
古典H1N1亚型猪流感病毒PB2 K627E突变对病毒复制能力的影响
2016-07-12 01:49汪秀会宫晓倩阮宝阳刘晓敏李泽君周艳君童光志
畜牧兽医学报 2016年3期
汪秀会,宫晓倩,阮宝阳,2,刘晓敏,汪 琪,张 鹏,2,李泽君,周艳君,童 武,郑 浩,童光志*,于 海*
(1.中国农业科学院上海兽医研究所,上海 200241;2.山东农业大学动物科技学院,泰安 271000)
古典H1N1亚型猪流感病毒PB2 K627E突变对病毒复制能力的影响
汪秀会1,宫晓倩1,阮宝阳1,2,刘晓敏1,汪琪1,张鹏1,2,李泽君1,周艳君1,童武1,郑浩1,童光志1*,于海1*
(1.中国农业科学院上海兽医研究所,上海 200241;2.山东农业大学动物科技学院,泰安 271000)
摘要:旨在探索古典H1N1亚型猪流感病毒A/Swine/Guangdong/1/2011(H1N1) PB2 K627E突变对病毒复制能力的影响。利用反向遗传操作技术和点突变技术成功获得古典H1N1亚型猪流感病毒的拯救株(627K)和突变株(627E)后,分别在MDCK细胞和小鼠体内对拯救株和突变株的复制能力进行比较。体外MDCK细胞试验结果显示,突变株与拯救株相比,细胞病变(CPE)减小、空斑形态减小、生长曲线差异极其显著,表明突变株在MDCK细胞上的复制能力减弱。小鼠攻毒试验结果表明,突变株对小鼠体重变化的影响不明显,在小鼠肺中的病毒滴度明显下降,引起病理损伤较轻。体外MDCK细胞及小鼠攻毒试验结果均表明,突变株与拯救株相比复制能力降低。该结果不仅证实了猪流感病毒 PB2 627位氨基酸是一种重要的毒力分子标记,同时还为进一步阐明其致病机制提供了条件。
关键词:古典H1N1亚型猪流感病毒;PB2 K627E突变;体外MDCK细胞试验;小鼠攻毒试验;复制能力
猪流感(swine influenza,SI)是由猪流感病毒(swine influenza virus,SIV)引起的以咳嗽、流涕、喷嚏、高烧、昏睡、呼吸困难和食欲下降等[1]为主要特征的猪的一种常见病。该病一年四季均可发生,并可继发或同时感染猪繁殖与呼吸障碍综合征、猪伪狂犬、流行性腹泻等[2],给养猪业造成了巨大的经济损失。由于在猪的呼吸道上皮细胞同时具有禽流感病毒受体SAα-2,3-Gal和人流感病毒受体SAα-2,6-Gal,猪可以同时被禽流感病毒、猪流感病毒和人流感病毒感染。因此,猪被认为是禽流感病毒和人流感病毒的中间宿主和发生基因重组的“混合器”[3]。猪对流感病毒的生态分布和遗传演变起着举足轻重的作用,成为流感病毒在“禽-猪-人”种间传播链中的重要环节。目前,在世界范围内已从猪体内分离出的流感病毒约有14种不同的亚型,其中主要为H1N1、H3N2和 H1N2三种[4-6]。
导致流感病毒致病性的因素很多,病毒蛋白在其中扮演着不可忽视的角色,其中的PB2 627位氨基酸被公认为流感病毒的一种重要的毒力分子标记[7-8],且具有宿主特异性[8-9]。研究发现,不同来源的流感病毒PB2第627位所编码的氨基酸不同。分离得到的禽流感病毒的PB2 627位氨基酸通常为谷氨酸(E),人源或猪源流感病毒该位点大多数为赖氨酸(K),当该位点编码的氨基酸由E突变为K后,病毒的聚合酶活性会明显升高,病毒在细胞内的复制能力也明显增强。当将K突变为E后,病毒的复制能力及致病性明显减弱,在体内及体外试验中的毒力明显降低[10]。本研究中所选用的毒株为古典H1N1亚型SIV,其PB2 627位编码的氨基酸为K,为了探索该位点的氨基酸由K突变为E后,其致病性及在体内外复制能力会发生怎样的变化,作者进行了一系列试验。
1材料与方法
1.1毒株、菌种及细胞
古典H1N1亚型SIV A/Swine/Guangdong/1/2011(H1N1)(简称G11)的八质粒反向遗传系统由作者实验室构建并保存。大肠杆菌DH5α感受态细胞购于北京天根生物有限公司。293T 细胞,MDCK细胞由本实验室保存。
1.2 主要试剂
4%多聚甲醛,低熔点琼脂糖,TPCK,5%结晶紫,0.5%鸡红细胞,PBS,DMEM培养基、FBS、BSA、2×MEM均购自GIBCO 公司(美国)。抗生素由作者实验室保存。
1.3主要仪器
电子天平,组织破碎仪,Nikon TS100 倒置显微镜(Nikon公司),生物安全柜(Forma-scientific 公司),二氧化碳培养箱(Forma-scientific 公司),eppendorf 8道和12道排枪,乙醚,50 mL离心管,低温水平离心机。
1.4拯救株病毒和突变株病毒的获得
将G11反向遗传系统的八个阳性质粒分别按照1 μg量共转染293T细胞,6 h后换液,48 h后吸取上清接种MDCK细胞,待细胞出现明显病变时,收集病毒,提取RNA,反转录,进行序列测定,相应病毒命名为拯救株。在突变株病毒的拯救过程中,首先要设计一对反向互补的内部引物,这对引物的作用是使PB2基因的1 879—1 881位由AAG突变为GAG;通过融合 RT-PCR方法获得627位氨基酸由K突变为E的突变PB2基因,进而构建它的阳性质粒;将G11原始毒株的PB1、PA、HA、NA、NP、M、NS阳性质粒和突变PB2基因的阳性质粒共转染293T细胞,进行病毒的拯救,最后也进行序列测定,相应病毒命名为突变株。
1.5细胞病变的观察
将拯救株和突变株病毒以同种剂量分别接种到单层铺满的MDCK细胞的6孔细胞培养板中,在不同时间点(6、12、24、36、48、60、72 h)观察CPE情况。
1.6空斑试验
拯救株和突变株病毒分别用含TPCK的感染液依次稀释至10-6,然后分别接种于单层铺满MDCK细胞的6孔细胞培养板中,充分吸附1 h后,弃去病毒液,每孔加入4 mL配制好的胶(含LMP、MEM、BSA和TPCK等),待胶凝固后,倒置于37 ℃、5%CO2培养箱中培养,每隔一段时间观察空斑的有无及大小形态变化。待空斑形成大小适中(48~72 h)时,5%龙胆紫染色。
1.7生长曲线的绘制
拯救株与突变株分别以0.001MOI接种于单层铺满且状态良好的MDCK细胞中,分别在不同的时间点(6、12、24、36、48、60、72 h)收集细胞上清,每个时间点设3个重复。将每个时间点收集到的每个重复视作一个单独的样品,将每个样品由10-1依次稀释至10-12后接种于MDCK细胞,置于37 ℃培养箱,72 h后在显微镜下观察细胞的生长情况,并结合血凝方法进行检测。
1.8小鼠体重变化及存活率
将拯救株与突变株分别以1.0×105TCID50的攻毒剂量滴鼻接种小鼠,每天同一时间点进行连续称重两周,观察并记录小鼠体重变化及存活情况,绘制小鼠的体重变化曲线和存活率曲线。
1.9小鼠肺组织病理学观察
分别取攻毒后第3天和第5天的试验组及对照组的小鼠左肺组织,置于注满4%多聚甲醛的5 mL离心管中,做好标记后送公司制备组织病理切片,进行组织病理学观察。
1.10小鼠肺病毒滴度的检测
分别取攻毒后第3天和第5天的试验组和对照组小鼠右肺组织,充分研磨后离心取上清,在无菌环境下倍比稀释至10-12,接种MDCK细胞,进行病毒滴度的测定。对照组小鼠接种的是同种剂量的DMEM,具体操作流程与试验组小鼠一致。
2结果
2.1古典H1N1亚型SIV G11拯救株及突变株序列分析
利用反向遗传技术及点突变技术,成功获得了古典H1N1亚型SIV G11拯救株及突变株病毒,提取病毒RNA,进行RT-PCR扩增及序列测定,序列分析结果表明,拯救株的PB2 627位氨基酸为K,而突变株的PB2 627位氨基酸为E,进而证明成功获得了古典H1N1亚型猪流感病毒的突变株。
2.2不同时间点MDCK细胞形态
不同时间点MDCK细胞形态的观察结果(图1)表明:拯救株与突变株在12 h时均没有肉眼可见的CPE;24 h时拯救株出现CPE,突变株的病变仍然不明显;36 h时,突变株细胞出现肉眼可见的病变,拯救株CPE较明显;48 h时,拯救株细胞约有50%发生病变,突变株CPE程度轻于拯救株;60 h时,拯救株与突变株的病变程度均很明显,但突变株较拯救株相比程度较轻;72 h时,突变株与拯救株细胞完全脱落。
2.3空斑形态
在相同条件下突变株与拯救株分别接种于MDCK细胞,观察两毒株在MDCK细胞上产生空斑形态的大小。空斑形态观察结果(图2)表明,PB2 K627E突变导致古典H1N1亚型SIV在MDCK细胞上的空斑形态变小,含PB2 627E位点的突变株的空斑明显小于含PB2 627K位点的拯救株,说明突变株在MDCK细胞上的复制能力弱于拯救株。
2.4拯救株与突变株生长曲线
古典H1N1亚型猪流感病毒拯救株和突变株分别以相同剂量接种MDCK细胞,测定病毒滴度(TCID50),利用GraphPad Primer 5.0软件对各时间点的病毒滴度进行差异性分析,结果(图3)表明:12 h时突变株与拯救株的病毒滴度差异不显著(P>0.05),24、36、48 h时突变株与拯救株的病毒滴度差异极其显著(P<0.001),60、72 h差异非常显著(P<0.01),表明突变株在细胞上的复制能力较弱。
2.5小鼠肺病理切片的观察
分别取攻毒后第3天和第5天左肺组织,制作病理切片,HE染色,由图4可知,拯救株(A、B)具有明显的炎性细胞灶性浸润(图中a黑色箭头),在同一视野下可见多个浸润灶,部分肺泡壁增粗(图中b红色箭头);突变株(C、D)局部炎性细胞灶性浸润,炎性细胞较少,肺泡结构正常,肺泡壁结构正常,支气管结构正常。病理切片结果表明,攻毒后第3天和第5天,突变株与拯救株相比,突变株对小鼠肺组织损伤较轻。
2.6小鼠体重变化及存活率
由图5和图6可以看出:突变株(627E)组与对照组小鼠体重变化差异不显著(P>0.05),说明突变株在小鼠体内的复制能力较弱,导致小鼠体重变化不明显;病毒感染小鼠后第3天至第13天(dpi 3—dpi 13),与突变株组和对照组小鼠体重变化情况相比,拯救株接种小鼠体重有明显的下降,差异极其显著(P<0.001),表明拯救株在小鼠体内的复制能力较强;而拯救株在使小鼠体重明显下降的同时对小鼠具有致死性,导致小鼠部分死亡。

A.拯救株(rG11)在MDCK细胞不同时间点(12、24、36、48、60、72 h)的CPE;B.突变株(rG11-K627E)在MDCK细胞不同时间点(12、24、36、48、60、72 h)的CPEA.CPE of rescued strain in MDCK at different time;B.CPE of and mutant strain in MDCK at different time图1 H1N1亚型猪流感病毒G11拯救株与突变株6个时间点MDCK CPE(400×)Fig.1 CPE of rescued and mutant strain of G11 at different time(400×)
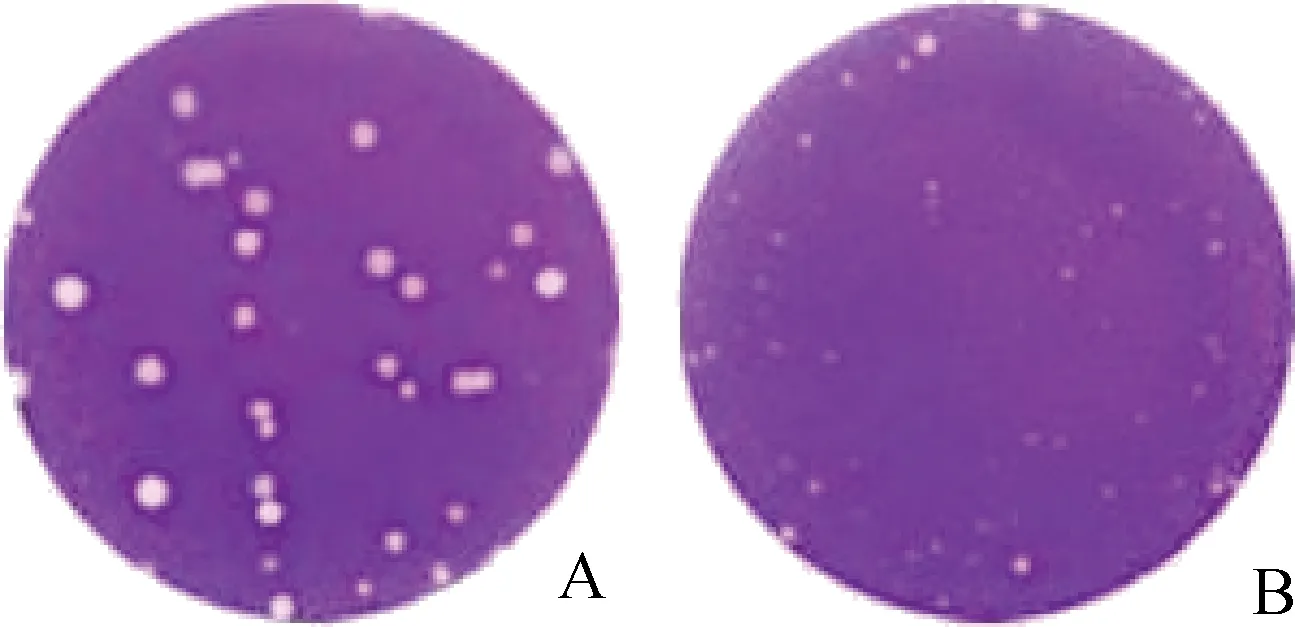
A.拯救株空斑形态;B.突变株空斑形态A.Plaque morphology of rescued strain;B.Plaque morphology of mutant strain图2 G11拯救株与突变株空斑形态的观察和比较Fig.2 The observation and comparison of plaque morphology of rescued and mutant strain
图3 G11拯救株与突变株生长曲线的比较Fig.3 The growth curve of the rescued and mutant strain
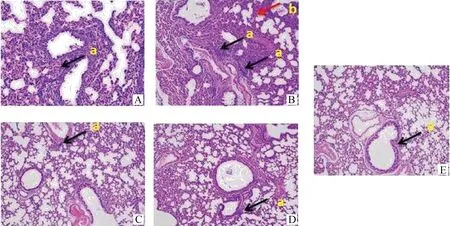
A.rG11-3 d病理切片;B.rG11-5 d病理切片;C.G11-K627E-3 d病理切片;D.G11-K627E-5 d病理切片;E.对照组;a.炎性细胞灶性浸润; b.肺泡壁增粗A.rG11-3 d lung pathological;B.rG11-5 d lung pathological;C.G11-K627E-3 d lung pathological;D.G11-K627E-5 d lung pathological;E.Control;a.Focal infiltration of inflammatory cell;b.Thickening of alveolar walls图4 第3、5天小鼠左肺组织拯救株及突变株病理切片的观察(200×)Fig.4 The result of the rescued and mutant strains’ left lung pathological(200×)
2.7病毒滴度
利用GraphPad Primer 5.0软件,对攻毒后小鼠肺组织的病毒滴度(TCID50)进行差异性分析,结果(图7)表明:攻毒后第3、5天小鼠右肺组织拯救株和突变株病毒滴度差异极显著(P<0.001),说明突变株与拯救株相比在小鼠肺的复制能力差异显著,且突变株的复制能力明显低于拯救株,而对照组小鼠未检测到病毒。
图5 G11拯救株与突变株小鼠体重变化情况Fig.5 Body weight changes of mice of the rescued and mutant strain
图6 拯救株及突变株病毒感染小鼠存活率Fig.6 Survival rate of the mice infected by rescued and mutant strain
图7 第3、5天小鼠右肺组织拯救株及突变株病毒的滴度检测结果Fig.7 The result of the rescued and mutant strains’ titer of mice’ right lung
3讨论
本研究所选用的古典H1N1亚型SIV拯救株的PB2 627位编码的氨基酸为K,利用点突变技术将该位点的氨基酸突变为E后,分别在MDCK细胞和小鼠体内对突变前后病毒的复制能力进行了比较。MDCK细胞试验中,分别在CPE、空斑形态、病毒滴度、生长曲线等几方面进行其生物学特性的比较,结果表明突变株与拯救株间在体外的生物学特性差异明显。与拯救株相比,突变株的CPE较小;在完全相同条件下,突变株所产生的空斑的形态较小;且突变株与拯救株相比,其病毒滴度较低,生长曲线差异极其显著。同时,本研究还以BALB/c雌性小鼠为哺乳动物模型,为探索拯救株与突变株在体内条件下的差异提供了条件。在小鼠攻毒试验中,分别在小鼠体重变化、小鼠肺病毒滴度、小鼠肺病理切片等方面对拯救株与突变株生物学特性间的差异进行了比较。试验结果表明,拯救株能够导致小鼠体重明显下降甚至死亡,而突变株使小鼠体重下降不明显,攻毒至第13天,小鼠的适应性增强,体重恢复并不断增加;小鼠肺病毒滴度差异极其显著;肺病理切片的结果表明两毒株对小鼠致病性差异不显著,但突变株与拯救株相比致病性较弱。从而表明PB2 627K是影响古典H1N1亚型SIV复制能力的重要因素之一 。
研究发现,人流感病毒PB2 627位氨基酸为K而禽源流感病毒该位点为E[11]。分离于北美的SIV,于1998年之前分离得到的大多数PB2 627位氨基酸为K,而1998年之后分离的绝大多数为E[12]。禽源流感病毒PB2 627位氨基酸为E,但病毒仍具有较强的感染性,究其原因是该位点与其他位点的相互作用[13]有关。SIV PB2 627位氨基酸由K突变为E后,病毒的复制能力及致病性明显降低,在体内及体外试验中的毒力明显减弱,究其原因,这可能是由于PB2 627位氨基酸影响着病毒RNA复制的温度敏感性决定的[14]。不同来源的流感病毒复制时所需的最适宜的温度不同,人/猪流感病毒在上呼吸道复制,最适宜温度约33 ℃,而禽流感病毒在禽类肠道中复制的最适宜的温度为41 ℃左右。也就是说,当PB2 627为E 时,该病毒在哺乳动物的上呼吸道是不能进行复制或者说复制效率很低[15]。有研究报道,PB2 627E之所以对低温敏感,可能是因为低温条件下PB2和NP蛋白之间的相互作用受到了破坏[16],也可能是与细胞内某些限制因子[17]或者某些宿主因子有关[18],具体机制还需要进一步探索。
目前国内外对SIV分子致病机制的研究比较少,主要研究仍然集中于人流感病毒和禽流感病毒。本研究通过对古典H1N1亚型SIV拯救株和突变株在体外和体内条件下生物学特性的比较,解析了古典H1N1亚型SIV在体外及体内的致病机制,弥补了该方面的不足,同时也为古典H1N1亚型SIV PB2 627位氨基酸的分子致病机制提供了一定的参考依据。
参考文献(References):
[1]YU H,ZHANG G H,HUA R H,et al.Isolation and genetic analysis of human origin H1N1 and H3N2 influenza viruses from pigs in China[J].BiochemBiophysResCommun, 2007,356(1):91-96.
[2]ELLIS J.Porcine circovirus-2 and concurrent infections in the field[J].VetMicrobiol, 2004,98(2):159-163.
[4]MA W,VINCENT A L,GRAMER M R,et al.Identification of H2N3 influenza A viruses from swine in the United States[J].ProcNatlAcadSciUSA, 2007,104(52):20949-20954.
[5]陈义祥,蒙雪琼.猪流感病毒在世界范围内的流行情况及公共卫生意义[J].微生物学通报,2008,35(4):582-588.
CHEN Y X,MENG X Q.The epidemiology of swine influenza virus in the world and its public health implication[J].Microbiology, 2008,35(4):582-588.(in Chinese)
[6]唐续,杨焕良,鄢明华,等.一株人源H1N1亚型猪流感病毒的进化分析与生物学特性研究[J].中国预防兽医学报,2012,34(4):270-273,336.
TANG X,YANG H L,YAN M H,et al.Evolution analysis and biological characteristics of a human origin H1N1 subtype swine influenza virus[J].ChineseJournalofPreventiveVeterinaryMedicine, 2012,34(4):270-273,336.(in Chinese)
[7]HATTA M,GAO P,HALFMANN P,et al.Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses[J].Science, 2001,293(5536):1840-1842.
[8]SUBBARAO E K,LONDON W,MURPHY B R.A single amino acid in the PB2 gene of influenza A virus is a determinant of host range[J].JVirol, 1993,67(4):1761-1764.
[9]ALMOND J W.A single gene determines the host range of influenza virus[J].Nature, 1977,270(5638):617-618.
[10]ITOH Y,SHINYA K,KISO M,et al.Invitroandinvivocharacterization of new swine-origin H1N1 influenza viruses[J].Nature,2009,460(7258):1021-1025.
[11]CHEN G W,CHANG S C,MOK C K,et al.Genomic signatures of human versus avian influenza A viruses[J].EmergInfectDis,2006,12(9):1353-1360.
[12]BUSSEY K A,BOUSSE T L,DESMET E A,et al.PB2 residue 271 plays a key role in enhanced polymerase activity of influenza A viruses in mammalian host cells[J].JVirol, 2010,84(9):4395-4406.
[13]LI J,ISHAQ M,PRUDENCE M,et al.Single mutation at the amino acid position 627 of PB2 that leads to increased virulence of an H5N1 avian influenza virus during adaptation in mice can be compensated by multiple mutations at other sites of PB2[J].VirusRes, 2009,144(1-2):123-129.
[14]MASSIN P,VAN DER WERF S,NAFFAKH N.Residue 627 of PB2 is a determinant of cold sensitivity in RNA replication of avian influenza viruses[J].JVirol,2001,75(11):5398-5404.
[15]HATTA M,HATTA Y,KIM J H,et al.Growth of H5N1 influenza A viruses in the upper respiratory tracts of mice[J].PLoSPathog,2007,3(10):1374-1379.[16]RAMEIX-WELTI M A,TOMOIU A,DOS SANTOS AFONSO E,et al.Avian influenza A virus polymerase association with nucleoprotein,but not polymerase assembly,is impaired in human cells during the course of infection[J].JVirol,2009,83(3):1320-1331.
[17]MEHLE A,DOUDNA J A.An inhibitory activity in human cells restricts the function of an avian-like influenza virus polymerase[J].CellHostMicrobe,2008,4(2):111-122.
[18]MONCORGÉ O,MURA M,BARCLAY W S.Evidence for avian and human host cell factors that affect the activity of influenza virus polymerase[J].JVirol,2010,84(19):9978-9986.
(编辑白永平)
The Effect of PB2 K627E Mutation on the Replication Competence of Classical H1N1 Subtype Swine Influenza Virus
WANG Xiu-hui1,GONG Xiao-qian1,RUAN Bao-yang1,2,LIU Xiao-min1,WANG Qi1,ZHANG Peng1,2,LI Ze-jun1,ZHOU Yan-jun1,TONG Wu1,ZHENG Hao1,TONG Guang-zhi1*,YU Hai1*
(1.ShanghaiVeterinaryResearchInstitute,ChineseAcademyofAgriculturalSciences,Shanghai200241,China;2.CollegeofVeterinaryMedicine,ShandongAgriculturalUniversity,Tai’an271000,China)
Abstract:This experiment was conducted to explore the effect of PB2 K627E mutation on the replication competence of classical H1N1 subtype swine influenza virus(A/Swine/Guangdong/1/2011).The rescued and mutant strains of classical H1N1 subtype swine influenza virus were obtained by reverse genetics and point mutation technology,and researches were taken on MDCK cells and mice.The results on MDCK cells showed that,compared with the rescued strain,the mutant virus had a slight degree of CPE;the morphology of plaque of mutant virus was much smaller;the growth curve showed that it had a significant distinct difference.The results of experiments in mice showed that the mutant strain had a low pathogenicity and could not lead the mice loss weight,and virus titer in lung tissues on the 3rd and 5th day post challenge showed that there was a significant distinct difference between the two strains and the pathological damage caused by mutant one was not serious.The results on MDCK and mice both showed that the mutant strain’s replication competence was lower than the rescued one.This conclusion demonstrates that the amino acid of PB2 627 is not only one of the important virulence markers,but also provided the condition to further clarify its pathogenic mechanism.
Key words:classical H1N1 subtype swine influenza;PB2 K627E mutation;vitroexperiment in MDCK cells;vivoexperiment in mice;virus replication
doi:10.11843/j.issn.0366-6964.2016.03.017
收稿日期:2015-08-04
基金项目:国家青年自然科学基金(31201916);上海市自然科学基金青年项目(12ZR1453500);中央级公益性科研院所基本科研业务费项目(2015JB07);中国农业科学院创新工程科研团队“动物流感病毒病原生态学”项目
作者简介:汪秀会(1988- ),女,山东聊城人,硕士生,主要从事猪流感病毒分子致病机制的研究,E-mail:wangxiuhui123@163.com *通信作者:童光志,研究员,E-mail:gztong@shvri.ac.cn;于海,副研究员,E-mail:haiyu@shvri.ac.cn
中图分类号:S852.659.5
文献标志码:A
文章编号:0366-6964(2016)03-0543-06
猜你喜欢
临床与实验病理学杂志(2022年5期)2022-08-06
昆明医科大学学报(2022年2期)2022-03-29
华侨大学学报(自然科学版)(2021年4期)2021-07-30
世界科学技术-中医药现代化(2021年12期)2021-04-19
右江医学(2019年6期)2019-08-10
中国兽医杂志(2019年2期)2019-06-25
猪业科学(2018年6期)2018-01-23
现代检验医学杂志(2016年3期)2016-11-15
现代检验医学杂志(2016年1期)2016-11-12
中国兽医杂志(2016年3期)2016-06-22
