
不同干扰生境下河口莲座蕨遗传多样性分析
2016-08-30 07:31:07朱晓媛和兆荣王崇云张滋第
广西植物 2016年7期
朱晓媛,杨 斌,和兆荣,王崇云*,张滋第,刘 伟
( 1. 云南大学 生态学与地植物学研究所,昆明 650091; 2. 云南大学 生命科学学院,昆明 650091; 3. 文山学院,云南 文山 663000 )
不同干扰生境下河口莲座蕨遗传多样性分析
朱晓媛1,杨斌1,和兆荣2,王崇云1*,张滋第2,刘伟3
( 1. 云南大学 生态学与地植物学研究所,昆明 650091; 2. 云南大学 生命科学学院,昆明 650091; 3. 文山学院,云南 文山 663000 )
该研究利用 ISSR 分子标记,对不同生境下的6个河口莲座蕨种群进行遗传多样性分析。结果表明:从44条ISSR引物中筛选出8条能够扩增出清晰、稳定条带的引物,对6个河口莲座蕨种群进行基因组 DNA 扩增,共扩增出144条带,其中多态性条带119个,多态性条带比率为93.7%,Nei’s遗传多样性指数(H)为0.296,Shannon多样性指数(I)为0.457,遗传分化指数(Gst)为0.152 0,种群遗传一致度和遗传距离分别为0.913 8~0.954 8和0.058 4~0.090 1;UPGMA聚类分析表明,种群间遗传距离与空间距离和生境类型有关。河口莲座蕨在不同干扰程度的生境中,种群具有高水平的遗传多样性,一定强度或频率的干扰生境中,种群遗传多样性较高,与海拔、坡度和坡向无相关性。
河口莲座蕨, 干扰, ISSR, 遗传多样性
近年来,生物遗传多样性(Genetic diversity)研究越来越受到重视,其中大多以分子标记(RAPD标记、ISSR标记、SSR标记等)展开研究,为生物多样性保护、中度干扰、系统发育关系、交配系统等提供理论依据(潘丽芹等,2005;文陇英等,2006;周媛等,2009)。遗传多样性是生物多样性的重要组成部分,是生态系统和物种多样性的基础。物种以种群为具体的存在形式,任何一个物种都具有独特的基因库和遗传结构,同时物种多样性也显示了基因多样性(施立明,1990)。对一个物种而言,其遗传多样性既是维持繁殖活力和长期适应环境变化的基础,又是生物多样性保护的重要方面(陈灵芝等,2001)。中度干扰假说(Intermediate Disturbance Hypothesis)指出群落中物种丰富度在中等干扰水平时最大(Connell,1978;文陇英等,2006)。环境干扰导致选择压力差异,或引起种群波动,对种群遗传多样性分布与格局也会产生重要影响,但对此认识还比较欠缺(Banks et al, 2013)。中国水青冈在人为活动频繁的环境下,几乎没有实生苗,种群即使能生存主要也是靠根蘖萌生更新,这将不利于遗传多样性的保存(李俊清,1996)。濒危物种夏腊梅(Sinocalycanthuschinensis)在人为干扰的适宜生境中具有较高的遗传多样性(张文标等,2005);观光木(Tsoongiodendeonodorum)的遗传多样性也较高,但对生境要求严格,生境干扰是观光木濒危的主要原因之一(黄九香等,2002)。广布种冷蒿(Aremisafrigida)随干扰强度的增加,遗传多样性降低(王静等,2004);荒漠地带性广布种红砂(Reaumuriasoongorica)在不同的干扰生境下,具有不同水平的遗传多样性,无干扰的自然保护区种群遗传多样性水平较高(张颖娟等,2008)。不同干扰环境下,濒危种或广布种检测出不同的遗传多样性分布式样。对于一些具有较高遗传多样性的濒危植物来说,其濒危的主要因素之一是生境的破坏,随生境的破坏程度加强,遗传多样性逐渐降低,但有些植物在一定频率或强度干扰的适宜生境中可维持较高的遗传多样性。中度干扰假说是否适用于解释种群遗传多样性的分布与格局呢?
河口莲座蕨(Angiopterishokouensis)为莲座蕨科莲座蕨属植物,属广布种。河口莲座蕨产于云南罗平、西畴、麻栗坡、马关、屏边、河口、金平、绿春、江城等地(钱崇澍等,1959;朱维明等,2006;王崇云等,2012),生于常绿阔叶林及混交林下溪边,常为植物群落中草本层的优势种,其分布生境多样化,从原生性山地雨林到香蕉园地橡胶林,林缘农田沟边都有该植物生长,可见河口莲座蕨具有较广的分布范围并适应异质性的生境。野外观察到河口莲座蕨叶柄瘤突明显程度、大小和排列状况等存在较大形态变异,此外,云南植物志卷(二十卷)对该种收录了13个异名(朱维明等,2006),一度分类处理的混乱侧面反映了其丰富的形态变异。本文以河口莲座蕨为研究对象,利用ISSR分子标记技术对不同干扰生境下的河口莲座蕨种群开展遗传多样性研究,分析其遗传多样性格局和种群遗传分化程度,探讨不同干扰强度的河口莲座蕨种群遗传多样性特征,为莲座蕨科中的广布种、狭域种、濒危种的遗传多样性和种群分化,及其与生境干扰的关系研究积累资料,为这一类大型肉质蕨类植物的保护提供理论指导。
1 材料与方法
1.1 材料
以河口莲座蕨主要分布区—红河州河口县为中心,对滇东南的该物种分布区(图1)进行了生境实地调查,根据人类干扰强度划分群落生境类型并记录海拔、坡度、坡向,同时采集6个种群(表1),每个种群最少采集15株个体的新鲜叶片,选取植株上较好(鲜嫩、干净、无生虫、发黄等)的叶片,去除中间的主脉及叶边有破损的地方,后放入装有干燥硅胶的密封袋保存,同时保存一定数量的凭证标本。
1.2 方法
1.2.1 总DNA提取及检测采用改良的CTAB法(Murray et al,1980;邹喻萍等,2001)提取DNA,用0.8%琼脂糖凝胶电泳后于凝胶成像系统检验DNA质量。DNA样品置于-20 ℃冰箱中保存、备用。
1.2.2 ISSR引物筛选及反应体系选取8个DNA模板,根据加拿大哥伦比亚大学公布的ISSR引物序列设计44个ISSR引物进行筛选。经过反复实验,选择扩增条带清晰、反应稳定且多态性较好的8个ISSR引物(表2)对所有DNA样品进行扩增。
PCR反应体系的总体积为25 μL,模板DNA 32 ng·μL-1,Taq聚合酶1U,dNTP 0.2 mmol·L-1,引物1.3 mmol·L-1,Mg2+1.5 mmol·L-1。扩增程序: 94 ℃,5 min,38个循环;94 ℃,45s;48~58 ℃,1 min;72 ℃,1 min;72 ℃,5 min,4 ℃保存。PCR产物用2% (含EB约0.5 μg·mL-1) 的琼脂糖凝胶电泳检测,电压为3 V·cm-1,电泳2 h。电泳结果用Gene System凝胶成像系统照相,记录结果。

表 1 分子实验材料采集地群落样地综合简表Table 1 General description of communit of Angiopteris hokouensis
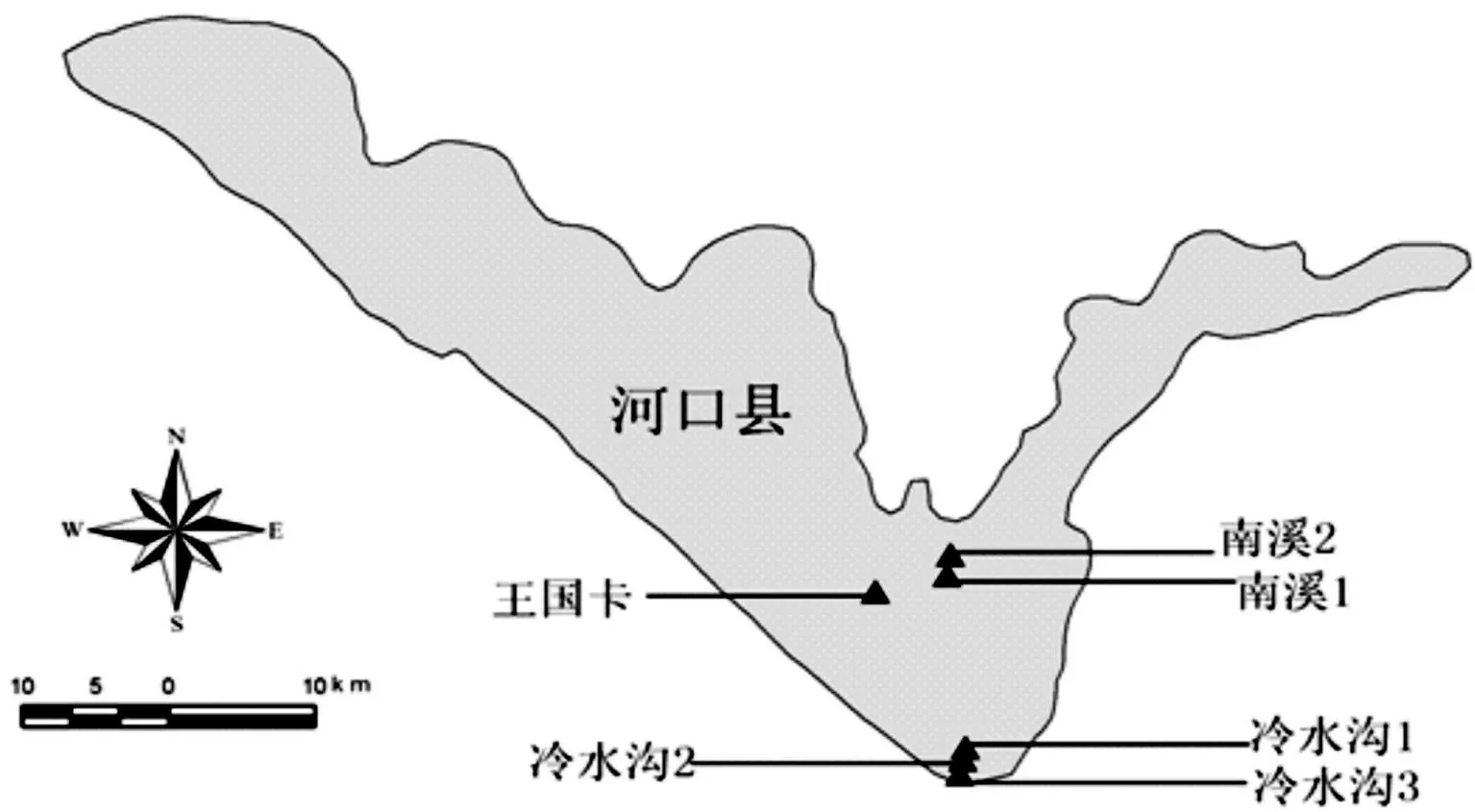
图 1 分子实验材料采集地分布图Fig. 1 Distribution of Angiopteris hokouensis
1.2.3 统计与分析每个样品的扩增条带通过UVIPhoto软件判读,POPGENE32软件进行遗传多样性参数分析,遗传多样性参数分别为多态性位点比(PPB)、有效等位基因数、Shannon多样性指数(I)、Nei’s遗传多样性指数(H)、基因分化系数(Gst)、Nei’s遗传一致度、遗传距离;采用SPSS17.0软件进行平均数差异的显著性检验和相关性分析。
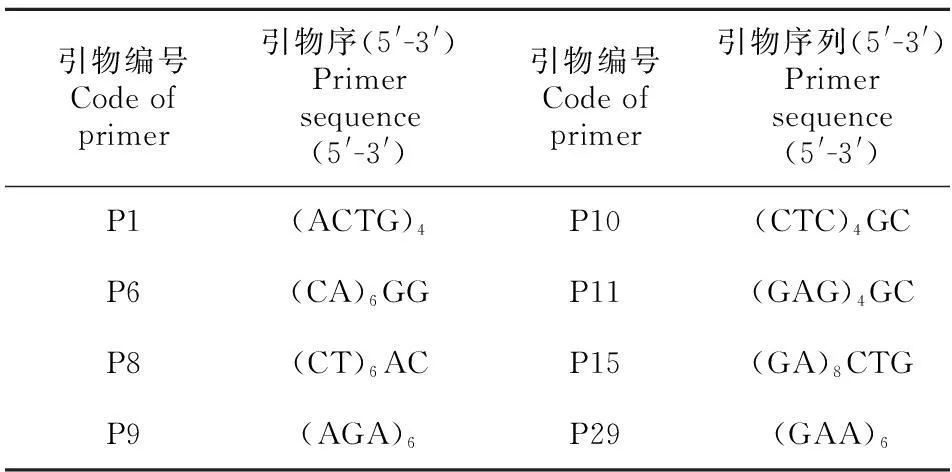
表 2 ISSR引物序列Table 2 Sequence of 8 primers
2 结果与分析
2.1 遗传多样性及遗传分化
8个ISSR引物在6个种群中共扩增出127条,平均每个引物扩增出14.88条带,多态性条带数为119个,PPB为93.7%。
表3显示,Shannon指数的遗传多样性各种群大小顺序为冷水沟2>冷水沟3>南溪1>冷水沟1>王国卡>南溪2,与多态位点比率稍有差异。Nei’s基因多样性指数计算的结果与Shannon指数计算的结果一致,各种群在0.230~0.270 0之间变化。
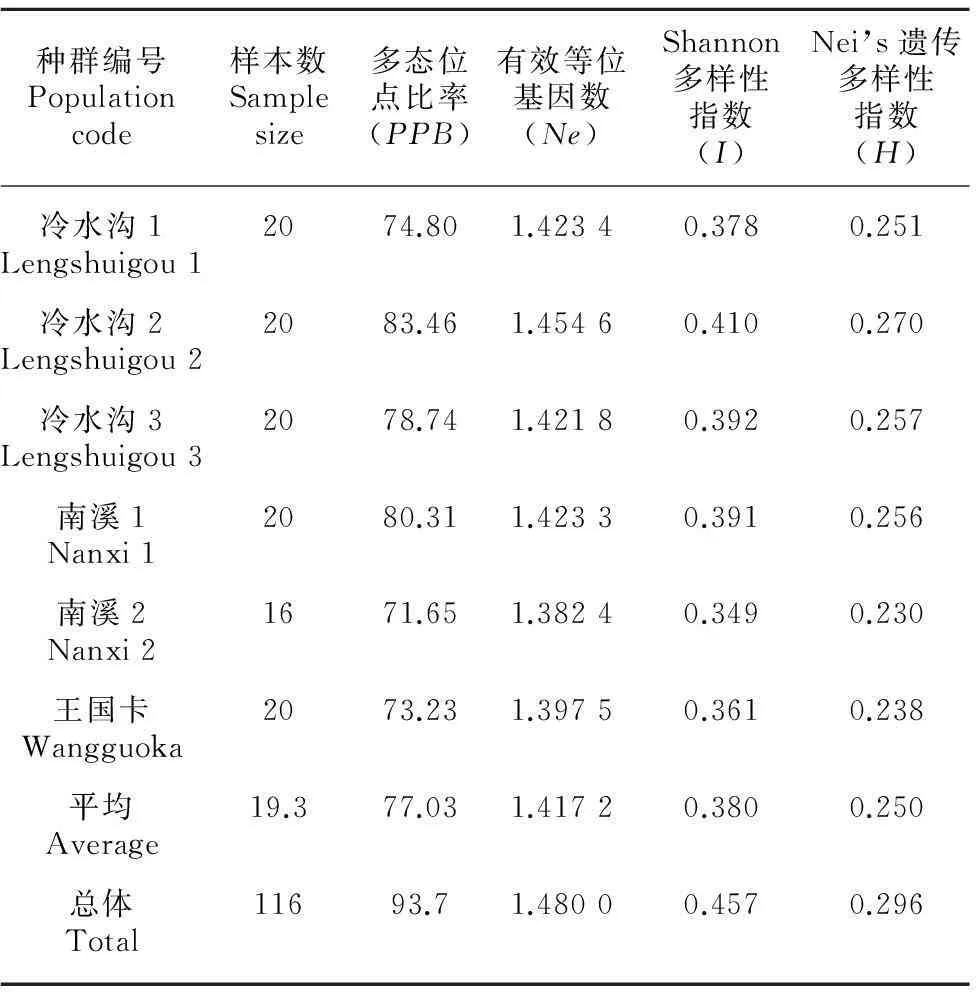
表 3 6个河口莲座蕨种群多态性及遗传差异统计Table 3 Polymorphic loci and genetic diversity in 6 Angiopteris hokouensis populations
根据Nei’s基因多样性指数计算的种群遗传分化结果,6个种群总的遗传多样性Ht=0.295 2,种群内遗传多样性Hs=0.250 3,种群间遗传多样性Dst=0.044 9,各种群间存在一定遗传变异,遗传分化系数Gst=0.152 0,即15.2%的变异存在于种群间,84.8%的遗传变异存在于种群内。基于种群间遗传分化系数计算的基因流Nm=2.789 4>1,说明不同生境的种群间存在一定基因流,防止因遗传漂变导致的种群间的遗传分化。
2.2 种群遗传一致度和聚类分析
Nei’s遗传一致度和遗传距离反映了种群间的亲缘关系远近。河口莲座蕨6个种群的遗传一致度在0.913 8~0.954 8之间,遗传距离在0.046 3~0.090 1之间(表4)。
根据遗传距离,运行POPGENE32软件,采用UPGMA法构建的种群遗传关系聚类图(图2)。聚类图中可看出,6个种群可分为两大类群:第一类群仅南溪2种群;第二类群包括其余的5个种群,即冷水沟1、冷水沟2、冷水沟3、南溪1和王国卡种群。
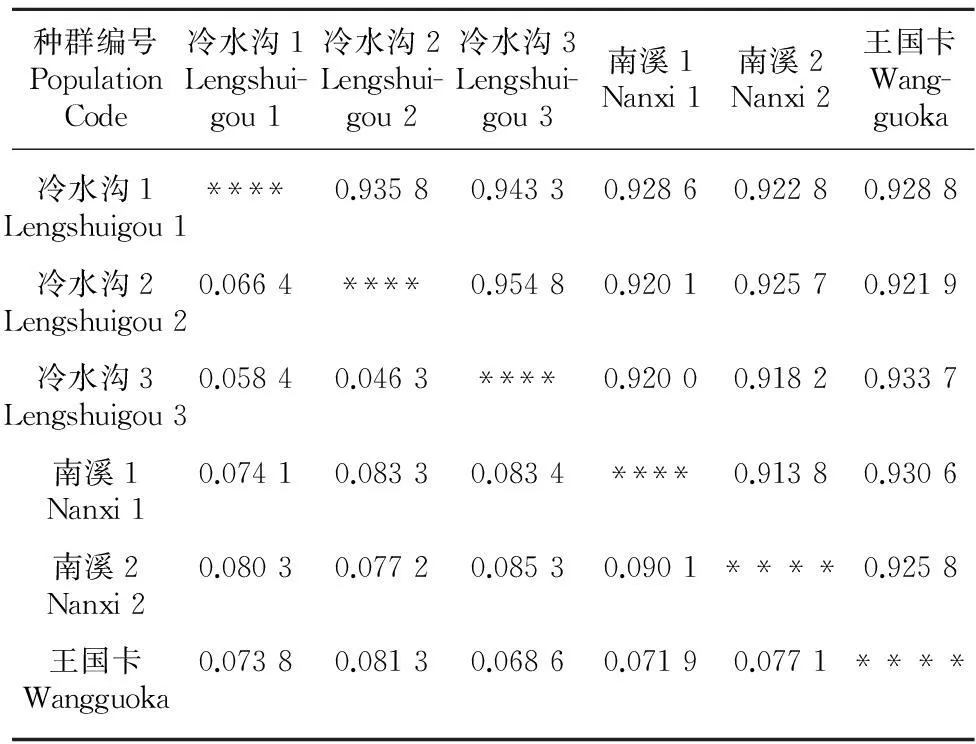
表 4 河口莲座蕨种群的遗传一致度和遗传距离Table 5 Genetic identity and genetic distance among the populations of Angiopteris hokouensis
注:对角线上为遗传一致度;对角线下为遗传距离。
Note: On the diagonal, genetic identity; below diagonal, genetic distance.
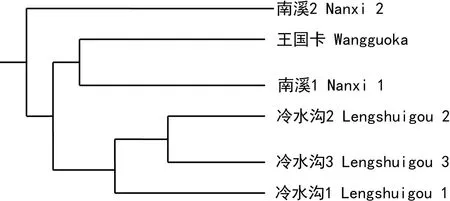
图 2 ISSR-PCR种群遗传距离聚类图Fig. 2 Dendrogram of ISSR cluster analysis of 6 populations
在第二类群中,冷水沟2和冷水沟3的遗传距离最小,为0.046 3,首先聚为一支,其次冷水沟1与它们的距离也较小,分别为0.066 4和0.058 4,再聚为一支;再次是南溪1和王国卡种群聚为一支,二者的遗传距离为0.071 9。
其中冷水沟1、冷水沟2、冷水沟3空间分布上较为接近,其次是南溪1和南溪2空间分布上较为接近,但冷水沟2、冷水沟3与冷水沟1生境类型不同,南溪1和南溪2生境类型也不同,有着较大的遗传距离。
2.3 海拔、坡度和坡向与遗传多样性
6个河口莲座蕨的遗传多样性与海拔、坡度和坡向进行相关性分析,结果表明海拔、坡度和坡向与多态位点比率(r=0.606,P=0.202)、Shannon多样性指数(r=0.740,P=0.092 )和Nei’s遗传多样性指数(r=0.763,P=0.077)均无显著相关关系。
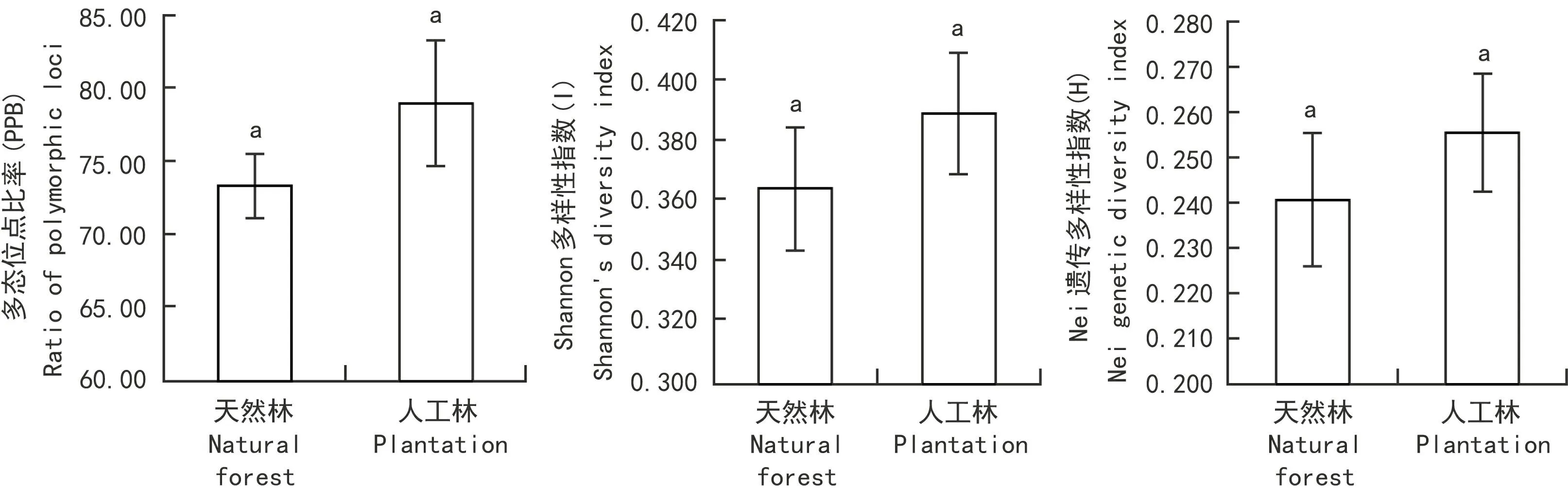
图 3 生境类型与遗传多样性的相关性Fig. 3 Relationship between habitat types and genetic diversity
2.4 生境与遗传多样性
结合采集地的不同干扰程度,将6个种群分为两类,即人工林型种群的冷水沟2、冷水沟3、南溪1、王国卡为一类,代表天然林种群的冷水沟1(次生林)和南溪2(原生林)为一类,两种生境类型的种群,多态位点比率、Shannon多样性指数、Nei’s遗传多样性指数(P>0.05)显著性差异(图3)。
3 讨论
Connell(1978)通过大量生态学研究提出了中度干扰假说,即中等程度的干扰水平能维持高度生物多样性。中度干扰假说是生态学中引用率最高的假说之一,但也存在广泛的质疑,Fox(2013)指出仅有少于20%的研究显示中度干扰水平下生物多样性水平达到最高峰。干扰,尤其是中度干扰能增加生境及生态过程的多样性(叶林奇,2000),导致歧化选择过程,并发生遗传分化,形成不同的遗传多样性水平。广布种出现濒危状况的主要原因之一是生境遭到人类的破坏,导致其生境类型改变,这些广布种往往具有较高水平的遗传多样性,因此,干扰对其种群遗传结构将会有重要的影响。
本研究野外观察发现,作为林下植物的河口莲座蕨在不同群落生境中长势有明显差异,人工林和次生林中长势较好。本研究6个调查种群均表现出较高程度的遗传多样性,在不同干扰程度的群落生境中具有不同水平的遗传多样性,一定强度干扰的种群与其它种群相比,具较高的遗传多样性,但无显著性差异。同时,遗传多样性与海拔、坡度、坡向无显著相关关系。产于英属Pitcairn岛的Angiopterischauliodonta及产于台湾的伊藤氏原始莲座蕨(Archangiopterisitoi)为濒危及特有种,这两物种的RAPD分子标记表明,在人类干扰下其遗传多样性较低(Naomi et al,2004;Xu et al,2000)。与上述两物种相比,河口莲座蕨遗传多样性较高,具更好的环境适应能力,且不同群落生境类型下的河口莲座蕨种群都有较高的遗传多样性,存在一定强度人为干扰的生境中,种群具有相对高的遗传多样性水平。
通常良好的环境资源、较高的空间异质性和一定强度的歧化选择,都有利于植物维持较高的遗传多样性(红雨等,2006;张志红等,2005;黎中宝等,2000)。因此,对于适应环境能力较强、分布较广的河口莲座蕨来说,自然环境条件的不同(海拔、坡向)并不是造成其遗传多样性较高的因素,但一定强度或频率的干扰可能对维持其遗传多样性水平有更重要的意义。较高的遗传多样性可能是对干扰的一种适应,其它广布种也存在类似的情况(张颖娟等,2008)。
4 结论
(1)用8条引物进行ISSR分析,扩增共产生127条带,其中119为多态性条带,PPB为93.7%,说明6个河口莲座蕨种群具有高水平的遗传多样性。(2)河口莲座蕨采自河口县不同地方,地形因子存在一定的差异,相关性分析显示海拔、坡度和坡向的差异对遗传多样性无显著影响。(3)不同干扰生境中,河口莲座蕨的遗传多样性存在一定差异。不同生境类型中,遗传多样性差异不显著,但一定程度或频率的干扰种群和其它种群相比,遗传多样性水平更高。
BANKS SC,GEOFFREY JC,ANNABEL LS,et al,2013. How does ecological disturbance influence genetic diversity? [J]. Trends Ecol Evol ,28(11) :670-679.
CHEN LZ, MA KP. 2001. Biodiversity science: principle and practice [M].Shanghai: Shanghai Scientific & Technical Publishers. [陈灵芝, 马克平. 2001. 生物多样性科学:原理与实践 [M].上海: 上海科学技术出版社.]
CONNELL JF, 1978. Diversity in tropical rain forests and coral reefs [J]. Sci,199(4 335):1 302-1 310.
FOX JW, 2013. The intermediate disturbance hypothesis should be abandoned [J]. Trends Ecol Evol,28(2) :86-91.
HONG Y,WANG LH,LIANG XH, 2006. Analysis on the genetic diversity of sabina vulgaris at different habitats and the influence of the environmental factors [J]. J Arid Land Res Environ,20(3):184-187. [红雨,王林和,梁小和, 2006. 不同生境臭柏种群的遗传多样性分析及其与环境因子的相关性 [J]. 干旱区资源与环境,20(3):184-187. ]
HUANG JX,ZHUANG XY, 2002. Genetic diversity of structure of the population ofTsoongiodendeonodorum[J]. Acta Phytoecol Sin,26(4):413-419. [黄九香,庄雪影, 2002. 观光木种群遗传多样性研究 [J]. 植物生态学报,26(4):413-419. ]
LI LQ, 1996. Studies on intra-and inter-species gene diversity of chinese beeches [J]. Biol Divers Sci,4(2):63-68. [李俊清, 1996. 中国水青冈种内种间遗传多样的初步研究 [J]. 生物多样性,4(2):63-68. ]
LI ZB,LI P,DENG CY, 2000. Genetic diversity and differentiation ofAegicerascorniculatumpopulations in three different habitat [J]. J Oceanogr Taiwan St,19(3):379-385. [黎中宝,林鹏,邓传远, 2000. 三种不同生境的桐花树种群的遗传多样性和遗传分化 [J]. 台湾海峡,19(3):379-385. ]
MURRAY MG,THOMPSON WF, 1980. Rapid isolation of high molecular weight plant DNA [J]. Nucl Acid Res,8(19):4 321-4 325.
NAOMI K,STEVE W,NOELEEN S, 2004. Conservation genetics and ecology ofAngiopterischauliodontaCopel. (Marattiaceae),a critically endangered fern from Pitcairn Island,South Central Pacific Ocean [J]. Biol Conserv,(117):309-319 .PAN LQ,JI H,CHEN LQ, 2005.Genetic diversity of the natural populations ofAdiantumreniformevar.sinense[J]. Biol Divers Sci,13(2):122-129. [潘丽芹,季华,陈龙清, 2005.荷叶铁线蕨自然居群的遗传多样性研究 [J]. 生物多样性,13(2):122-129.]
QIAN CS, CHEN HY. 1959. Flora of China (Vol.2) [M].Beijing: Science Press. [钱崇澍, 陈焕镛. 1959. 中国植物志(第2卷) [M].北京:科学出版社.]
SHI LM, 1990. Genetic diversity and its conservation [J]. Bioin formatics,2(4):158-164. [施立明, 1990. 遗传多样性及其保存 [J]. 生物科学信息,2(4):158-164.]
WANG CY,YANG B,HE ZR,et al, 2012.Analysis on the geographical distribution characteristics of angiopteridaceae and its causes in Yungui plateau and Hengduanmountains,China [J]. Plant Diverers Res,34(4):317-325. [王崇云,杨斌,和兆荣,等, 2012.云贵高原及横断山区莲座蕨科植物地理分布特征及其格局成因初探 [J]. 植物分类与资源学报,34(4):317-325.]
WANG J,YANG C,YIN J,et al,2004. Changes of the genetic diversity ofAremisafrigidapopulation under the disturbance of grazing [J]. Acta Ecol Sin,24(11):2 466-2 471. [王静,杨持,尹俊,等,2004. 冷蒿种群在放牧干扰下遗传多样性的变化 [J]. 生态学报,24(11):2 466-2 471.]
WEN LY,LI ZF, 2006.The effects of disturbance on maintaining mechanism of species diversity [J]. J NW Norm Univ,42(4):87-91. [文陇英,李仲芳, 2006.干扰对物种多样性维持机制的影响 [J]. 西北师范大学学报,42(4):87-91.]
XU ZW,WANG JB, 2000. Low RAPD polymorphism inArchangiopterisitoi,a rare and endemic fern in Taiwan [J]. Bot Bull Acd Sin,41:15-18.
YE LQ, 2000. Relation between discurbance and biodiversity [J]. J Guizhou Univ: Nat Sci Ed,17(2):129-134. [叶林奇, 2000. 干扰与生物多样性 [J]. 贵州大学学报·自然科学,17(2):129-134.]
ZHANG WB,JIN ZX,LI JM,2005. Genetic diversity ofSinocalycanthuschinensisin four different habitats revealed by RAPD [J]. Bull Bot Res,32(3):313-318. [张文标,金则新,李钧敏,2005. 不同生境夏腊梅群体遗传多样性的RAPD分析 [J]. 植物研究,32(3):313-318.]ZHANG YJ,GAO RX,LI QF,2008. Analysis on the population genetic diversity of desert shrubReaumuriasoongoricain disturbed habitats [J]. J Arid Land Res Environ,22(3):147-151. [张颖娟,高瑞霞,李青丰,2008. 不同干扰生境中荒漠小灌木红砂种群遗传多样性研究 [J]. 干旱区资源与环境,22(3):147-151.]
ZHANG ZH,TANG T,ZHONG RZ,et al, 2005. Effects of divergent habitat on genetic structure of population ofExcoecariaagallocha,a mangrove associate [J]. J Genet Genomics,32(12):1 286-1 292. [张志红,唐恬,周仁超,等, 2005. 异质性生境对半红树植物海漆(Excoecariaagallocha)居群遗传结构的影响 [J]. 遗传学报,32(12):1 286-1 292.]
ZHOU Y,GAO L,WANG ZW,et al,2009. Application of molecular marker techniques in genetic diversity of pteridophytes [J]. J Wuhan Bot Res,27(6):667-673. [周媛,高磊,汪志伟,等,2009. 分子标记技术在蕨类植物遗传多样性研究中的应用 [J]. 武汉植物学研究,27(6):667-673.]
ZHOU YP, GE S, WANG XD. 2001. Molecular markers in systematic and evolutionary botany [M]. Beijing: Science Press [皱喻萍, 葛颂, 王晓东. 2001. 系统与进化植物学中的分子标记 [M]. 北京:科学出版社. ]
ZHU WM, ZHANG GF, LU SG, et al. 2006. Flora of Yunnan (Vol. 20) [M]. Beijing: Science Press. [朱维明, 张光飞, 陆树刚, et al. 2006. 云南植物志(第20卷) [M]. 北京:科学出版社.]
Analysis on the genetic diversity of Angiopterishokouensisin habitats under different disturbances
ZHU Xiao-Yuan1, YANG Bin1, HE Zhao-Rong2,WANG Chong-Yun1*, ZHANG Zi-Di2, LIU Wei3
( 1. Institute of Ecology and Geobotany, Yunnan University, Kunming 650091, China; 2. College of Life Sciences,YunnanUniversity, Kunming 650091, China; 3.WenshanTeachers’College, Wenshan 663000, China )
Genetic diversity of 6Angiopterishokouensispopulations collected from different habitats were analyzed by means of intersimple sequence repeat (ISSR) markers, and 8 proper primers with rich polymorphism and stable bands were selected from a total of 44 ISSR’s primers. The genome DNAs of the 6 populations were amplified to a total of 144 putative bands by the 8 primers, in which 96 were polymorphism bands. Polymorphism rate was 93.7%; Nei’s gene diversity index was 0.296, Shannon information index was 0.457, andGst was 0.152 0. The genetic distance coefficient and the genetic similarity were 0.058 4-0.090 1 and 0.913 8-0.954 8. Cluster analysis by UPGMA indicated that the genetic distance between the populations related to spatial distance, as well as to habitats. The results showed thatA.hokouensispopulations had high levels of genetic diversity at disturbed habitats. Even there was no significant differentiation of genetic diversity among populations,the populations in moderate disturbed habitats had higher genetic diversity. The genetic diversity was not significantly related toaltitude,slope and aspect.
Angiopterishokouensis, disturbance,ISSR, genetic diversity
10.11931/guihaia.gxzw201312056
2014-12-30
2015-03-27
国家自然科学基金(30260011,31260078) [Supported by the National Natural Science Foundation of China (30260011,31260078)]。
朱晓媛(1987- ),女,云南玉溪人,硕士研究生,主要从事外来入侵生物分子标记技术研究,(E-mail)zhuxiaoyuan1987@126.com。
王崇云,博士,副教授,研究方向为植物生态学、进化生态学、生物多样性保护等,(E-mail)cywang@ynu.edu.cn。
Q943
A
1000-3142(2014)07-0818-06
朱晓媛,杨斌,和兆荣,等. 不同干扰生境下河口莲座蕨遗传多样性分析 [J]. 广西植物,2016,36(7):818-823
ZHU XY,YANG B,HE ZR,et al. Analysis on the genetic diversity ofAngiopterishokouensisin habitats under different disturbances [J]. Guihaia,2016,36(7):818-823
猜你喜欢
新少年(2023年3期)2023-03-20 10:16:30
中国书法(2023年12期)2023-02-02 15:51:36
中国民族民间医药(2021年22期)2021-12-17 05:16:02
新少年(2021年3期)2021-03-28 02:30:27
东坡赤壁诗词(2018年6期)2018-12-22 10:57:10
西部林业科学(2016年2期)2016-05-07 06:15:47
少儿科学周刊·少年版(2015年11期)2015-12-17 20:59:10
少儿科学周刊·少年版(2015年11期)2015-12-17 20:57:47
少儿科学周刊·少年版(2015年11期)2015-12-17 20:55:51
少儿科学周刊·少年版(2015年11期)2015-12-17 20:48:15
