点,将腹鳍棘和第二背鳍棘绝对长度收缩到稚幼鱼阶段的最小值作为进入幼鱼阶段的标志。郭仁湘等[18]将鞍带石斑鱼各鳍基本形成作为进入稚鱼期的标志。综合以上学者的划分标准,可以将本研究27日龄作为结束后期仔鱼期进入稚鱼期的时间,46日龄进入幼鱼阶段。通过比较发现杂交子代具有其父本生长速度快的优势,杂交斑27日龄时第二背鳍棘和腹鳍棘长度达到最大值,各鳍已基本上发育完成,发育到稚鱼阶段的时间要晚于鞍带石斑鱼的22日龄(水温27~30℃)[18],但是进入稚、幼鱼阶段要明显早于云纹石斑鱼的31日龄和66日龄[水温(23±1)℃][19]。
3.3亲本组合的可行性
云纹石斑鱼和鞍带石斑鱼为同属不同种,染色体数目2n=48[20-21],这具备了杂交的细胞学基础。本研究属于种间的远缘杂交,该杂交组合的母本云纹石斑鱼与父本鞍带石斑鱼能产生正常分裂的受精卵,并能够发育到幼鱼阶段。杂交组合的孵化率为91.5%,略低于对照组的93.1%,影响杂交受精卵孵化率的时期主要是在卵裂期和囊胚期,但是杂交受精卵在神经胚期和器官形成期一般都能正常发育,造成这一现象的原因还有待进一步研究。杂交受精卵的畸形率为10.5%,要高于对照组的5.3%,畸形发生主要分布在仔鱼出膜前后:(1)仔鱼在卵膜中就已经是畸形造成不能正常出膜;(2)仔鱼出膜之后出现畸形不能正常发育。杨求华等[14]对云纹石斑鱼与赤点石斑鱼杂交的研究也表明,杂交受精卵的孵化率较云纹石斑鱼与赤点石斑鱼低,畸形率也高于对照组。
目前,对云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交育种的研究还未见报道,对杂交子代各方面特性了解还不够充分,有关杂交子代的低温耐受性、抗病力等方面是否优于亲本,还需进一步实验证实。杂交子代有生长发育速度较快的杂种优势,也有其不同于亲本的发育特点。本文探讨云纹石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代胚胎发育和仔、稚幼鱼生长发育,为杂交石斑鱼的鱼苗培育和人工养殖提供了科学参考。
[1]郭明兰,苏永全,陈晓峰,等.云纹石斑鱼与褐石斑鱼形态比较研究[J].海洋学报:中文版,2008,30(6):106-114.
[2]张海发,王云新,刘付永忠,等.鞍带石斑鱼人工繁殖及胚胎发育研究[J].广东海洋大学学报,2008,28(4):36-40.
[3]HeemstraPC,RandallJE.Groupersoftheworld[J].FAOFisheriesSynopsis,1993,16(125):130-132.
[4]SadovyY,CornishAS.ReeffishesofHongKong[M].HongKong:HongKongUniversityPress,2000.
[5]麦贤杰,黄伟健,叶富良,等.海水鱼类繁殖生物学和人工繁育[M].北京:海洋出版社,2005:166-177.
[6]王大鹏,曹占旺,谢达祥,等.石斑鱼的研究进展[J].南方农业学报,2012,43(7):1058-1065.
[7]郑乐云.云纹石斑鱼和赤点石斑鱼杂交子一代线粒体相关基因的母性遗传特征分析[J].上海海洋大学学报,2015,23(3):351-358.
[8]李炎璐,王清印,陈超,等.云纹石斑鱼(♀)×七带石斑鱼(♂)杂交子一代胚胎发育及仔稚幼鱼形态学观察[J].中国水产科学,2012,(5):821-832.
[9]刘付永忠,赵会宏,刘晓春,等.赤点石斑鱼♂与斜带石斑鱼♀杂交的初步研究[J].中山大学学报:自然科学版,2007,(3):72-75.
[10]宋盛宪,许波涛.石斑鱼杂交新品种“青红斑”获得成功[J].海洋渔业,1987,(6):271-272.
[11]GlamuzinaB,GlavicN,SkaramucaB,etal.EarlydevelopmentofthehybridEpinephelus costae♀× E.marginatus♂ [J].Aquaculture,2001,198(2):55-61.
[12]广东省大亚湾水产试验中心.青龙斑和虎龙斑[J].海洋与渔业,2010,(9):37.
[13]吴水清,李加儿,区又君,等.斜带石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代仔、稚鱼的异速生长[J].中国水产科学,2014,21(3):503-510.
[14]杨求华,黄种持,郑乐云,等.云纹石斑鱼(♀)×赤点石斑鱼(♂)杂交子代胚胎发育及生长[J].海洋渔业,2014,36(3):224-231.
[15]周玲,翁文明,李金亮,等.鞍带石斑鱼胚胎发育及仔鱼形态发育、饵料转变的观察研究[J].中国农学通报,2010,26(1):293-302.
[16]邹记兴,向文洲,胡超群,等.点带石斑鱼仔、稚、幼鱼的生长与发育[J].高技术通讯,2003,(4):77-84.
[17]张海发,刘晓春,刘付永忠,等.斜带石斑鱼胚胎及仔稚幼鱼形态发育[J].中国水产科学,2006,(5):689-699.
[18]郭仁湘,符书源,杨薇,等.鞍带石斑鱼仔稚(幼)鱼的发育和生长研究[J].水产养殖,2011,(4):8-13.
[19]宋振鑫,陈超,瞿介明,等.云纹石斑鱼胚胎发育及仔、稚、幼形观察[J].渔业科学进展,2012,33(3):26-34.
[20]舒琥,魏秋兰,罗丽娟,等.广东沿海4种石斑鱼的染色体组型分析[J].广东农业科学,2012,(8):124-127.
[21]郭丰,王军,苏永全,等.云纹石斑鱼染色体核型研究[J].海洋科学,2006,30(8):1-3.


1.受精卵;2.胎盘隆起;3.2细胞期;4.4细胞期;5.8细胞期;6.16细胞期;7.32细胞期;8.64细胞期;9.多细胞期;10.桑葚期;11.高囊胚期;12.低囊胚期;13.原肠初期;14.原肠中期;15.原肠末期;16.胚体形成期;17.胚孔封闭期;18.视囊形成期;19.肌节出现期;20.听囊形成期;21.脑泡形成期;22.尾芽期;23.晶体形成期;24.心脏跳动期;25.孵化期;26.初孵仔鱼;27.死亡的囊胚期胚胎;28.畸形仔鱼;29.未能出膜的仔鱼。
1.Fertilizedegg;2.Blastodiscformation;3.2-cellstage;4.4-cellstage;5.8-cellstage;6.16-cellstage;7.32-cellstage;8.64-cellstage;9.Multi-cellstage;10.Morulastage;11.Highblastulastage;12.Lowblastulastage;13.Earlygastrulastage;14.Middlegastrulastage;15.Lategastrulastage;16.Embryobodystage;17.Closureofblastoporestage;18.Opticcapsulestage;19.Muscleburlstage;20.Otocyststage;21.Brainvesiclestage;22.Tail-budstage;23.Crystalstage;24.Heart-beatingstage;25.Hatchingstage;26.Newlyhatchedlarvae;27.Embryostoppedatblastulastage;28.Abnormallarvae;29.Deformedembryo.

1.1d仔鱼;2.2d仔鱼;3.3d仔鱼;4.4d仔鱼;5.7d仔鱼;6.10d仔鱼;7.15d仔鱼;8.27d仔鱼;9.30d稚鱼;10.38d稚鱼;11.46d幼鱼。
1.Larva1dafterhatching(DAH);2.2DAH;3.3DAH;4.4DAH;5.7DAH;6.10DAH.7.15DAH;8.27DAH;9.30djuvenile;10.38djuvenile;11.46dyoungfish.
Embryonic and morphological development in larva,juvenile,and young stages of hybrid grouper (Epinephelus moara ♀×E.lanceolatus ♂)
WU Shuiqing1,ZHENG Leyun1*,HUANG Zhongchi1,QIU Denggao1,XU Chaohui1,ZHENG Junpiao2,QIU Fengyan2,WU Jingling1,LIU Yinhua1
(1.KeyLaboratoryofCultivationandHigh-valueUtilizationofMarineOrganismsinFujianProvince,FisheriesResearchInstituteofFujian,Xiamen361013,China2.XiamenXiaoDengAquaticScienceandTechnologyCo.,Ltd.,Xiamen361013,China)
Hybridizationisthemostwidelyusedandeffectivemethodofartificialbreedinginaquaculture.Wedocumentedembryonicdevelopmentandthegrowthandmorphologicalcharacteristicsoflarva,juveniles,andyoungfishinthehybrid(Epinephelus moara ♀× E.lanceolatus ♂)byartificialmethods.Theembryonicdevelopmentofthehybridwasdividedintocleavagestage,blastulastage,gastrulastage,neurulastage,andorganogenesisstage.Inseawater[(22±1)℃,salinity33,pH7.8],35h23minwasrequiredforhybridembryonicdevelopment.Incleavagestage,blastulastage,beforeandafterthelarvae,thehybridhadhighermortality.Statisticsshowedthathybridhatchingratewas91.5%lowerthancontrolgroup(93.1%),deformityratewas10.5%higherthancontrolgroup(5.3%).Basedonthefeaturesoftheyolksac,seconddorsalfinspine,pelvicfinspine,scalesandbodycolor,thepost-embryonicdevelopmentofthehybridwasdividedintolarval(0~27d),juvenile(28~45d),andyoungfishstages(46d).TheresultssuggestedthatthehybridgrowsmorerapidlythanE.moara.
Epinephelus moara;E.lanceolatus;hybrid;embryonicdevelopment;morphologicalobservation
2015-12-30
厦门南方海洋研究中心项目(14PZY017NF17).
吴水清(1987-),男,实习研究员,硕士,主要从事海水鱼类繁育生物学.E-mail:wushuiqing@163.com
郑乐云(1969-),男,教授级高级工程师,主要从事海水鱼类繁养殖及育种.E-mail:lyzheng69@sina.com
S961.5
A
1006-5601(2016)01-0027-09

猜你喜欢
当代水产(2023年2期)2023-05-19 08:23:02
渔业研究(2022年5期)2022-10-28 01:56:14
当代水产(2020年3期)2020-06-15 12:02:22
当代水产(2019年7期)2019-09-03 01:01:56
当代水产(2019年5期)2019-07-25 07:51:06
传播力研究(2018年27期)2018-03-28 10:20:49
广东海洋大学学报(2015年4期)2016-01-13 08:39:40
纺织科技进展(2015年1期)2015-11-28 05:56:36
应用海洋学学报(2015年3期)2015-11-22 07:39:22
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:34
