
丛枝菌根真菌(AMF)对玉米/蚕豆和玉米/稗草互作体系植物生长的反馈调节
2016-08-30 02:51谢先进张俊伶
植物营养与肥料学报 2016年3期
谢先进, 张俊伶
(1中国农业大学资源与环境学院, 北京 100193; 2河南省土壤肥料站, 河南郑州 450002)
丛枝菌根真菌(AMF)对玉米/蚕豆和玉米/稗草互作体系植物生长的反馈调节
谢先进1,2, 张俊伶1*
(1中国农业大学资源与环境学院, 北京 100193; 2河南省土壤肥料站, 河南郑州 450002)
【目的】丛枝菌根真菌(arbuscular mycorrhizal fungi,简称AMF)对农田生态系统中作物的生长起着重要作用。不同宿主植物和作物种植方式对土壤中菌根真菌具有一定的选择性,从而影响菌根真菌对后茬植物生长的反馈效应。间作体系是农业生产中增产增效的一种重要的种植模式,明确AMF在间作体系中对植物生长的反馈作用,对理解间作体系地上-地下相互作用具有重要的意义。【方法】本试验采用三种常见的AMF(Funneliformismosseae、Rhizophagusintraradices和Claroideoglomusclaroideum)混合接种剂,在单作(玉米,蚕豆,稗草)和间作(玉米/蚕豆和玉米/稗草)条件下,通过两个阶段(AMF驯化及反馈)的反馈试验,模拟研究了三种植物在单作和间作中的反馈强度及AMF的调节作用。【结果】在驯化阶段,玉米/稗草间作体系中,与单作相比,间作玉米地上部生物量减少了64.0%,间作稗草地上部生物量增加了47.8%。玉米/蚕豆间作体系中,与单作相比,两者作物地上部生物量增量分别达21.7%和38.3%。反馈阶段中,单作时,与灭菌处理相比接种AMF后玉米、稗草和蚕豆的地上部生物量分别增加602.3%、 80.6%和21.1%; 间作时,与灭菌处理相比接种AMF后玉米地上部生物量平均增加613.1%,稗草增加80.7%, 蚕豆增加21.4%。单作玉米存在负到零反馈作用,与灭菌处理相比,接种AMF后玉米负反馈作用减弱至零反馈作用。从单作到间作,玉米的反馈作用由负变为零到正反馈作用。【结论】玉米和稗草之间是竞争关系,玉米处于竞争弱势稗草处于竞争优势。玉米对AMF的响应最为强烈,其次是稗草,最后是蚕豆。接种AMF或与其它植物间作后均减弱玉米的负反馈作用,表明丛枝菌根真菌可通过减弱单作玉米的负反馈实现间作增产。
丛枝菌根真菌; 植物-土壤反馈; 蚕豆; 玉米; 稗草
目前已有很多的理论来解释种间互惠作用,其中土壤微生物在间套作增产中的作用日益引起人们的关注[1-2]。丛枝菌根真菌(arbuscular mycorrhizal fungi,简称AMF)是最重要的有益土壤微生物之一,是维持自然生态系统中植物群落多样性和生产力的主要因素之一[3-7]。由于AMF与宿主植物之间存在一定的宿主偏好性,导致在种植不同植物的土壤中AMF群落组成和数量存在差异,并进一步影响该种或者共存植物的生长,从而影响植物种间的相互作用[8-10]。在间套作体系中,地上部植物多样性的增加为土壤微生物提供了区别于单作的生长环境[11],导致土壤微生物群落和功能的改变,而这种微生物群落的改变反过来又会影响该种植物或其它植物的生长和适应性[4],该过程被称为反馈作用。在自然生态系统中,植物-土壤反馈作用是植物群落多样性和生态功能的主要驱动力之一[12-14],然而关于反馈作用在间套作种植体系中的作用还缺乏系统的认识。现有研究发现,作物的种植模式(如间、套作)显著影响土壤中AMF群落组成和多样性[2,15],AMF则能够影响作物产量[16-17]。因此定量反馈作用的强度对深入理解AMF在间套作体系的作用具有重要的意义。
玉米和蚕豆是我国西北地区重要的间作模式之一。间作的双方存在互惠作用,表现在对土壤养分和地上光热资源的高效利用上,这一体系的增产效应是作物地上-地下相互作用的结果[18-19]。玉米/稗草则为竞争体系,且最近研究发现玉米和杂草对菌根响应不同[7]。因此,本试验选用这两种间作种植体系进行盆栽模拟试验来研究间作体系AMF对植物生长的调节反馈作用。
1 材料与方法
1.1试验材料
供试植物为玉米—郑单958(ZeamaysL. cv. Zhengdan No. 958)、蚕豆—89-147(ViciafabaL. cv. 89-147)、稗草(Echinochloacrus-galliL.P. Beauv.)。3种植物组合成两种植物互作体系,即玉米-稗草和玉米-蚕豆间作体系,和3种植物单作,共5个处理。
试验用土为农田土壤和河沙的混合物。农田土壤取自北京市昌平区昌平长期定位试验站,土壤有机质含量7.8 g/kg、Olsen-P 2.9 mg/kg、有效钾(NH4OAc浸提)84.6 mg/kg、pH 7.2。土壤过2 mm筛,用γ射线(剂量为25 kGy)灭菌后按土沙质量比3 ∶1混匀。
供试菌种选用三种AMF菌种,分别为Funneliformismosseae(BGC XZ01)、Rhizophagusintraradices(BGC USA07)和Claroideoglomusclaroideum(BGC HZB07D)。菌种由北京市农林科学院植物营养与资源研究所王幼姗副研究员提供。三种菌种按照相同的孢子数量混合均匀,作为驯化阶段的接种剂。
试验装置: 驯化阶段用小塑料盆25个,装基质约1.6 kg; 大塑料盆25个,装基质约10 kg。反馈阶段用塑料盆375个,装基质约1.0 kg。
1.2试验方法
该试验中植物的培养是在中国农业大学资源与环境学院日光温室内完成,培养温度在14℃到30℃之间。第一步AMF驯化阶段的时间为2013年3月25日至2013年6月14日; 第二步反馈阶段的时间从2013年7月25日到2013年9月12日。
驯化阶段: 先将所有种子用10%的 H2O2浸泡30 min灭菌,用去离子水冲洗干净后放入25℃培养箱中催芽。每盆称取1.6 kg无菌基质,以溶液的形式加入养分,加入量(mg/kg)为N 200、P 20、K 200、Mg 100、Fe 5.0、Mn 5.0、Cu 5.0、Zn 5.0、B 5.0。将AMF菌种充分混匀后加入到盆中种子下2 cm左右。每个处理重复5盆。出苗一周左右,单作处理玉米、稗草、蚕豆分别每盆保留2、12、2株,玉米/稗草间作处理玉米、稗草分别每盆留1、6株,玉米/蚕豆间作处理玉米、蚕豆分别每盆留1、1株。为了保证菌根能较好地侵染,在生长4周后将植物与其所培养的基质一起移栽至装有8 kg无菌基质的塑料盆中。每隔1 d称重浇水,保持含水量为最大持水量的75%左右,每隔两周随机调换试验盆的位置。生长约3个月时,收获植株地上部,在65℃烘至恒重,然后称重。将驯化的基质过2 mm筛,即得到5种相应的驯化土壤,将其风干,于4℃保存,以备反馈阶段使用。
反馈阶段: 灭菌,消毒及催芽同驯化阶段。为排除菌根真菌外其它微生物的影响,此阶段共有两种土壤微生物处理,1)-M—接种除AMF外其它微生物(100 g经γ射线灭菌驯化基质 + 100 g不灭菌驯化基质<20 μm的滤液); 2)+M—接种AMF(100 g经γ射线灭菌驯化基质 + 100 g不灭菌驯化基质中AMF孢子)。每盆称取900 g无菌基质作为背景基质(即接种量为10%[9]),养分的添加以及接种方式同上。每种土壤处理上分别种植单作玉米、单作稗草、单作蚕豆、间作玉米/稗草和玉米/蚕豆。共有5种驯化土壤,每种有2个微生物处理,5个植物处理,每处理重复5次,共250盆。出苗一周左右定苗,单作处理玉米、稗草、蚕豆每盆分别保留1、 2、 1株,玉米/稗草间作处理每盆1株玉米和2株稗草,玉米/蚕豆间作处理每盆1株玉米和1株蚕豆。浇水、管理和收获同上。
主要计算公式如下:
植物的间作效应(intercropping effect,%)=(BI - BM)/BM×100
其中, BI为间作时植物地上部生物量,BM为单作时植物地上部生物量。
植物-土壤反馈强度(feedback strength,%)=(Oi-F)/F×100[20]
其中,Oi表示反馈阶段中第i个植物种植在由本地驯化的土壤中的地上部生物量,F表示反馈阶段中植物种植在由外来植物驯化的土壤中的生物量的平均值。若结果为正值,为正反馈,反之为负反馈; 若结果与零没有差异,则为零反馈。
1.3统计分析
植物的间作效应和AMF的效应通过独立样本t检验(independentt-test)进行分析。植物的反馈作用通过单样本t检验(one samplet-test)进行分析。在反馈阶段中,在每种种植方式处理下,以土壤来源(soil origin)和土壤微生物(soil microbial communities)为因素进行Two-way ANOVA分析,并采用DUNCAN方法进行多重比较。
2 结果与分析
表1表明,除了与玉米间作的蚕豆以外,微生物处理对单作和间作玉米、稗草和单作蚕豆的生长菌有显著性影响。驯化土壤来源对与蚕豆间作的玉米生长存在显著性影响。并且两个因素的交互作用显著地影响了与蚕豆间作的玉米和单作稗草的生长。
2.1间作效应
在玉米/稗草间作体系中,玉米和稗草的间作效应分别为-64.0%和47.8%; 玉米/蚕豆间作体系中,玉米和蚕豆的间作效应分别为21.7%和38.3%(表2)。
2.2菌根效应
在-M处理中植物根系未被丛枝菌根真菌侵染,而+M处理植株根系侵染率(M%)在65%85%之间,不同土壤来源之间无显著性差异。从表3可以看出,与-M处理相比,接种AMF能显著提高玉米地上部生物量,单作时生物量增加602.3%,与稗草间作时增加637.8%,与蚕豆间作时增加588.5%。接种AMF比不接种AMF的单作和间作稗草地上部生物量分别显著提高约80.6%和80.7%。+M处理比-M处理单作蚕豆地上部生物量显著提高21.1%,与玉米间作时提高21.4%。
2.3反馈作用
图1显示,单作条件下,在-M处理下,除了当外来土壤为稗草单作时玉米为正反馈外,其它处理玉米植株的生长均表现出从零到负的反馈作用,而在+M处理下,玉米对所有的外来土壤反馈作用值接近于零。单作稗草对玉米/稗草间作表现为零反馈作用,而对单作玉米的反馈作用随着AMF从零到负反馈转变为正反馈作用。单作蚕豆的反馈作用较弱,以零反馈为主。间作玉米和间作稗草以零到正反馈作用为主,间作蚕豆以零反馈为主。
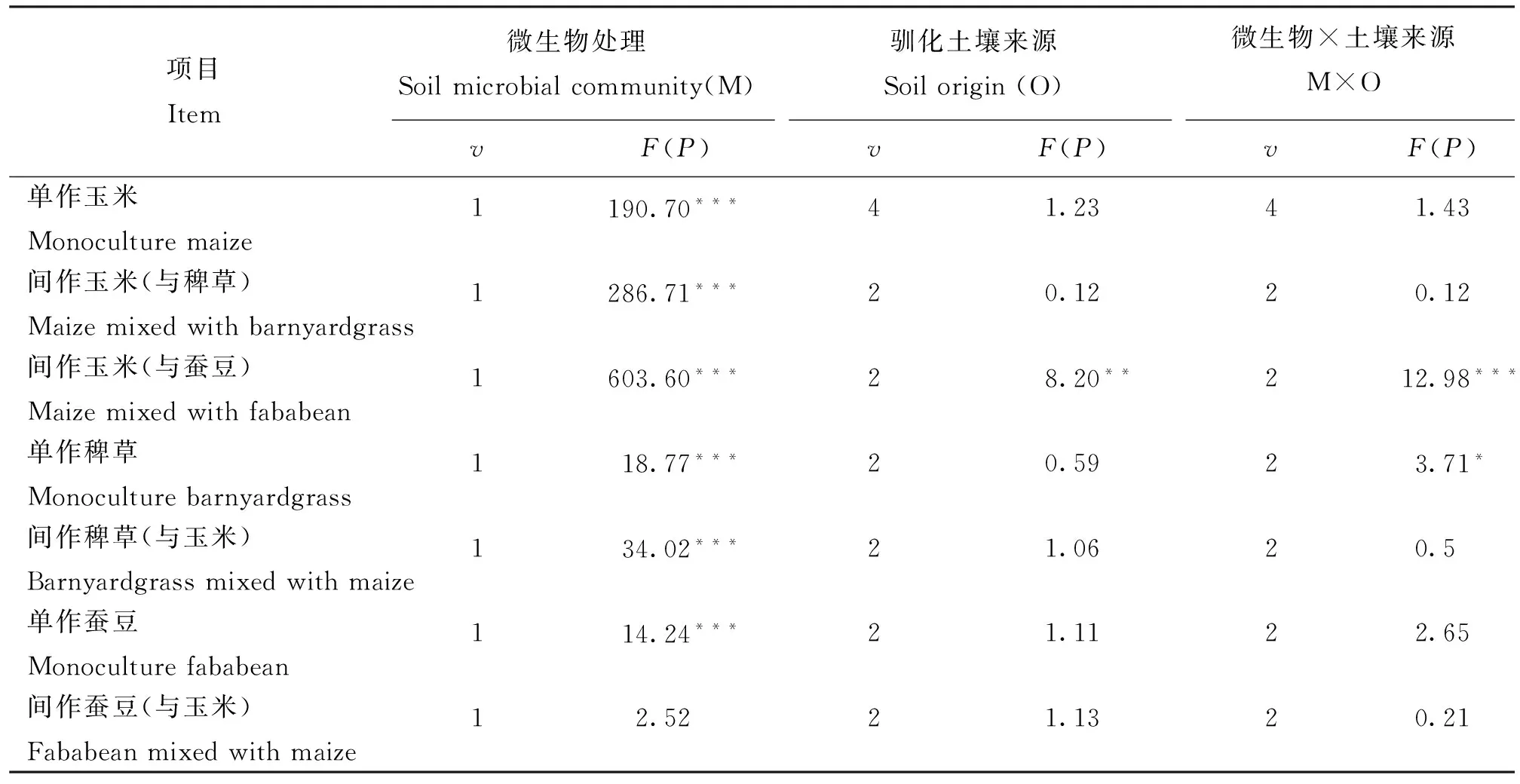
表1 土壤来源和微生物处理对玉米、稗草和小麦单株地上部生物量的影响的方差分析结果
注(Note): *—P≤ 0.05; **—P≤ 0.01; ***—P≤0.001.
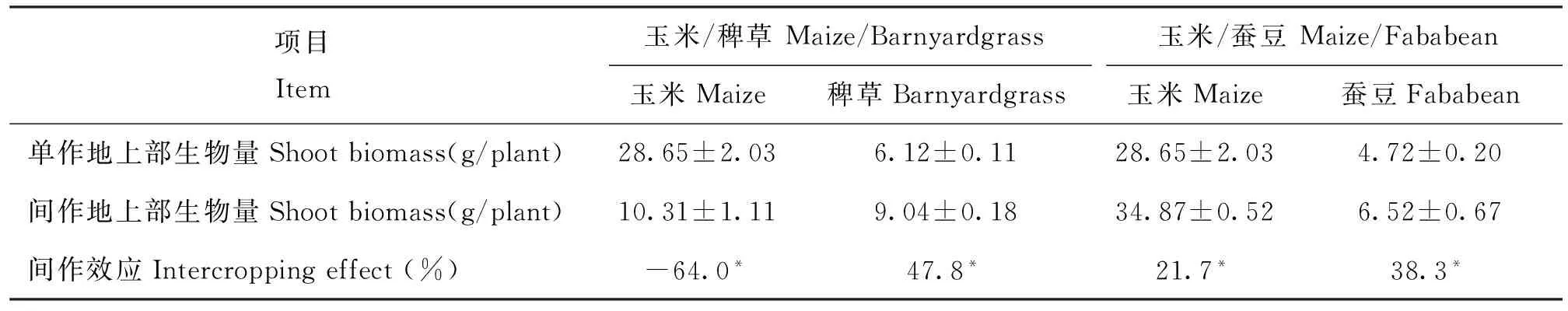
表2 反馈阶段两种间作体系中植物的间作效应
注(Note): *表示经过独立样本t检验后单作与间作之间单株生物量存在显著性差异(P≤ 0.05) Asterisk means significant difference in shoot biomass of monoculture and intercropping after independentt-test (P≤ 0.05).
3 讨论
3.1间作生长效应
在玉米/稗草间作体系中,稗草的生物量增加,而玉米生物量减少,表明该体系是一种竞争体系,这可能是由于玉米、稗草均为禾本科植物,相似的生态位和生长习性可能导致两种植物在资源和空间上产生竞争作用。同样在小麦/玉米间作体系中小麦处于竞争优势,而玉米处于竞争劣势[21]。除此之外,稗草的竞争优势还有可能跟本试验的种植时间有关。大田条件下往往是玉米出苗后杂草才开始萌发生长,而在本试验中两种植物的出苗时间相当,因此稗草在生长初期时就已经表现出竞争优势地位。在玉米/蚕豆间作体系中,两者生物量均增加(表3),说明玉米和蚕豆之间存在互惠作用,这一结果与目前的大多数研究结果相符[22]。
3.2菌根效应
从表1可以看出,AMF对三种植物的生长均产生了显著的影响,但是三种植物对菌根的响应程度差异很大,玉米响应最为强烈,其次是稗草,最弱的是蚕豆(表3)。大量的研究同样表明,玉米是一种对菌根依赖性高的作物[23-24],说明AMF对玉米的生长起着关键作用。最近的研究报道显示,在与玉米间作的情况下, AMF能有效抑制田间杂草(稗草)的生长[7]。这与本试验结果相反。其原因可能是由于本试验中植物生长基质中有效磷含量较低(低于3 mg/kg),而低磷水平促进了植物与AMF之间共生关系的形成。此外,植物对AMF的依赖性与植物的基因型密切相关,同种植物不同品系对AMF的响应存在差异,不同地区的AMF对同种植物的影响也存在差异。与玉米和稗草相比,AMF对蚕豆的促进作用最小。这种现象可能是由于种植时间较短造成的。相比玉米和稗草,蚕豆种子比较大,蚕豆苗期生长所需要的养分只要来源于种子营养,AM真菌对蚕豆生长的调节作用可能体现在蚕豆生长的后期。
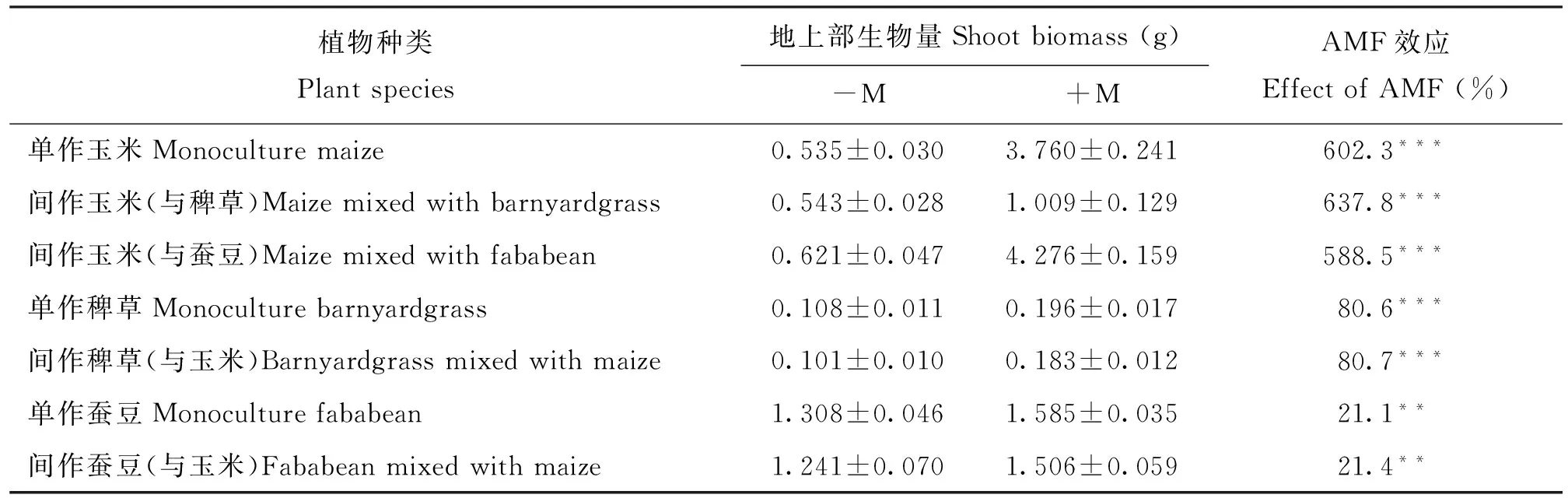
表3 AMF对3种植物地上部生物量的影响
注(Note): 星号表示经过独立样本t检验-M与+M处理下植物地上部生物量之间存在显著性差异(**—P≤ 0.01; ***—P≤0.001)Asterisk means significant difference in shoot biomass of -M and +M after independent t-test (**—P≤ 0.01; ***—P≤0.001).
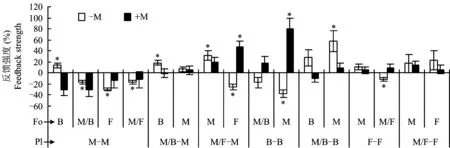
图1 玉米、稗草和蚕豆的菌根反馈作用强度Fig. 1 Feedback strength of inoculated and un-inoculated maize, barnyardgrass and fababean
注(Note):Fo—外来土壤类型 Foreign soil types; Pl—植物类型Plnat species; M-M—单作玉米Maize-Maize; M/B-M—玉米/稗草 间作中的玉米Maize/Barnyardgrass-Maize. 种植类型M星号表示经过单样本t检验后反馈强度与数值“0”之间存在显著性差异(P≤ 0.05) Asterisk means significant difference in feedback strength and “0” after one-samplet-test (P≤ 0.05).]
3.3AMF的反馈调节作用
单作玉米以负反馈作用为主(图1),说明即使在短期培养条件下,玉米也能出现连作障碍。通常连作障碍源于土壤理化性质的改变、自毒作用以及土壤微生物[25]。本试验反馈阶段的培养基质10%来自上茬植物,90%为背景基质,因此土壤理化性质可能不是影响玉米生长的主要限制因子。土壤来源和AMF的交互作用可以显著影响玉米/蚕豆体系中玉米的生长(表1),在接种AMF后,玉米的负反馈减弱或者消失(图1),说明AMF与玉米根系分泌物之间存在相互作用,缓解了非生物因素带来的负反馈作用,减轻了玉米的连作障碍。
而在-M处理中,当外来植物为稗草时,玉米出现正反馈作用,而蚕豆对玉米产生强烈的负反馈作用(图1),说明在不接种AMF的情况下,尽管驯化土壤的接种量仅为培养基质的10%,稗草在土壤中的根系或者分泌物的残留物抑制了玉米生长,而蚕豆的残留物能促进玉米生长。
与单作玉米不同,除外来植物为单作蚕豆外,间作玉米表现为零到正反馈作用(图1),说明玉米与其它植物的间作可能改变了土壤中的非生物(根系分泌物)或者生物因素(微生物的群落结构),从而有效克服玉米的连作障碍。根系分泌物在植物与土壤微生物相互作用的早期发挥着重要作用[26-27]。此外,AMF与宿主植物之间存在一定的宿主偏好性,也会导致土壤中的AMF群落产生显著性的差异[9]。研究发现,土壤环境中的非生物因素与生物因素之间的相互作用能显著影响植物生长。例如,Hage-Ahmed 等[1]发现接种AMF处理番茄根系分泌物中的绿原酸能有效抑制尖孢镰刀菌的发病率。因此不同种植体系中根系分泌物的差异可能引起土壤中AMF群落结构的差异,而正是这种差异缓解了玉米连作的生长抑制作用。
稗草的反馈作用以从零到正反馈作用为主(图1)。一方面说明稗草偏爱本地土壤,另一方面也可能外来土壤(玉米驯化土壤)对稗草的生长存在抑制作用。大量的研究表明,稗草会诱导水稻和其它植物分泌一些化感物质,进而抑制稗草的生长[28]。接种AMF后单作稗草的正反馈作用加强,表明稗草更喜欢本地的AMF群落。研究表明,通过改变根的养分资源的分配比例,植物可以抑制或者优先促进某种共生菌的生长,以选择对自身生长更有益的共生微生物群落[29-30]。
蚕豆的反馈作用主要从零到正反馈(图1),表明蚕豆更加倾向于本地驯化的土壤环境,即蚕豆的残留物不仅对玉米的生长有积极的影响,而且还不影响蚕豆自身的生长,因此玉米/蚕豆间作体系间作生物量的增加主要源于蚕豆对玉米生长的促进作用。研究表明,豆科作物根际磷酸酶的活性显著高于玉米,能够促进与其间作的玉米对土壤磷的吸收[31]。此外,豆科作物能够分泌以柠檬酸为主的有机酸来活化土壤中难溶性的磷供给蚕豆自身和玉米吸收[32]。
4 结论
1)在驯化阶段,玉米和稗草之间是竞争关系,间作中玉米处于竞争弱势,与单作相比,其生物量减少了64.0%,稗草处于竞争优势,相比于单作生物量增加了47.8%; 玉米和蚕豆之间是互惠关系,与单作相比,两种作物生物量增量分别达21.7%和38.3%。
2)AMF均促进了三种植物的生长,作用大小依次为玉米>稗草>单作蚕豆。
3)在反馈阶段,玉米连作生物量明显下降。蚕豆的残留物促进了玉米的生长,且未影响蚕豆自身的生长,是玉米/蚕豆体系生物量增加的主要原因。与玉米和蚕豆相比,稗草在自身驯化土壤上生长较好。
4)单作玉米以负反馈作用为主,间作玉米表现为零到正反馈; 蚕豆无论单作还是间作均以零反馈为主; 接种AMF或与其它植物间作后均减弱了玉米的负反馈作用,表明丛枝菌根真菌减弱单作玉米的负反馈实现间作增产。
本研究以两种不同类型的农田间作体系为研究对象,利用植物-土壤反馈理论来理解AMF对间作优势的调节作用,不仅充实了植物-土壤反馈理论,而且为揭示间作高产高效的机理提供了一个新的研究视角。合适的作物间套作可以通过充分挖掘和发挥土壤有益微生物的作用,提高资源利用效率。
[1]Hage-Ahmed K, Moyses A, Voglgruber A,etal. Alterations in root exudation of intercropped tomato mediated by the arbuscular mycorrhizal fungusGlomusmosseaeand the soilborne pathogenFusariumoxysporumf. sp.Lycopersici[J]. Journal of Phytopathology, 2013, 161(11-12): 763-773.
[2]Lacombe S, Bradley R L, Hamel C, Beaulieu C. Do tree-based intercropping systems increase the diversity and stability of soil microbial communities?[J]. Agriculture Ecosystems and Environment, 2009, 131(1-2): 25-31.
[3]Bever J D. Feedback between plants and their soil communities in an old field community[J]. Ecology, 1994, 75(7): 1965-1977.
[4]Van der Heijden M G A, Klironomos J N, Ursic M,etal. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity[J]. Nature, 1998, 396(6706): 69-72.
[5]Bennett A E, Bever J D. Trade-offs between arbuscular mycorrhizal fungal competitive ability and host growth promotion inPlantagolanceolata[J]. Oecologia, 2009, 160(4): 807-816.
[6]Mangan S A, Herre E A, Bever J D. Specificity between neotropical tree seedlings and their fungal mutualists leads to plant-soil feedback[J]. Ecology, 2010, 91(9): 2594-2603.
[7]Veiga R S, Jansa J, Frossard E, van der Heijden M G. Can arbuscular mycorrhizal fungi reduce the growth of agricultural weeds?[J]. PLoS One, 2011, 6(12): e27825.
[8]Hage-Ahmed K, Krammer J, Steinkellner S. The intercropping partner affects arbuscular mycorrhizal fungi andFusariumoxysporumf. sp.lycopersiciinteractions in tomato[J]. Mycorrhiza, 2013, 23(7): 543-550.
[9]Bever J D. Negative feedback within a mutualism: host-specific growth of mycorrhizal fungi reduces plant benefit[J]. Proceedings of The Royal Society. Biological Sciences, 2002, 269(1509): 2595-2601.
[10]Bever J D, Morton J B, Antonovics J, Schultz P A. Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland[J]. Journal of Ecology, 1996, 84(1): 71-82.
[11]Horwith B. A role for intercropping in modern agriculture[J]. Bioscience, 1985, 35(5): 286-291.
[12]Bever J D. Soil community feedback and the coexistence of competitors: conceptual frameworks and empirical tests[J]. New Phytologist, 2003, 157(3): 465-473.
[13]Wardle D A, Bardgett R D, Klironomos J N,etal. Ecological linkages between aboveground and belowground biota[J]. Science, 2004, 304(5677): 1629-1633.
[14]Van der Putten W H, Bardgett R D, Bever J D, Cobbold S M. Plant-soil feedbacks: the past, the present and future challenges[J]. Journal of Ecologist., 2013, 101(2): 265-276.
[15]Bainard L D, Koch A M, Gordon A M, Klironomos J N. Temporal and compositional differences of arbuscular mycorrhizal fungal communities in conventional monocropping and tree-based intercropping systems[J]. Soil Biology and Biochemistry, 2012, (45): 172-180.
[16]Sheng P P, Liu R J, Li M. Inoculation with an arbuscular mycorrhizal fungus and intercropping with pepper can improve soil quality and watermelon crop performance in a system previously managed by monoculture[J]. American-Eurasian Journal of Agricultural and Environmental Sciences, 2012, 12(11): 1462-1468.
[17]Dania S O, Fagbola O, Dania M I. Response of maize-pigeon pea intercrop to arbuscular mycorrhizal fungi in nutrient depleted soil[J]. Annual Review of Materials Research, 2013, 3(3): 232-245.
[18]Li Q Z, Sun J H, Wei X J,etal. Overyielding and interspecific interactions mediated by nitrogen fertilization in strip intercropping of maize with faba bean, wheat and barley[J]. Plant and Soil, 2011, 339(1-2): 147-161.
[19]Li L, Zhang F S, Li X L,etal. Interspecific facilitation of nutrient uptake by intercropped maize and faba bean[J]. Nutrient Cycling in Agroecosystems, 2003, 65(1): 61-71.
[20]Kardol P, Cornips N J, van Kempen M M L,etal. Microbe-mediated plant-soil feedback causes historical contingency effects in plant community assembly[J]. Ecological Monographs, 2007, 77(2): 147-162.
[21]Li L, Sun J H, Zhang F S,etal. Wheat/maize or wheat/soybean strip intercropping I. Yield advantage and interspecific interactions on nutrients[J]. Field Crops Research, 2001, 71(2): 123-137.
[22]Li Y, Ran W, Zhang R,etal. Facilitated legume nodulation, phosphate uptake and nitrogen transfer by arbuscular inoculation in an upland rice and mung bean intercropping system[J]. Plant and Soil, 2009, 315(1-2): 285-296.
[23]姚青, 冯固, 李晓林. 不同作物对VA菌根真菌的依赖性差异[J]. 作物学报, 2000, 26(6): 874-878.
Yao Q, Feng G, Li X L. Variation between mycorrihizal dependency of different crops[J]. Acta Agronomica Sinica, 2000, 26(6): 874-878.
[24]李侠, 张俊伶. 丛枝菌根根外菌丝对铵态氮和硝态氮吸收能力的比较[J]. 植物营养与肥料学报, 2009, 15(3): 683-689.
Li X, Zhang J L. Uptake of ammonium and nitrate by external hyphae of abuscular mycorrhizal fungi[J]. Plant Nutrition and Fertilizer Science, 2009, 15(3): 683-689.
[25]Yu J, Komada H, Yokoyama H,etal. Sugi (CryptomeriajaponicaD. Don) bark, a potential growth substrate for soilless culture with bioactivity against some soilborne diseases[J]. Journal of Horticultural Science, 1997, 72(6): 989-996.
[26]Bertin C, Yang X, Weston L A. The role of root exudates and allelochemicals in the rhizosphere[J]. Plant and Soil, 2003, 256(1): 67-83.
[27]Bais H P, Weir T L, Perry L G,etal. The role of root exudates in rhizosphere interactions with plants and other organisms[J]. Annual Review of Plant Biology, 2006, 57: 233-266.
[28]孔垂华, 徐效华, 梁文举, 等. 水稻化感品种根分泌物中非酚酸类化感物质的鉴定与抑草活性[J]. 生态学报, 2004, 24(7): 1317-1322.
Kong C H, Xu X H, Liang W J,etal. Non-phenolic alleochemicals in root exudates of an allelopathic rice variety and their identification and weed-suppressive activity[J]. Acta Ecologica Sinica, 2004, 24(7): 1317-1322.
[29]Kiers E T, Rousseau R A, West S, Denison R F. Host sanctions and the legume-rhizobium mutualism[J]. Nature, 2003, 425(6953): 78-81.
[30]Bever J D, Richardson S C, Lawrence B M,etal. Preferential allocation to beneficial symbiont with spatial structure maintains mycorrhizal mutualism[J]. Ecology Letters, 2009, 12(1): 13-21.
[31]汤小群. 间作豆科作物根际过程促进玉米磷吸收的机理研究[D]. 北京: 中国农业大学博士论文, 2008.
Tang X Q. Legume rhizosphere processes facilitate phosphorus upttake by maize in intercropping[D]. Beijing: PhD Dissertation of China Agricultural University, 2008.
[32]Gerke J, Beiβner L, Römer W. The quantitative effect of chemical phosphate mobilization by carboxylate anions on P uptake by a single root. I. The basic concept and determination of soil parameters[J]. Journal of Plant Nutrition and Soil Science, 2000, 163(2): 207-212.
Feedback of abuscular mycorrhizal fungi on interactions between maize/fababean and maize/barnyardgrass
XIE Xian-jin1, 2, ZHANG Jun-ling1 *
(1CollegeofResourcesandEnvironmentalSciences,ChinaAgriculturalUniversity,Beijing100193,China;2HenanSoilandFertilizerStation,Zhengzhou450002,China)
【Objectives】 Arbuscular mycorrhizal fungi (AMF) plays an important role in contributing to plant growth in agro-ecosystems. Feedback of AMF communities pre-trained by different host plants and cropping types may influence the growth of subsequent plants, and intercropping system contributes to increase in yield and resource use efficiency. Therefore, understanding on feedback of AMF has a positive significance in elucidating yield increase in intercropping systems.【Methods】 Three isolates of AM fungi ofFunneliformismosseae,RhizophagusintraradicesandClaroideoglomusclaroideum, three plant species of maize (ZeamaysL.), fababean (ViciafabaL.) and barnyardgrass (Echinochloacrus-galliL.), and two intercropping systems (facilitation system of maize/fababean and competition system of maize/barnyardgrass) were established to investigate the feedback effects at the phases of two plant training and feedback.【Results】 Data showed that shoot biomass of maize decreased by 64.0% during training phase compared to monoculture in maize/barnyardgrass intercropping system, and the later increased shoot biomass of barnyardgrass by 47.8%. Maize and fababean respectively enhanced the shoot biomass of the plants by 21.7% and 38.3% based on the comparison between double plant cropping and monocropping. During the feedback phase, shoot biomass of maize, barnyardgrass and fababean increased by 602.3%, 80.6% and 21.1% respectively in monoculture system after inoculated with AMFs. In intercropping system, shoot biomass of maize increased by 613.1%, and shoot biomass of barnyardgrass by 80.7%, and shoot biomass of fababean by 21.4%. Monoculture maize exhibited a negative feedback, and the inoculation of AMF and intercropping with barnyardgrass or fababean relieved negative feedback in monocropping maize.【Conclusions】 The competition dominated in maize/barnyardgrass intercropping system compared to monoculture. Facilitation between maize and fababean enhanced the biomass of the respective plants. AMF inoculation increased the biomass of maize, followed by those of barnyardgrass and fababean. The inoculation of AMF, and intercropping with barnyardgrass or fababean relieved negative feedback of monocropping maize. In conclusion, AMF can relive negative feedback of maize and are attributed to the facilitation of maize/fababean intercropping system.
arbuscular mycorrhizal fungi; AMF; plant-soil feedback; maize; fababean; barnyardgrass
2014-09-10接受日期: 2014-11-25网络出版日期: 2015-04-21
国家自然科学基金重大国际合作项目(31210103906); 国家自然科学基金委面上项目(31071872和31272251)资助。
谢先进(1989—),男,河南商丘人,硕士研究生,主要从事菌根生态学方面的研究。E-mail: xie_xj2881@sina.com
E-mail: junlingz@cau.edu.cn
S154.3; Q945.1
A
1008-505X(2016)03-0643-07
猜你喜欢
经济林研究(2022年4期)2023-01-12
辽宁林业科技(2022年2期)2022-11-24
广西林业科学(2022年2期)2022-05-09
北方果树(2022年1期)2022-01-21
食药用菌(2021年4期)2021-01-08
河北果树(2020年4期)2020-11-26
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
干旱地区农业研究(2017年5期)2017-12-18
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
