滤泡辅助性T细胞在实验性自身免疫性脑脊髓炎发病过程中的作用①
2016-06-29 05:21:58平昌昀李呼伦詹晓霞
中国免疫学杂志 2016年6期
平昌昀 李呼伦 詹晓霞 孙 博
(哈尔滨医科大学神经生物教研室,哈尔滨150081)
滤泡辅助性T细胞在实验性自身免疫性脑脊髓炎发病过程中的作用①
平昌昀李呼伦詹晓霞孙博
(哈尔滨医科大学神经生物教研室,哈尔滨150081)
[摘要]目的:探讨滤泡辅助性T细胞在实验性自身免疫性脑脊髓炎中的作用机制。方法:随机将C57BL/6小鼠分为CFA组、EAE组、anti-ICOSL组和control 组。用流式细胞术检测不同时间点Tfh细胞的变化及T细胞的活化情况,石蜡切片染色观察炎细胞浸润、脱髓鞘及生发中心的改变,ELISA法检测髓鞘蛋白特异性抗体的分泌水平。结果:anti-ICOSL治疗组较EAE对照组临床评分低(P<0.05),炎症浸润减轻,无脱髓鞘病变,Tfh细胞比例降低,但活性不受影响,不能够形成生发中心,并且髓鞘蛋白特异性抗体的分泌减少。结论:滤泡辅助性T细胞通过与B细胞的相互作用促进生发中心形成,分泌抗神经髓鞘蛋白MOG35-55特异性抗体发挥致病性作用。
[关键词]滤泡辅助性T细胞;实验性自身免疫性脑脊髓炎;生发中心
多发性硬化(Multiple sclerosis,MS)是T细胞介导的以中枢神经系统(Central nerve system,CNS)白质脱髓鞘病变为特点的自身免疫性疾病[1-3],患者出现神经症状前的数周或数月,多有疲劳、体重减轻、肌肉和关节隐痛等。MS多次复发后,病人症状逐渐加重,出现无力、僵直、感觉障碍、视力损害和尿失禁[4]。各种非特异性免疫抑制治疗不仅能够引起副作用,如感染、肿瘤及骨髓抑制等,而且无法完全防止MS的复发[5]。因此,特异性MS的新疗法仍是许多研究者努力的方向[6,7]。2000年,研究人员首次提出滤泡辅助性T细胞(T follicular helper cells,Tfh)为主要负责辅助B细胞的T细胞亚群,在刺激B细胞增殖、分化以及免疫球蛋白类别的转换中起着十分重要的作用[8-11],但是Tfh细胞参与自身免疫性疾病的作用机制至今仍未被彻底地阐明和解释。随着研究的深入,越来越多的实验证据表明,B细胞和自身抗体通过多种途径影响着MS的发病和病程转归[12-15]。而作为辅助B细胞的Tfh细胞是通过辅助B细胞还是通过自身的致病性参与多发性硬化、在多发性硬化中Tfh细胞又是如何变化等问题目前仍是空白。实验性自身免疫性脑脊髓炎(Experimental autoimmune encelphalomyelitis,EAE)是目前应用最广泛的MS实验动物模型,本研究通过EAE小鼠模型,探讨Tfh在EAE中的作用机制,旨为临床对MS的治疗提供理论依据。
1材料与方法
1.1材料
1.1.1实验动物SPF级C57BL/6小鼠,雌性,24只,6~8周龄,体重16~18 g,购自北京维通利华实验动物技术有限公司,SCXK(京)2015-0001。对C57BL/6小鼠昼夜12 h 交替光照,自由食水,于室温(20±2)℃条件下饲养。
1.1.2仪器和试剂本研究所用到的仪器为流式细胞计数仪(美国BD公司),Digital Camera(日本HITACHI公司),酶标仪(澳大利亚Bio-Rad公司)。
本研究所用到的试剂为结核菌素(H37RA,美国DIFCO LABORATORIES),不完全弗氏佐剂(IFA,美国Sigma),MOG35-55(西安联美生物科技有限公司),百日咳毒素(PT,List biological laboratories,inc.),ELISA抗体(美国BD公司),anti-ICOSL抗体(由北京博奥森生物技术有限公司合成),HE染色试剂(天津市凯通化学试剂有限公司),LFB染色试剂(杭州临安金龙化工有限公司)。
1.2实验方法
1.2.1动物分组及EAE小鼠模型的建立随机将C57BL/6小鼠分为CFA组:注射完全弗氏佐剂(CFA,250 μg结核菌素+50 μl IFA+50 μl 0.01 mol/L PBS);EAE组;anti-ICOSL组和control 组:均注射CFA和MOG35-55,每组6只。免疫试剂配好后,用漩涡振荡器充分振荡成均匀乳白色液体后,于小鼠两侧腋窝皮下进行注射免疫。免疫当天和免疫后的第2天,EAE组、control组和anti-ICOSL治疗组小鼠尾静脉注射百日咳毒素(PT),200 ng/只;EAE建模的第2、4、6天,anti-ICOSL组小鼠腹腔注射anti-ICOSL抗体,100 μg/次;control 组小鼠腹腔注射与anti-ICOSL 抗体同等剂量的生理盐水。临床评分标准:0分:正常;0.5分:部分尾瘫;1分:尾瘫;1.5分:尾瘫伴蹒跚步态;2分:一侧后肢瘫痪;2.5分:一侧后肢瘫痪伴另一侧后肢无力;3分:双后肢瘫痪;3.5分:上行性瘫痪;4分:躯干无力或瘫痪;4.5:濒死;5分:死亡。
1.2.2T淋巴细胞的提取眼球取血处死发病早期、高峰期、缓解期EAE模型小鼠,无菌取小鼠腋窝、腹股沟、腹主动脉旁淋巴结细胞和脾单核细胞。将淋巴结和脾单核细胞混合,以5×106个/ml细胞的浓度加入1640完全培养液,同时加入MOG35-55至终浓度为25 μg/ml,置37℃、5%CO2培养箱中培养。将经过MOG35-55肽段体外刺激所获得的特异性增殖的淋巴细胞作为实验T淋巴细胞。
1.2.3T淋巴细胞活性及滤泡辅助性T细胞的检测分别采用流式细胞术检测Th细胞(主要为CD3+、CD44+及CD4+的细胞)活性和滤泡辅助性T细胞的表达(Tfh,主要为BCL-6-PE、CXCR5-PE、ICOS-PerCP)。于EAE免疫后3、5、8、15 d时分离淋巴结和脾,研磨过滤,制成淋巴细胞悬液,加入MOG(25 μg/ml)抗原刺激细胞后,于37℃,5% CO2培养箱中进行体外培养。Tfh细胞的表面标志为CD4+ICOS+CXCR5+[16]。
1.2.4组织病理学检测于发病高峰期(第15天)取小鼠脊髓腰骶段,制成石蜡切片,用5 μm厚度的组织切片进行HE染色[17]、8 μm厚度的组织切片进行LFB染色[17],高倍显微镜观察病理改变。取发病高峰期(第15天)小鼠脾脏,制作冰冻切片,用6 μm 厚度的组织切片进行免疫组织化学染色[18],高倍显微镜观察脾组织中生发中心的形成。
1.2.5血清IgG抗体分泌水平的检测采用酶联免疫吸附反应(ELISA)法[19]。
1.3统计学处理采用SPSS11.5软件对相关数据进行统计学处理,实验中任意两独立样本之间的比较用t检验,多个样本均数的比较用单因素方差分析法(One-way classification,ANOVA),以P<0.05为差异有统计学意义。
2结果
2.1EAE小鼠模型的建立试验结果显示,EAE组小鼠从免疫后第10天开始出现临床症状,于免疫后第15天左右达到发病高峰期,免疫后第21天开始缓解。EAE组发病小鼠的临床评分为0.5~3.0分,第11天EAE组评分明显高于CFA组(P<0.05);EAE组在第11天开始出现明显的、逐渐加重的临床症状:尾部无力、蹒跚步态、单后肢至双后肢瘫痪,严重者出现前肢瘫痪;第12、13、14、15天EAE组小鼠EAE评分均高于CFA组(P<0.001)。对照组在相同时间内未出现上述临床症状,活动正常,见图1。
免疫后第15天,进行HE染色,发现与CFA组相比, EAE组脊髓白质有大量的炎性细胞浸润(单核-巨噬细胞,T淋巴细胞,B淋巴细胞,浆细胞等), LFB染色可以观察到EAE组脊髓有明显的脱髓鞘病灶,病灶中髓鞘变性崩解成颗粒状,并被吞噬细胞吞噬,形成泡沫细胞,表明小鼠EAE模型建立成功。见图2。
2.2滤泡辅助性T细胞在淋巴结和脾中的变化情况结果表明:免疫后第5天,淋巴结中的CD4+ICOS+CXCR5+Tfh细胞增多,且Tfh数量达到高峰;EAE组和CFA组相比,差异具有统计学意义(P<0.001)。第8天,Tfh细胞在EAE组所占百分比明显高于CFA组(P<0.001);第15天,Tfh细胞在EAE组所占百分比有所下降,但仍高于对应CFA组,见图3A。免疫后第3、5、8、15天,EAE组小鼠脾中的Tfh细胞数量均高于CFA组,且在第5天时表达最高,与CFA组比较,差异具有统计学意义(P<0.001),见图3B。
2.3选择性滤泡辅助性T细胞阻断剂anti-ICOSL IgG的治疗结果

图1 EAE临床评分及体重变化Fig.1 Clinical scores and weights of EAENote: *.P<0.05,**.P<0.01,***.P<0.001.

图2 脊髓组织HE染色及LFB染色(×200)Fig.2 HE staining and LFB staining of spinal cord(×200)Note: A and B are EAE group:A shows HE staining,in which arrow indicates inflammatory cell infiltration;B shows LFB staining,in which arrows indicate demyelination;C and D are CFA group:C shows HE staining and D shows LFB staining.
2.3.1临床评分和体重实验结果表明,给予选择性滤泡辅助性T细胞阻断剂能够缓解EAE疾病的进程。EAE组实验鼠在免疫后的第10天开始发病,且随时间延长,病情逐渐加重,约至实验的第15天达到高峰,随后缓解。anti-ICOSL组实验鼠第11天开始发病,随后以较轻的病情持续12天后缓解。EAE组实验鼠第13~17天临床评分与给药组实验鼠相比,差异具有统计学意义(P<0.05,P<0.01)。给药组实验鼠在发病期间体重平均维持在约16 g;在第14~17天,EAE组实验鼠体重下降较给药组明显,见图4。
2.3.2实验鼠脊髓脱髓鞘和淋巴细胞浸润情况实验结果表明,EAE组小鼠脊髓组织中可见明显的、多发且散在的炎性细胞浸润和脱髓鞘,anti-ICOSL治疗组小鼠脊髓组织中少见明显的炎性细胞浸润和脱髓鞘,CFA组小鼠脊髓组织中未见有炎性细胞浸润和脊髓脱髓鞘发生。表明滤泡辅助性T细胞选择性抑制剂anti-ICOSL 治疗可以减轻疾病,见图5。

图3 滤泡辅助性T细胞在C57BL/6小鼠淋巴结(A)和脾(B)中的表达Fig.3 Expression of T follicular cells on lymph nodes(A) and spleen(B)Note: *.P<0.001.
2.3.3滤泡辅助性T细胞选择性阻断剂anti-ICOSL对T细胞活化影响实验结果表明,anti-ICOSL组和EAE组相比,CD44高表达的CD3+T细胞数目无差异,说明T细胞活化不受anti-ICOSL的影响;在免疫5 d后,无论是anti-ICOSL组还是EAE组淋巴结CD44高表达的CD3+T细胞数目都显著增加(见图6),说明免疫5 d后淋巴结T细胞参与到免疫反应中。
2.3.4滤泡辅助性T细胞选择性阻断剂anti-ICOSL对ICOS高表达的CD4+CXCR5-T细胞数目影响实验结果显示,EAE组和anti-ICOSL组淋巴结及脾中ICOS高表达的CD4+CXCR5-T细胞数量相近,差异无统计学意义(P>0.05),见图7。说明anti-ICOSL治疗未抑制ICOS分子在T细胞上的表达。
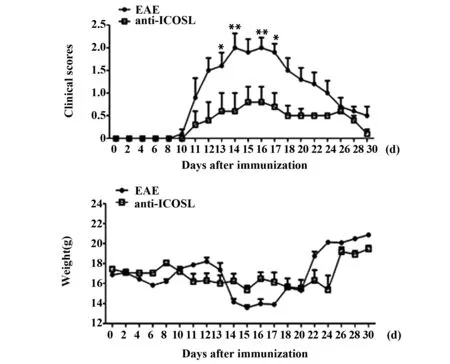
图4 anti-ICOSL 治疗的临床评分及体重变化Fig.4 Clinical scores and weight of therapy by anti-ICOSLNote: *.P<0.05;**.P<0.01.

图5 脊髓组织LFB染色(ACE)及HE(BDF)染色(×200)Fig.5 LFB staining(ACE) and HE staining(BDF) of spinal cord(×200)
2.3.5滤泡辅助性T细胞选择性阻断剂anti-ICOSL 对Tfh细胞数量的影响结果表明:在给予选择性阻断剂anti-ICOSL后,小鼠CD4+ICOS+CXCR5+Tfh细胞水平表达下降;anti-ICOSL组与EAE组相比,各时间点的小鼠淋巴结中Tfh细胞表达水平均下降;至第8天,EAE组小鼠的Tfh细胞所占百分比明显高于anti-ICOSL组(P<0.01),见图8A。anti-ICOSL组小鼠脾中的Tfh细胞百分比较EAE组均有所下降,见图8B。
2.3.6滤泡辅助性T细胞表达减少对生发中心形成的影响结果表明,至第8天,EAE组脾组织中可以看到完整的次级滤泡的形态,略呈圆形,次级滤泡中可见散在的PNA阳性细胞分布。在CFA组中未见次级滤泡形态,在anti-ICOSL组中仅可见完整的次级滤泡形态,未见PNA阳性细胞。至第15天,EAE组小鼠脾组织次级滤泡中可见成簇的PNA阳性细胞聚集,表明此时成熟的生发中心已经形成;CFA组和anti-ICOSL组中仍未见PNA阳性细胞,见图9。
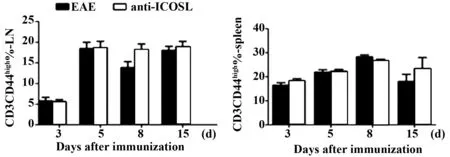
图6 anti-ICOSL对淋巴结和脾中T细胞活化的影响Fig.6 Influence of anti-ICOSL IgG on activation of T cells in lymph nodes and spleen

图7 anti-ICOSL对淋巴结和脾中ICOS高表达CD4+CXCR5- T细胞数目的影响Fig.7 Influence of anti-ICOSL IgG on number of CD4+CXCR5- T cells with high ICOS expression in lymph nodes and spleen
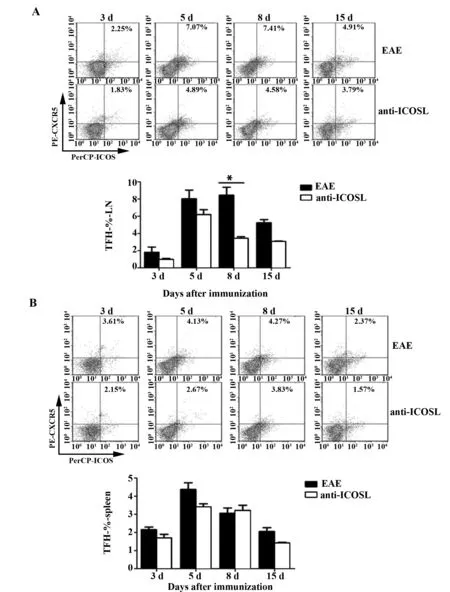
图8 anti-ICOSL对滤泡辅助性T细胞在淋巴结(A)和脾(B)中表达的影响Fig.8 Influence of anti-ICOSL IgG on expression of Tfh cells in lymph nodes(A) and spleen(B)Note: *.P<0.01.

图9 滤泡辅助性T细胞减少对生发中心形成的影响Fig.9 Influence of decreased Tfh cells on germinal centreNote: A,D.EAE;B,E.Anti-ICOSL group;C,F.CFA group.A,B,C.×40;D,E,F.×200;Arrows indicate PNA positive cells.
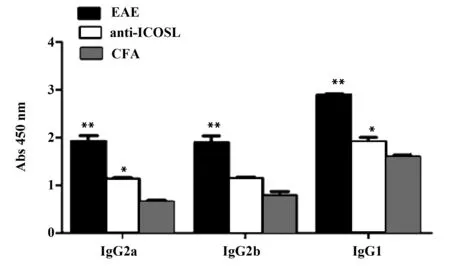
图10 滤泡辅助性T细胞减少对髓鞘蛋白特异性抗体分泌的影响Fig.10 Influence of decreased T follicular helper cells on MOG-specific AbsNote: *.P<0.01;**.P<0.001.
2.3.7滤泡辅助性T细胞减少对髓鞘蛋白特异性抗体分泌的影响结果显示,anti-ICOSL组各抗体分泌水平较EAE组低(IgG2a,P<0.001;IgG2b,P<0.001; IgG1,P<0.001);anti-ICOSL组各抗体分泌水平较CFA组高(IgG2a,P<0.01; IgG1,P<0.01)。随着TFH细胞减少,髓鞘蛋白特异性抗体分泌水平也相应降低,见图10。
3讨论
MS一直被认为是由CD4+T细胞所介导的自身免疫性疾病,在遗传或者感染等因素的作用下,CD4+T细胞被刺激活化成中枢神经系统抗原特异性Th1、Th17细胞,并分泌IFN-γ、TNF-α、IL-12、IL-17、IL-21、IL-22等致炎性细胞因子引起一系列炎症反应引起MS的发生[3]。此外,有实验表明MS与B细胞也有着密切的关系。中枢神经系统包含多种B细胞营养因子,如BAFF(B-cell-activating factor of the TNF family)、APRIL(A proliferation-inducing ligand)以及浆细胞存活因子CXCL12,为B细胞发育提供了良好的微环境[20]。在多发性硬化患者的脑和脑脊液中B细胞增殖并分泌IgG[21],同时B细胞作为专职抗原提呈细胞将抗原提呈给T细胞[22]。
滤泡辅助性T细胞(Tfh),定位于淋巴滤泡,表型为CXCR5+CD40L+ICOS+,主要通过分泌IL-21辅助B细胞产生对T细胞依赖抗原应答[23]。Tfh已被证实了在许多免疫缺陷病和自身免疫性疾病中发挥作用[16,24]。众所周知,自身免疫紊乱主要和高亲和力致病性抗体的产生息息相关。lupus疾病模型中,错误的Tfh细胞的扩增,可以导致自身抗体分泌增加[25]。在Roquin san/san 转基因鼠中发现过多的Tfh细胞表达,使生发中心的反应增强,利用分离出来的Tfh细胞回输给正常的野生型鼠,会自发性形成生发中心。在SLE模型鼠中,阻断ICOSL-ICOS 信号通路可以通过减少Tfh细胞,从而减弱生发中心的反应,从而抑制疾病发生[26]。
Tfh细胞是辅助B细胞发挥体液免疫作用的主要效应性T细胞。这种辅助作用可以分为四个阶段。①携带特异性抗原的树突状细胞将抗原提呈给初始T细胞,使之分化为效应性Tfh细胞;②效应性Tfh细胞迁移到滤泡T-B细胞边界与抗原预激的B细胞相互接触;③经识别共同抗原的B细胞接触后,一部分效应性Tfh细胞迁移进入生发中心,调节抗原特异性记忆B细胞的产生;④另一部分记忆性Tfh细胞将持续存在于初始活化的微环境中调节抗原特异性记忆B细胞对再次抗原免疫的应答[27]。抗原与受体的结合程度,T-B细胞间接触的持久性,识别共同抗原的T-B细胞上信号分子的相互作用等影响因素始终贯穿始终。Tfh细胞的形成与维持和以下信号通路有关:T细胞活化的第二信号CD28和B7(即CD80/CD86),CD4+CD3-淋巴组织刺激细胞表达的OX40L和CD30L提供的刺激信号,T-B细胞接触并相互作用的双重信号刺激CD40/CD40L,ICOS/ICOSL,表面信号淋巴细胞激活分子相关蛋白(SLAM-associated protein,SAP)介导的信号通路[28]。上述作用最终都直接或间接引起Bcl-6的表达,而Bcl-6转录因子,对于T细胞持续表达CXCR5,活化T细胞迁移进入淋巴滤泡以及GC的形成都具有重要作用[29]。
我们在前期工作中利用RT-PCR以及Western blot的方法检测了Tfh细胞主要转录因子Bcl-6在外周淋巴器官脾和淋巴结的表达情况,结果Bcl-6在EAE发病的起始期表达有所增高。我们把检测的时间点集中在免疫后第3、5、8、15天,利用流式细胞术检测Tfh细胞的表面标志,结果显示,Tfh细胞参与了EAE的疾病发生,并且在免疫后的初始期,即第5天时表达明显增高,与CFA组相比有统计学意义,高峰期即免疫后15 d表达略有下降。
由于Tfh细胞在EAE中表达主要集中在发病起始期,我们提出用阻断剂选择性抑制Tfh细胞能否通过抑制Tfh细胞介导的体液免疫,进而缓解EAE的发病症状而起到治疗作用这样一个疑问。Tfh细胞的分化要求与B细胞相互作用得以维持,在这个过程中参与的关键信号通路是ICOS-ICOSL共刺激分子信号[30]。ICOS广泛表达于活化的效应性T细胞上,而ICOSL是B细胞组成性表达的表面分子。因此我们在免疫早期隔日给予anti-ICOSL封闭B细胞上的ICOSL分子,试图通过阻断ICOS-ICOSL的信号通路,抑制Tfh细胞的表达。本实验显示,给予anti-ICOSL选择性封闭性抗体后,该组小鼠的发病减轻,高峰期中枢炎性细胞浸润减少,脱髓鞘损伤减轻。然而,整体T细胞在免疫早期及高峰期的活化程度不受影响,CD44高表达的CD3+T细胞百分比在各组间没有差异。原因是初始T细胞主要被树突状细胞活化,活化后的T细胞与识别共同抗原的活化B细胞间的相互作用中,存在其他信号通路如CD80/86-CD28、CD40-CD40L等信号通路代偿了被阻断的ICOS-ICOSL信号通路的作用。因此,给予anti-ICOSL抗体后并没有影响ICOS在活化T细胞上的表达,具有分泌致炎性细胞因子的ICOS高表达CD4+CXCR5-细胞数量也没有减少。流式结果显示,在发病起始期Tfh细胞在anti-ICOSL组的数量较EAE组下降,这种趋势在淋巴结中更为明显。由于Tfh细胞的错误扩增与体内高水平自身抗体密切相关[25],那么Tfh细胞减少是否意味着自身抗体水平降低呢?我们进一步实验发现在EAE组免疫第8天时脾中生发中心已经开始形成,第15天时可检测到完整的生发中心,同时血清中可检测到高水平的抗体滴度。在anti-ICOSL组免疫第15天时仍未见有明显的生发中心形成,血清中仅可检测到低水平的自身抗体。
本实验结果表明Tfh细胞参与了EAE的发病过程,anti-ICOSL可以通过选择性抑制Tfh细胞的B细胞辅助作用,抑制生发中心的形成和自身抗体的分泌,从而能够有效缓解EAE的临床表现。
参考文献:
[1]Ciccarelli O,Barkhof F,Bodini B,etal.Pathogenesis of multiple sclerosis:insights from molecular and metabolic imaging[J].Lancet Neurol,2014,13(8):807-822.
[2]Hemmer B,Kerschensteiner M,Korn T.Role of the innate and adaptive immune responses in the course of multiple sclerosis[J].Lancet Neurol,2015,14(4):406-419.
[3]Dendrou CA,Fugger L,Friese MA.Immunopathology of multiple sclerosis[J].Nat Rev Immunol,2015,15(9):545-558.
[4]Steinman L.Immunology of relapse and remission in multiple sclerosis[J].Annu Rev Immunol,2014,32:257-281.
[5]Feinstein A,Freeman J,Lo AC.Treatment of progressive multiple sclerosis:what works,what does not,and what is needed[J].Lancet Neurol,2015,14(2):194-207.
[6]Levin MC,Jackson WC.Developing a therapeutic plan for treating MS:evidence for new treatments[J].J Clin Psychiatry,2014,75(12):e34.
[7]Ransohoff RM,Hafler DA,Lucchinetti CF.Multiple sclerosis-a quiet revolution[J].Nat Rev Neurol,2015,11(3):134-142.
[8]Breitfeld D,Ohl L,Kremmer E,etal.Follicular B helper T cells express CXC chemokine receptor 5,localize to B cell follicles,and support immunoglobulin production[J].J Exp Med,2000,192(11):1545-1552.
[9]Crotty S.T follicular helper cell differentiation,function,and roles in disease[J].Immunity,2014,41(4):529-542.
[10]Ise W.Development and function of follicular helper T cells[J].Biosci Biotechnol Biochem,2015,80(1):1-6.
[11]Hutloff A.Regulation of T follicular helper cells by ICOS[J].Oncotarget,2015,6(26):21785-21786.
[12]Habib J,Deng J,Lava N,etal.Blood B cell and regulatory subset content in multiple sclerosis patients[J].J Mult Scler (Foster City),2015,2(2):1000139.
[13]Monson NL,Cravens P,Hussain R,etal.Rituximab therapy reduces organ-specific T cell responses and ameliorates experimental autoimmune encephalomyelitis[J].PLoS One,2011,6(2):e17103.
[14]Gupta IV,Jewell RC.Ofatumumab,the first human anti-CD20 monoclonal antibody for the treatment of B cell hematologic malignancies[J].Ann N Y Acad Sci,2012,1263:43-56.
[15]Hauser SL,Waubant E,Arnold DL,etal.B-cell depletion with rituximab in relapsing-remitting multiple sclerosis[J].N Engl J Med,2008,358(7):676-688.
[16]Xiao N,Eto D,Elly C,etal.The E3 ubiquitin ligase Itch is required for the differentiation of follicular helper T cells[J].Nat Immunol,2014,15(7):657-666.
[17]Tullius SG,Biefer HR,Li S,etal.NAD+ protects against EAE by regulating CD4+T-cell differentiation[J].Nat Commun,2014,5:5101.
[18]Yao Z,Yang S,He W,etal.P311 promotes renal fibrosis via TGFbeta1/Smad signaling[J].Sci Rep,2015,5:17032.
[19]Brickshawana A,Hinson SR,Romero MF,etal.Investigation of the KIR4.1 potassium channel as a putative antigen in patients with multiple sclerosis:a comparative study[J].Lancet Neurol,2014,13(8):795-806.
[20]Krumbholz M,Derfuss T,Hohlfeld R,etal.B cells and antibodies in multiple sclerosis pathogenesis and therapy[J].Nat Rev Neurol,2012,8(11):613-623.
[21]Franciotta D,Salvetti M,Lolli F,etal.B cells and multiple sclerosis[J].Lancet Neurol,2008,7(9):852-858.
[22]Townsend MJ,Monroe JG,Chan AC.B-cell targeted therapies in human autoimmune diseases:an updated perspective[J].Immunol Rev,2010,237(1):264-283.
[23]King C,Tangye SG,Mackay CR.T follicular helper (TFH) cells in normal and dysregulated immune responses[J].Annu Rev Immunol,2008,26:741-766.
[24]Linterman MA,Rigby RJ,Wong RK,etal.Follicular helper T cells are required for systemic autoimmunity[J].J Exp Med,2009,206(3):561-576.
[25]Sweet RA,Lee SK,Vinuesa CG.Developing connections amongst key cytokines and dysregulated ger minal centers in autoimmunity[J].Curr Opin Immunol,2012,24(6):658-664.
[26]Hu YL,Metz DP,Chung J,etal.B7RP-1 blockade ameliorates autoimmunity through regulation of follicular helper T cells[J].J Immunol,2009,182(3):1421-1428.
[27]McHeyzer-Williams LJ,Pelletier N,Mark L,etal.Follicular helper T cells as cognate regulators of B cell immunity[J].Curr Opin Immunol,2009,21(3):266-273.
[28]Nurieva RI,Chung Y.Understanding the development and function of T follicular helper cells[J].Cell Mol Immunol,2010,7(3):190-197.
[29]Hatzi K,Nance JP,Kroenke MA,etal.BCL6 orchestrates Tfh cell differentiation via multiple distinct mechanisms[J].J Exp Med,2015,212(4):539-553.
[30]Choi YS,Kageyama R,Eto D,etal.ICOS receptor instructs T follicular helper cell versus effector cell differentiation via induction of the transcriptional repressor Bcl6[J].Immunity,2011,34(6):932-946.
[收稿2016-02-03修回2016-04-14]
(编辑张晓舟)
Mechanism of T follicular cells in experimental autoimmune encephalomyelitis
PING Chang-Yun,LI Hu-Lun,ZHAN Xiao-Xia,SUN Bo.
Department of Neurobiology,Harbin Medical University,Harbin 150081,China
[Abstract]Objective:To evaluate the mechanism of T follicular helper cells (Tfh) in experimental autoimmune encephalomyelitis (EAE) via in vivo experiments.Methods: C57BL/6 mice were randomly divided into four groups,CFA group,EAE group,anti-ICOSL group and control group.Lymphocytes of different time points isolated from draining lymph nodes and spleen were stained for T follicular helper cells surface marker and T cells activation surface marker and analyzed by FACS.Observed parameters include inflammatory infiltration,demyelination in spinal cord and germinal center in spleen.ELISA was used to measure the level of antigen specific antibodies.Results: Mice in anti-ICOSL treated group developed mild disease was with lower clinical scores when compared with the EAE group.HE staining results turned out with alleviated inflammation and Luxol Fast Blue staining(LFB) showed no demyelization in anti-ICOSL treated mice compared with non-treated EAE models.Flow cytometry results revealed that percentages of T follicular helper cells decreased though the whole activated degree T cells was not influenced in anti-ICOSL treated group.Fewer ger minal center was found in both anti-ICOSL group and CFA group with reduced secretion of MOG-specific Ab.Conclusion: T follicular helper cells supported the development of cognate B cells,promoted the formation of germinal center,facilitate pathogenic MOG-specific Ab secretion,thus enhance EAE.
[Key words]T follicular helper cells(Tfh);Experimental autoimmune encephalomyelitis(EAE);Germinal center(GC)
doi:10.3969/j.issn.1000-484X.2016.06.004
作者简介:平昌昀(1990年-),女,医学硕士,主要参与多发性硬化及其动物模型的研究,E-mail:pingchangyunhmu@163.com。 通讯作者及指导教师:孙博(1979年),女,博士,教授,硕士生导师,主要从事神经免疫学的研究,E-mail:sunbo720@aliyun.com。
中图分类号Q189
文献标志码A
文章编号1000-484X(2016)06-0787-07
①本文受国家自然科学基金(81571168;81430035)资助。
猜你喜欢
Journal of Hainan Medical College(2023年15期)2023-12-06 07:54:42
影像研究与医学应用(2021年3期)2021-11-30 12:54:47
实用临床医学(2018年9期)2018-12-04 05:35:30
西南医科大学学报(2015年1期)2015-08-22 13:01:46
中国医疗美容(2015年1期)2015-07-12 10:06:52
医学研究杂志(2015年6期)2015-07-01 17:41:11
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
现代检验医学杂志(2015年3期)2015-02-06 01:52:36
结核与肺部疾病杂志(2014年1期)2014-07-18 11:09:33
郑州大学学报(理学版)(2014年2期)2014-03-01 04:20:57