
胡萝卜肉桂醇脱氢酶基因的克隆及其对非生物胁迫的响应
2016-09-02 02:41刘堰珺王广龙熊爱生
西北植物学报 2016年7期
刘堰珺,马 静,王广龙,黄 蔚,王 枫,熊爱生
胡萝卜肉桂醇脱氢酶基因的克隆及其对非生物胁迫的响应
刘堰珺,马静,王广龙,黄蔚,王枫,熊爱生*
(南京农业大学作物遗传与种质创新国家重点实验室,园艺学院,农业部华东地区园艺作物生物学与种质创制重点实验室,南京210095)
该实验采用RT-PCR技术,从胡萝卜‘黑田五寸’中克隆获得了编码肉桂醇脱氢酶(Cinnamyl alcohol dehydrogenase, CAD)的基因DcCAD。DcCAD序列长1 074 bp,编码357个氨基酸。亲水/疏水性分析表明,DcCAD属于亲水性蛋白。系统进化分析显示,DcCAD与番茄CAD(XP_010314515.1)亲缘关系最近。荧光定量PCR结果显示,DcCAD基因在胡萝卜根、叶片和叶柄中的表达量差异显著,其相对表达量为叶片>根>叶柄。DcCAD基因对高温(38 ℃)、低温(4 ℃)、干旱(20% PEG)和盐(0.2 mol·L-1NaCl)胁迫均有响应,尤其对高温胁迫和低温胁迫响应明显,而且高温处理后1 h和低温处理后2 h表达量最高。研究推测,DcCAD基因对胡萝卜抗逆性具有一定的作用。
肉桂醇脱氢酶;同源分析;非生物胁迫;表达分析;胡萝卜
木质素(lignin)主要分布于木质化的次生细胞壁中,它为植物细胞壁提供了必要的强度、疏水性以及对外界恶劣环境的抗性[1-3]。木质素的生物合成是一个由多种酶参与的复杂调控网络,肉桂醇脱氢酶(cinnamyl alcohol dehydrogenase, CAD)作为其中的关键酶,人们在很早之前就开始对其进行研究[4]。国内外已对植物中肉桂醇脱氢酶的结构性质、相应的功能以及转基因方面进行了一些研究[5]。
由于肉桂醇脱氢酶对木质素单体的生物合成起着重要的作用,目前,已有多个编码肉桂醇脱氢酶的基因从不同植物中被成功克隆,如火炬松(Pinustaeda)[6]、九眼独活(Araliacordata)[7]、紫花苜蓿(Medicagosativa)[8-9]、多年生黑麦草(Loliumperenne)[10]和高粱(Sorghumbicolor)[11]等。研究也证实,调控木质素合成途径中相关基因的表达,可以控制木质素的生物合成。通过分析杨树(Populuslaurifolia)[12]、小麦(Triticumaestivum)[13]、二穗短柄草(Brachypodiumdistachyon)[14]中肉桂醇脱氢酶基因的表达模式发现,肉桂醇脱氢酶基因在植物中以多基因家族的形式存在。植物在受到各种生物及非生物胁迫时,其木质素代谢相关基因的表达会发生变化,其木质素含量也会相应改变。
胡萝卜(DaucuscarotaL.)是伞形科胡萝卜属的一种重要蔬菜,在世界各地广泛栽培。胡萝卜以肉质根为食用部位,含有丰富的营养物质和水溶性纤维[15]。作为根用型蔬菜,胡萝卜肉质根中木质素含量的增加会影响胡萝卜的品质。目前,有关伞形科蔬菜作物中木质素代谢相关研究报道很少。
本研究以胡萝卜‘黑田五寸’为实验材料,基于转录组和基因组数据,通过RT- PCR方法克隆出木质素合成关键酶—肉桂醇脱氢酶的基因(DcCAD),并对其进行了较为详尽的分析。通过实时定量PCR技术,研究DcCAD基因在胡萝卜不同组织及不同逆境处理下的表达情况,为进一步深入研究胡萝卜中肉桂醇脱氢酶的功能及应用奠定基础。
1 材料和方法
1.1材料
胡萝卜‘黑田五寸’于2015年10月份种植于南京农业大学作物遗传与种质创新国家重点实验室人工气候室。胡萝卜植株种植在蛭石和有机质1∶1的混合基质中,光照和温度设定为:14 h 25 ℃光照以及10 h 18 ℃黑暗,相对湿度60%~70%以及240 μmol·m-2·s-1光照强度。
大肠杆菌菌株DH5α由本实验室保存;质粒载体pMD19-T vector simple、ExTaq酶、反转录酶MluI、DL marker 2000和dNTP荧光定量染料SYBR Green I等均购自大连TaKaRa公司。
1.2方法
1.2.1取样对‘黑田五寸’2月龄植株分别进行高温(38 ℃)、低温(4 ℃)、干旱(20% PEG)和盐(0.2 mol·L-1NaCl)处理,分别取处理 0、1、2、4、8和24 h的成熟健康叶片,同时取未处理植株叶片为对照。取2月龄‘黑田五寸’植株,剪取其生长健康的叶片用于叶片总RNA提取,反转录成cDNA,用于克隆DcCAD基因。取2月龄‘黑田五寸’植株的叶片、叶柄、根为材料进行组织表达分析。每次取样3个重复,取样后立即用液氮速冻,并保存于-80 ℃冰箱。
1.2.2RNA提取及cDNA合成用RNA simple Total Kit(Tiangen公司,中国北京)提取总RNA,RNA浓度和质量利用Nanodrop ND-100分光光度计(NanoDrop公司,美国)检测。利用Prime Script RT reagent Ki(TaKaRa公司,中国大连)将提取的总RNA反转录成cDNA。所有cDNA样品稀释10倍用于PCR。
1.2.3胡萝卜DcCAD基因序列的获得与克隆基于本实验室测定的胡萝卜转录组数据和基因组数据[16],拼接得到DcCAD基因的序列,并设计正向引物DcCAD-F(5′-ATGGCTCAAACAACTCCCAATCACA-3′)和反向引物DcCAD-R(5′-CTATGGTTTTCCAGCAATATCAAT-3′)。以2月龄‘黑田五寸’新鲜主根cDNA第一链为模板进行扩增,反应条件为94 ℃预变性4 min;94 ℃变性30 s;54 ℃退火30 s;72 ℃延伸1 min;35个循环;72 ℃延伸10 min。利用1.2%琼脂糖凝胶电泳分离扩增产物,回收后连接到pMD19-T simple载体上,并转化到大肠杆菌DH5α。随后提取质粒进行PCR鉴定,委托南京金斯瑞生物科技有限公司进行DNA测序。
1.2.4序列分析不同植物相关基因和氨基酸序列均来自NCBI数据库;利用BioXM 2.6软件对DcCAD基因的可阅读框长度进行预测,并翻译成对应的氨基酸序列。通过DNAMAN进行氨基酸序列比对及亲水性/疏水性分析,然后用Mega 5构建进化树和编辑并生成报告图形[17]。蛋白质基本性质分析使用ExPASy相关软件完成[18]。通过SWISS-MODEL(http://swiss-model.expase.org)预测DcCAD蛋白的三维结构。
1.2.5实时定量PCR反应实时定量PCR采用SYBR PremixExTaq试剂盒(大连TaKaRa公司),在ABI 7300荧光定量PCR仪上完成,采用iQTM5 Real-time PCR System完成,按照操作说明进行。相对定量方法参照2-ΔΔCt法。以胡萝卜Tubulin为参照基因[19],引物为DcA15-F(5′-GAGTGGAGTTACCTGCTGCCTTC-3′)和DcA15-R(5′-ATGTAGACGAGGGAACGGAATCCAG-3′)。DcCAD基因表达分析引物为DcCAD-F(5′-CGGATAAGCCTCTGGACCTACCT-3′)和DcCAD-R(5′-TTGACAACTTCGATACCACACCTGAT-3′),目标基因与Tubulin一起扩增。实验进行3次生物学重复,对数值进行了差异显著水平分析。
1.2.6数据分析采用 Excel 2003和SPSS 16.0进行数据整理和基因表达分析,采用Duncan测验分析数据差异显著性。
2 结果与分析
2.1胡萝卜DcCAD基因克隆
PCR结果(图1)显示,以胡萝卜‘黑田五寸’的cDNA为模版扩增得到1 000 bp左右产物,其长度与
预期相一致。测序结果显示,DcCAD基因长1 074 bp, 编码357个氨基酸(图2),预测其蛋白质相对分子质量为39.09 kD,等电点6.7。
2.2氨基酸序列比对和理化性质分析
对DcCAD的氨基酸序列进行BLASTp同源性检索,结果(图)表明:DcCAD的氨基酸序列与拟南芥(Arabidopsisthaliana,NP_195643.1)、黄蒿(Artemisiaannua,ACB54931.1)、茶树(Camelliasinensis,ACV74414.1)、辣椒(Capsicumannuum,ACF17645.1)、黄瓜(Cucumissativus,XP_004136373.1)、黑麦草(Loliumperenne,AAL99536.1)、苹果(Malusdomestica,XP_008359757.1)、烟草(Nicotianaattenuata,AFP43763.1)、马铃薯(Solanumtuberosum,XP_006356896.1)、小麦(Triticumaestivum,CDM85988.1)、玉米(Zeamays,NP_001136687.1)、水稻(Oryzasativa,NP_001052290.1)、高粱(Sorghumbicolor,XP_002446076.1)、菠菜(Spinaciaoleracea,KNA09004.1)和蓖麻(Ricinuscommunis,XP_011083448.1)等植物的氨基酸序列具有较高的相似度,相似性均在60%以上(图3)。
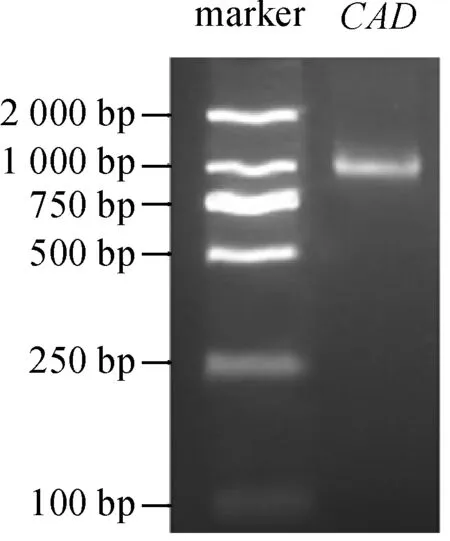
图1 胡萝卜DcCAD基因克隆图谱Fig. 1 PCR amplification of DcCAD gene from carrot

图2 胡萝卜DcCAD基因及其编码氨基酸序列Fig. 2 The nucleotide acid and deduced amino acid sequences of DcCAD gene from carrot

星号表示3个催化锌结合位点;箭头表示3个锌结合位点; CAD蛋白登录号依次为:蓖麻(XP_011083448.1)、菠菜(KNA09004.1)、茶树(ACV74414.1)、高粱(XP_002446076.1)、黑麦草(AAL99536.1)、胡萝卜(ALN38479.1)、黄瓜(XP_004136373.1)、黄蒿(ACB54931.1)、辣椒(ACF17645.1)、马铃薯(XP_006356896.1)、拟南芥(NP_195643.1)、苹果(XP_008359757.1)、水稻(NP_001052290.1)、小麦(CDM85988.1)、烟草(AFP43763.1)和玉米(NP_001136687.1)图3 胡萝卜DcCAD的氨基酸序列与其他物种类似蛋白质氨基酸序列的多重比对The asterisk marks the 3 catalytic Zn binding sites; The arrow marks the 3 structural Zn binding sites; The accession numbers of CAD from different plants were: Ricinus communis (XP_011083448.1), Spinacia oleracea (KNA09004.1), Camellia sinensis(ACV74414.1), Sorghum bicolor (XP_002446076.1), Lolium perenne (AAL99536.1), Daucus carota(ALN38479.1), Cucumis sativus (XP_004136373.1), Artemisia annua (ACB54931.1), Capsicum annuum (ACF17645.1), Solanum tuberosum (XP_006356896.1), Arabidopsis thaliana (NP_195643.1), Malus domestica (XP_008359757.1), Oryza sativa (NP_001052290.1), Triticum aestivum (CDM85988.1), Nicotiana attenuata (AFP43763.1), and Zea mays (NP_001136687.1)Fig. 3 Alignment of amino acid sequences of carrot DcCAD and other different plants
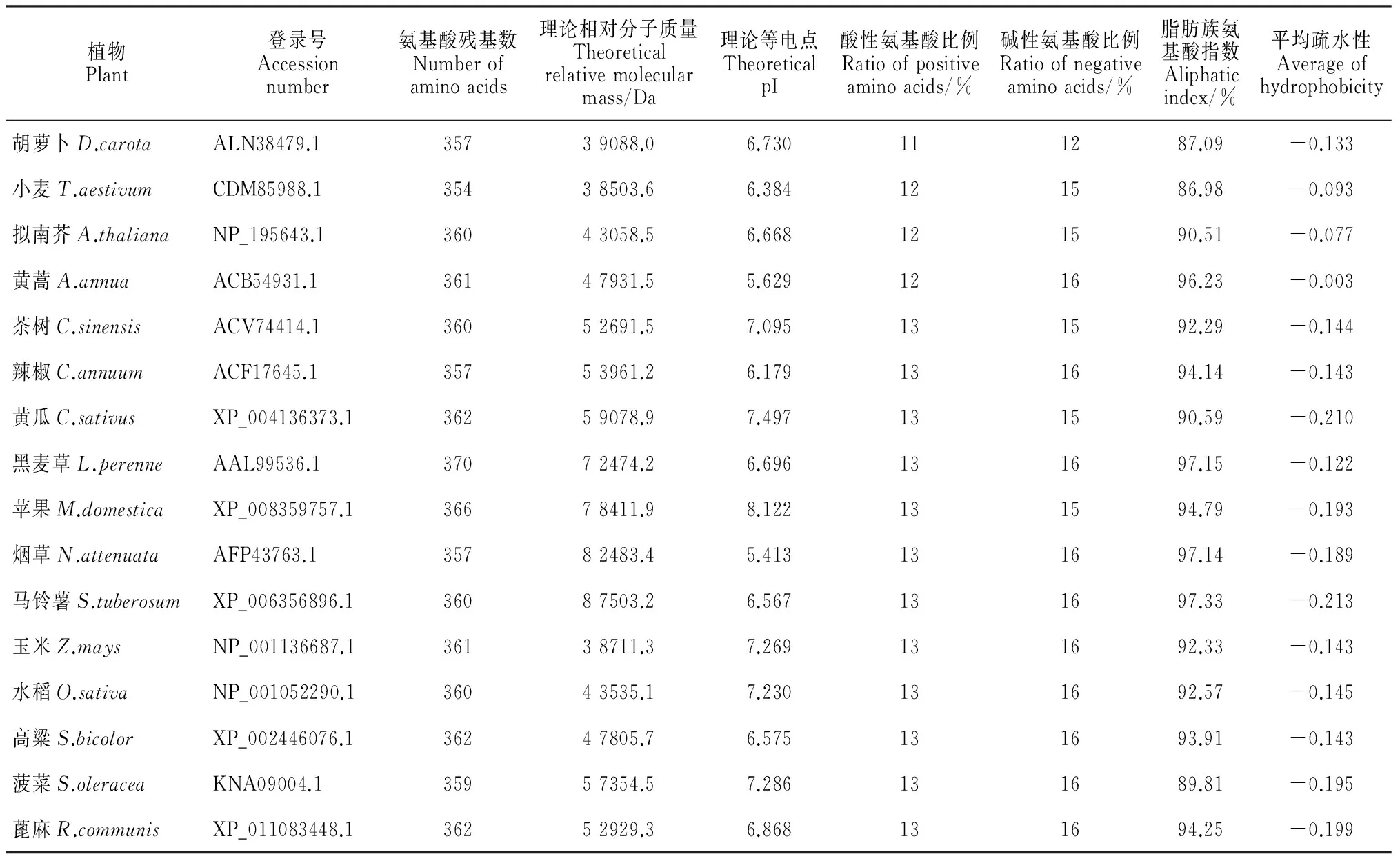
植物Plant登录号Accessionnumber氨基酸残基数Numberofaminoacids理论相对分子质量Theoreticalrelativemolecularmass/Da理论等电点TheoreticalpI酸性氨基酸比例Ratioofpositiveaminoacids/%碱性氨基酸比例Ratioofnegativeaminoacids/%脂肪族氨基酸指数Aliphaticindex/%平均疏水性Averageofhydrophobicity胡萝卜D.carotaALN38479.135739088.06.730111287.09-0.133小麦T.aestivumCDM85988.135438503.66.384121586.98-0.093拟南芥A.thalianaNP_195643.136043058.56.668121590.51-0.077黄蒿A.annuaACB54931.136147931.55.629121696.23-0.003茶树C.sinensisACV74414.136052691.57.095131592.29-0.144辣椒C.annuumACF17645.135753961.26.179131694.14-0.143黄瓜C.sativusXP_004136373.136259078.97.497131590.59-0.210黑麦草L.perenneAAL99536.137072474.26.696131697.15-0.122苹果M.domesticaXP_008359757.136678411.98.122131594.79-0.193烟草N.attenuataAFP43763.135782483.45.413131697.14-0.189马铃薯S.tuberosumXP_006356896.136087503.26.567131697.33-0.213玉米Z.maysNP_001136687.136138711.37.269131692.33-0.143水稻O.sativaNP_001052290.136043535.17.230131692.57-0.145高粱S.bicolorXP_002446076.136247805.76.575131693.91-0.143菠菜S.oleraceaKNA09004.135957354.57.286131689.81-0.195蓖麻R.communisXP_011083448.136252929.36.868131694.25-0.199
对上述植物进行氨基酸组成成分及理化性质分析,结果(表1)发现,各植物氨基酸残基数在354~370之间不等;理论相对分子质量的差异较大,理论等电点在5.413~8.122之间;碱性氨基酸所占比例略高于酸性氨基酸,脂肪族氨基酸的比例在86.98%~97.33%之间;蛋白质可溶性预测中平均疏水性为-0.213~-0.003,其中黄蒿的疏水性最强,马铃薯的疏水性最弱。
2.3氨基酸亲水性/疏水性分析及结构域分析
对DcCAD氨基酸序列进行疏水性/亲水性分析,结果(图4)表明,DcCAD疏水区域中,第187~202位疏水性较强,第165位的丙氨酸(Ala)和第189位、276位的缬氨酸(Val)疏水性最强。亲水区域中182~187位亲水性较强,第33位的天冬酰胺(Asn)、112位的谷氨酸(Glu)和223位的赖氨酸(Lys)疏水性最强。结果表明,DcCAD属于亲水性蛋白。对胡萝卜DcCAD编码的氨基酸序列进行Blastp同源性检索,结果显示胡萝卜DcCAD属于MDR超家族,归为CAD1家族,具有多个Zn结合位点,属于多结构域蛋白序列。
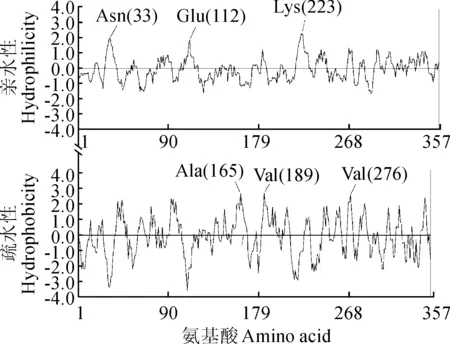
图4 胡萝卜DcCAD氨基酸序列的亲水性和疏水性Fig. 4 Predicted hydrophilicity and hydrophobicty of deduced amino acid sequence of DcCAD from carrot
2.4蛋白三级结构预测和进化分析
胡萝卜DcCAD进行蛋白质三级结构同源建模分析的结果如图5所示。胡萝卜DcCAD蛋白有9个α螺旋和16个β折叠,与拟南芥的CAD蛋白有较高的相似性。通过Blastp检索,将DcCAD的氨基酸序列与GenBank已报道的其他植物的CAD蛋白序列进行比对,用Mega 5构建系统进化树(图6)。

图6 胡萝卜DcCAD与其他植物CAD的系统进化树Fig. 6 Phylogenetic tree of DcCAD from carrot and CAD from other species
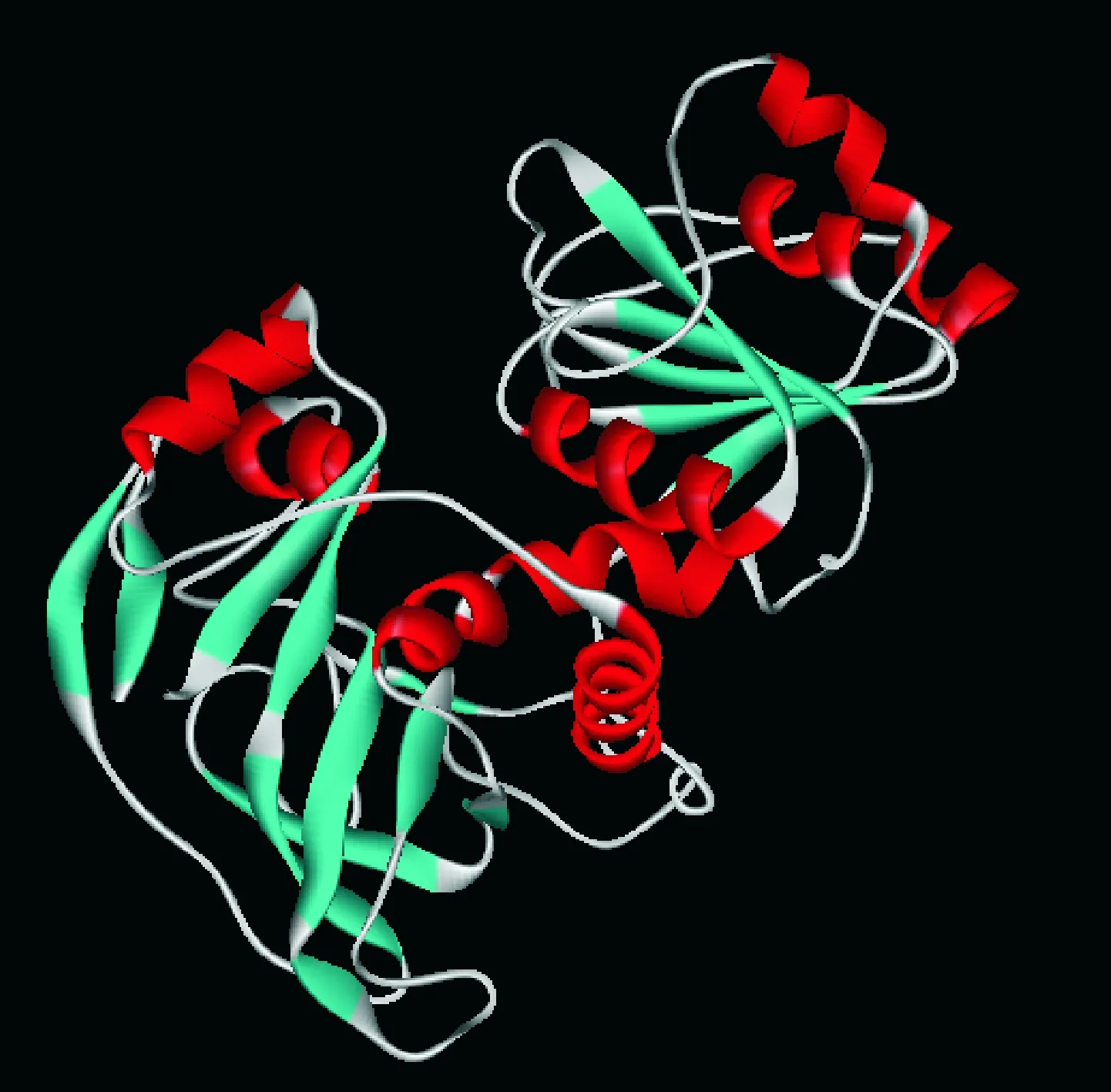
图5 胡萝卜DcCAD三级结构Fig. 5 The three-dimension structural model of DcCAD from carrot
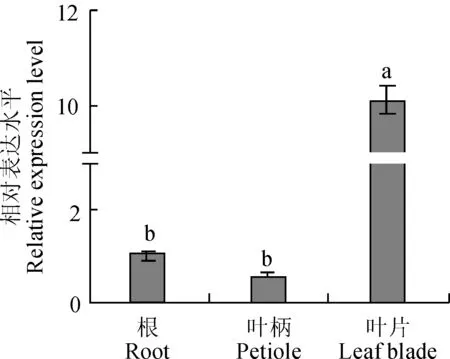
图中小写字母表示0.05水平差异显著性图7 胡萝卜DcCAD基因在不同组织中的表达情况The different letters mean significant difference at 0.05 levelFig. 7 Expression analysis of the DcCAD gene in different tissues of carrot
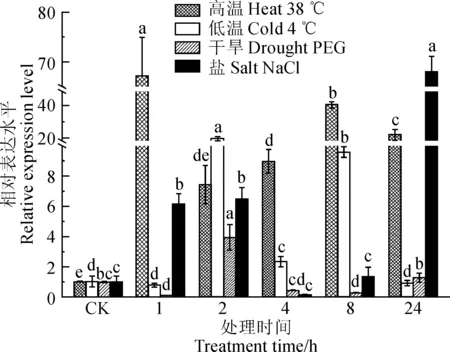
竖条上方不同字母代表同一处理在0.05水平上的差异显著性图8 胡萝卜DcCAD基因在不同逆境以及不同时间段处理的表达Different letters on the vertical bars indicate significant difference in the same treatment at 0.05 levelFig. 8 Expression profiles of the DcCAD gene under different abiotic stress treatments in carrot
结果表明:胡萝卜与茄科的番茄亲缘关系最近,两者氨基酸相似性达78.89%。与菠菜、茶树、葡萄CAD进化关系较近,与蔷薇科的苹果、胡麻科的芝麻、山茶科的蓖麻以及杨柳科的杨树和毛果杨CAD进化关系较远,与葫芦科的黄瓜、甜瓜以及桉树CAD进化关系最远。
2.5胡萝卜DcCAD基因在不同组织及不同非生物胁迫条件下的表达
以胡萝卜‘黑田五寸’的根、叶柄和叶片为材料,通过荧光定量PCR检测DcCAD基因在胡萝卜不同组织中的表达情况。结果(图7)显示,DcCAD基因在胡萝卜不同组织中的表达量不同,且差异较大。叶片中表达量高于叶柄和根,叶片中表达量分别是根的10倍、叶柄的17.7倍。
用荧光定量PCR方法测定胡萝卜‘黑田五寸’在高温(38 ℃)、低温(4 ℃)、干旱(20% PEG)和盐(0.2 mol·L-1NaCl)4种胁迫处理后的表达情况。结果(图8)显示,38 ℃处理下,1、2、4、8和24 h表达量均较高,分别是未处理对照的65.8、7.28、8.79、39.66和21.47倍。4 ℃处理下随着处理时间的延长,DcCAD基因的表达量呈现低-高-低的趋势,且处理2 h后表达量最高,是对照的18.75倍。20% PEG处理下,除2 h和24 h外,其余处理时间的表达低于对照,2 h表达量达到峰值,是对照的3.95倍。盐处理下,DcCAD基因总体表现为先上升,后下降,再上升,并于24 h达到峰值。
3 讨 论
肉桂醇脱氢酶在植物木质素生物合成中发挥着重要的作用,其活性的高低影响着植物木质素的含量和木质素单体的组成[20-21]。目前已经从多种植物中分离编码肉桂醇脱氢酶基因,并通过转基因方法对其功能进行了验证。本研究以‘黑田五寸’胡萝卜为研究对象,克隆获得编码肉桂醇脱氢酶基因DcCAD。氨基酸理化性质分析结果显示,DcCAD属亲水性蛋白,但不同物种间的氨基酸理化性质存在一定差异,这可能与其参与不同的逆境调控有关[22]。通过对胡萝卜DcCAD基因及编码蛋白的同源性分析发现,与DcCAD相似性最高的是番茄CAD。DcCAD与葡萄科、山茶科、藜科的CAD相似性较高,与单子叶禾本科、葫芦科、杨柳科的CAD亲缘关系较远。植株中的CAD在不同科之间保守性并不高,DcCAD与其他物种同源基因的相似性只有60 %左右。本研究通过对不同物种CAD进化关系进行分析,为探讨植物中CAD起源和进化提供一定的借鉴。
植物对外界环境变化的调控网络十分复杂,当处于逆境条件下时,各组织器官会行使其特定的生理生化功能,产生相应的应对机制[23-25]。木质化过程也是植物响应逆境胁迫的一种方式,它可能被臭氧、重金属、干旱、紫外线、机械损伤、病虫害等逆境所诱导[26-29]。肉桂醇脱氢酶通过将醛类转化为相应的醇类来参与木质素的生物合成,同时它还与植物的逆境应答有关[26,30-31]。当植株受到外界环境的伤害时,会合成木质素来响应逆境胁迫。本研究中对胡萝卜植株进行高温、低温、干旱、盐胁迫处理,发现DcCAD基因对非生物胁迫响应明显。此外DcCAD基因在胡萝卜不同组织中的表达量不同,在叶片中的表达量最高,叶柄中的表达量最低。这些结果说明胡萝卜DcCAD基因对逆境胁迫有响应,由此可推测它对胡萝卜的抗逆性起着一定的作用。
[1]PETER G, NEALE D. Molecular basis for the evolution of xylem lignification[J].CurrentOpinioninPlantBiology, 2004, 7(6): 737-742.
[2]GUO D M, RAN J H, WANG X Q. Evolution of the cinnamyl/sinapyl alcohol dehydrogenase (CAD/SAD) gene family: The emergence of real lignin is associated with the origin of bona fide CAD[J].JournalofMolecularEvolution, 2010, 71(3): 202-218.
[3]WENG J K, CHAPPLE C. The origin and evolution of lignin biosynthesis[J].NewPhytologist, 2010, 187(2):273-285.
[4]张鲁斌, 谷会, 弓德强, 等. 植物肉桂醇脱氢酶及其基因研究进展[J].西北植物学报, 2011, 31(1):204-211.
ZHANG L B, GU H, GONG D Q,etal. Research progress of cinnamyl alcohol dehydrogenase and its gene[J].ActaBotanicaBoreali-OccidentaliaSinica, 2011, 31(1):204-211.
[5]曹佳强, 李波, 杨洋, 等. 木质素生物合成中肉桂醇脱氢酶基因(CAD)的研究进展[J].分子植物育种, 2014,10(5):1 034-1 043.
CAO J Q, LI B, YANG Y,etal. Advanced progress in the cinnamyl alcohol dehydrogenase gene involved in lignin biosynthesis[J].MolecularPlantBreeding, 2014, 10(5):1 034-1 043.
[6]O’MALLEY D M, PORTER S, SEDEROFF R R. Purification, characterization, and cloning of cinnamyl alcohol dehydrogenase in loblolly pine (PinustaedaL.)[J].PlantPhysiology, 1992, 98(4), 1 364-1 371.
[7]HIBINO T, SHIBATE D, CHEN J Q,etal. Cinnamyl alcohol dehydrogenase fromAraliacordata: cloning of the cDNA and expression of the gene in lignified tissues[J].PlantCellPhysiology, 1993, 34(5): 659-665.
[8]BRILL E M, ABRAHAMS S, HAYES C M,etal. Molecular characterization and expression of a wound-inducible cDNA encoding a novel cinnamyl-alcohol dehydrogenase enzyme in lucerne (MedicagosativaL.)[J].PlantMolecularBiology, 1999, 41(2): 279-291.
[9]VAN DOORSSELAERE J, BAUCHER M, FEUILLET C,etal. Isolation of cinnamyl alcohol dehydrogenase cDNAs from two important economic species: alfalfa and poplar. Demonstration of a high homology of the gene within angiosperms[J].PlantPhysiologyandBiochemistry, 1995, 33(1): 105-109.
[10]LYNCH D, LIDGETT A, MCINNES R,etal. Isolation and characterisation of three cinnamyl alcohol dehydrogenase homologue cDNAs from perennial ryegrass (LoliumperenneL.)[J].JournalofPlantPhysiology, 2002, 159(6):653-660.
[11]TSURUTA S, EBINA M, NAKAGAWA H,etal. Isolation and characterization of cDNA encoding cinnamyl alcohol dehydrogenase (CAD) in sorghum (Sorghumbicolor(L.) Moench)[J].JapaneseSocietyofGrasslandScience, 2007, 53(2): 103-109.
[12]EUDES A, GEORGE A, MUKERJEE P,etal. Biosynthesis and incorporation of side-chain-truncated lignin monomers to reduce lignin polymerization and enhance saccharification[J].PlantBiotechnologyJournal, 2010, 10(5): 609-620.
[13]MA Q H. Functional analysis of a cinnamyl alcohol dehydrogenase involved in lignin biosynthesis in wheat[J].JournalofExperimentalBotany, 2012, 61(10): 2 735-2 744.
[14]BUKH C, NORD-LARSEN P H, RASMUSSEN S K. Phylogeny and structure of the cinnamyl alcohol dehydrogenase gene family inBrachypodiumdistachyon[J].JournalofExperimentalBotany, 2012, 63(17): 6 223-6 236.
[15]庄飞云, 欧承刚, 赵志伟, 等. 胡萝卜育种回顾及展望[J].中国蔬菜, 2008, 15(3): 41-44.
ZHUANG F Y, OU C G, ZHAO Z W,etal. Review and prospect of carrot breeding[J].ChinaVegetables, 2008, 15(3):41-44.
[16]XU Z S, TAN H W, WANG F,etal. CarrotDB: a genomic and transcriptomic database for carrot[J].DatabasetheJournalofBiologicalDatabases&Curation, 2014, 40 (5):1-8.
[17]TAMURA K, PETERSON D, PETERSON N,etal. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J].MolecularBiologyandEvolution, 2011, 28(10): 2 731-2 739.
[18]GASTEIGER E, GATTIKER A, HOOGLAND C,etal. ExPASy: the proteomics server for in-depth protein knowledge and analysis[J].NucleicAcidsResearch, 2003, 31(13): 3 784-3 788.
[19]TIAN C, JIANG Q, WANG F,etal. Selection of suitable reference genes for qPCR normalization under abiotic stresses and hormone stimuli in carrot leaves[J].PLOSOne, 2015, 10(2):128-136.
[20]PILATE G, GUINEY E, HOLT K. Field and pulping performances of transgenic trees with altered lignification[J].NatureBiotechnology,2002, 20(4):607-612.
[21]CHABANNES M, BARAKATE A, LAPIERRE C,etal. Strong decrease in lignin content without significant alteration of plant development is induced by simulataneous downregulation of cinnamoyl CoA reductase (CCR) and cinnamyl alcohol dehydrogenase (CAD) in tobacco plants[J].PlantJournal, 2001, 28(3):257-270.
[22]YAMAGUCHI-SHINOZAKI K, SHINOZAKI K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses[J].AnnualReviewofPlantBiology, 2006, 57(6): 781-803.
[23]HU Y, LI W C, XU Y Q,etal. Differential expression of candidate genes for lignin biosynthesis under drought stress in maize leaves[J].JournalofAppliedGenetics, 2009, 50(3): 213-223.
[24]ZELLER G, HENZ S R, WIDMER C K,etal. Stress-induced changes in theArabidopsisthalianatranscriptome analyzed using whole-genome tiling arrays[J].PlantJournal, 2009, 58(6): 1 068-1 082.
[25]WALLEY J W, DEHESH K. Molecular mechanisms regulating rapid stress signaling networks inArabidopsis[J].JournalofIntegrativePlantBiology, 2010, 52(4): 354-359.
[26]RICHET N, TOZO K, ATIF D,etal. The response to daylight or continuous ozone of phenylpropanoid and lignin biosynthesis pathways in poplar differs between leaves and wood[J].Planta, 2012, 236(2): 727-737.
[27]BETZ G, KNAPPE C, LAPIERRE C,etal. Ozone affects shikimate pathway transcripts and monomeric lignin composition in European beech (FagussylvaticaL.)[J].EuropeanJournalofForestResearch, 2009, 128(2):109-116.
[28]TRONCHET M, BALAGUÉ C, KROJ T,etal. Cinnamyl alcohol dehydrogenases-C and D, key enzymes in lignin biosynthesis, play an essential role in disease resistance inArabidopsis[J].MolecularPlantPathology, 2010, 11(1):83-92.
[29]ZHAO Q, DIXON R A. Transcriptional networks for lignin biosynthesis: more complex than we thought[J].TrendsinPlantScience, 2011, 16(4): 227-233.
[30]BI C, CHEN F, JACKSON L,etal. Expression of lignin biosynthetic genes in wheat during development and upon infection by fungal pathogens[J].PlantMolecularBiologyReporter, 2011, 29(1): 149-161.
[31]KAUR H, SHAKER K, HEINZEL N,etal. Environmental stresses of field growth allow cinnamyl alcohol dehydrogenase-deficientNicotianaattenuataplants to compensate for their structural deficiencies[J].PlantPhysiology, 2012, 159(4): 1 545-1 570.
(编辑:宋亚珍)
Cloning and Expression Profile Analysis of the Gene Encoding Cinnamyl Alcohol Dehydrogenase under Abiotic Stress in Carrot
LIU Yanjun, MA Jing, WANG Guanglong, HUANG Wei, WANG Feng, XIONG Aisheng*
(State Key Laboratory of Crop Genetics and Germplasm Enhancement, Ministry of Agriculture Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in East China, College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China)
Here, the gene encoding DcCAD were cloned from carrot cultivar ‘Kurodagosun’ with RT-PCR method. The length ofDcCADwas 1 074 bp, and encoding 357 amino acids. The DcCAD may belong to hydrophilic protein. Phylogenetic analysis showed that DcCAD closed to CAD from tomato(XP_010314515.1). Quantitative real-time PCR analysis showed that the expression profiles of theDcCADgene are significantly different in different tissues (root, leaf blade, and petiole) of carrot. Relative expression level of theDcCADgene is leaf blade>root>petiole. TheDcCADgene is responsive to various abiotic stresses, such as heat (38 ℃), cold (4 ℃), drought (20% PEG) and salt (0.2 mol·L-1NaCl). The expression level was significantly increased after high temperature or low temperature treatments, respectively. The expression levels peaked at 1 h heat treatment and 2 h cold treatment, respectively. These results suggested thatDcCADgene participate in the response process of carrot under the abiotic stress.
cinnamyl alcohol dehydrogenase; homologous analysis; abiotic stress; expression analysis;Daucuscarota
1000-4025(2016)07-1294-08
10.7606/j.issn.1000-4025.2016.07.1294
2016-04-20;修改稿收到日期:2016-06-13
江苏省自然科学基金杰出青年基金(BK20130027);江苏高校优势学科建设项目(PAPD)
刘堰珺(1993-),女,硕士研究生,主要从事蔬菜分子生物学研究。E-mail: 2015104076@njau.edu.cn
熊爱生,教授、博士生导师,主要从事蔬菜遗传育种与分子生物学研究。E-mail: xiongaisheng@njau.edu.cn
Q785; Q786; S631.2
A
猜你喜欢
家庭科学·新健康(2021年5期)2021-06-21
中国调味品(2020年7期)2020-07-24
上海包装(2019年8期)2019-11-11
中国调味品(2019年2期)2019-03-18
心肺血管病杂志(2018年11期)2018-12-18
天津造纸(2016年1期)2017-01-15
西部林业科学(2016年3期)2016-10-25
中国医药生物技术(2015年4期)2015-12-26
中国造纸学报(2015年1期)2015-12-16
现代检验医学杂志(2015年5期)2015-02-06
