TSLP促进肌成纤维细胞向哮喘小鼠气道募集①
2016-06-29 05:40:58陈壮桂张天托李洪涛邹小玲杨海玲
中国免疫学杂志 2016年6期
关键词:哮喘
孟 平 陈壮桂 张天托 李洪涛 邹小玲 杨海玲
(中山大学附属第三医院呼吸内科,广州510630)
·基础免疫学·
TSLP促进肌成纤维细胞向哮喘小鼠气道募集①
孟平陈壮桂②张天托李洪涛邹小玲杨海玲
(中山大学附属第三医院呼吸内科,广州510630)
[摘要]目的:探讨胸腺基质淋巴细胞生成素(Thymic stromal lymphopoietin,TSLP)与慢性哮喘小鼠气道组织中浸润的肌成纤维细胞的关系。方法:将12只BALB/c小鼠随机分为4组:生理盐水组、屋尘螨(House dust mite,HDM)组、anti-TSLP组、同型对照组。激光共聚焦检测气道组织中出现的肌成纤维细胞。ELISA法检测肺泡灌洗液中TSLP、TGF-β1、IL-25和IL-33的表达水平。结果:拮抗TSLP可显著抑制HDM持续暴露导致的气道结构改变,减少肌成纤维细胞向哮喘小鼠气道组织募集。Anti-TSLP组小鼠BALF中TSLP、TGF-β1和IL-33蛋白水平也较同型对照组明显降低(P值均<0.05)。结论:TSLP促进肌成纤维细胞向慢性哮喘小鼠气道募集是其参与气道重塑的主要机制之一。
[关键词]TSLP;肌成纤维细胞;哮喘;气道重塑
气道重塑和能够维持慢性气道炎症的微环境是慢性持续性哮喘形成的两个必要条件。吸入激素等抗炎治疗能够抑制哮喘的状态,但不能改变病人的预后[1,2]。因此,发现抗炎治疗以外的阻断气道重塑的治疗新方法,是目前亟待解决的临床问题。气道重塑病理结构主要包括上皮细胞黏液化生、肌成纤维细胞增多、细胞外基质过度沉积及上皮下纤维化等[3]。其中,肌成纤维细胞是造成细胞外基质沉积和上皮下纤维化的关键结构细胞。气道上皮在启动宿主气道重塑和纤维增殖的炎症反应中,起着关键作用。胸腺基质淋巴细胞生成素(Thymic stromal lymphopoietin,TSLP)在哮喘患者损伤的气道上皮细胞表达是增加的,且其表达水平和疾病的严重程度成正相关[4]。拮抗TSLP可明显减轻气道炎症、抑制甚至逆转哮喘气道结构的病理改变,被认为是气道炎症和气道重塑的总开关[2]。近年来,很多研究发现,TSLP可通过与气道上皮细胞、成纤维细胞、气道平滑肌细胞等多种结构细胞相互作用促进哮喘气道重塑[5-7]。然而,TSLP与肌成纤维细胞在慢性过敏性哮喘中的关系,目前尚未见文献报道。本研究利用慢性哮喘小鼠模型初步探讨TSLP在慢性哮喘气道重塑过程中对肌成纤维细胞的影响。
1材料与方法
1.1实验材料、试剂屋尘螨提取蛋白(House dust mite,HDM)购自购自美国Greer实验室。Rabbit anti-mouse collagen I(Col I)抗体、 Mouse anti-mouse α-smooth muscle Actin (α-SMA) 抗体均购美国abcam公司;Alexa Fluor@594 goat anti-Rabbit、 Alexa Fluor@488 goat anti-mouse二抗均购自美国Invitrogen公司;小鼠TSLP ELISA试剂盒、小鼠TGF-β1 ELISA试剂盒、小鼠IL-25 ELISA试剂盒和小鼠IL-33 ELISA试剂盒均购自中国深圳欣博盛公司。
1.2方法
1.2.1构建慢性哮喘小鼠气道重塑模型SPF级6周龄雌性BALB/c小鼠(购于上海斯莱克实验动物有限公司) 12只,体重(18±2)g。我们按随机数字表法随机均分为4组:生理盐水组,HDM组,同型对照组,anti-TSLP组。过滤式动物饲养柜内分笼饲养,喂食不含致敏原的特殊饲料。HDM气道暴露方法:小鼠用混合麻醉剂腹腔注射麻醉后,垂直悬挂于9号无菌手术缝线,经鼻腔缓慢滴入10 μl HDM悬液(15 μg HDM溶于10 μl NS),生理盐水组小鼠滴NS,每天1次,连续3 d,停止4 d,共持续5周。Anti-TSLP组从第4周起,在滴入HDM 30 min前,TSLP mAb 20 μg溶于100 μl生理盐水腹腔注射。同型对照组腹腔注射相同剂量的IgG同型对照抗体,生理盐水组和HDM组注射相同剂量的抗体稀释液。
1.2.2支气管激发试验第33天进行小鼠气道反应性测定,将小鼠放入体积描箱,随后雾化生理盐水作为空白对照,再进行乙酰甲胆碱激发,依次由低浓度开始,分别为6.25、12.5、25、50、100 mg/ml。每一个浓度雾化后检测3 min,两次检测之间间隔5 min,观察小鼠出现的生物学行为改变。
1.2.3收集支气管肺泡灌洗液将取血后的小鼠平放于小动物手术台,用医用胶布固定四肢,分离小鼠颈部皮肤、肌肉,切开分离出气管,24G聚乙烯导管行小鼠气管插管,自导管注入NS 1 ml,进行小鼠支气管肺泡灌洗,动作轻柔,反复灌洗3次,回收约0.8~0.9 ml灌洗液。2 000 r/min 4℃低温离心机离心10 min,上清液于-80℃冰箱冻存。
1.2.4小鼠肺脏组织染色及病理组织学观察摘除小鼠左肺,投入4%中性甲醛中固定过夜。梯度乙醇脱水,二甲苯透明30 min,石蜡包埋,切成5 μm厚薄片。脱蜡后进行H&E和PAS染色,分别观察肺组织炎性细胞浸润和气道上皮损伤情况,杯状细胞增生情况。
1.2.5ELISA法检测肺泡灌洗液中TSLP、TGF-β1、IL-25、IL-33的浓度(按试剂盒操作说明书)。
1.2.6免疫荧光标本经脱蜡、水化后,用二抗相同宿主的血清(0.3 mol/ml的甘氨酸+1%BSA+3%的山羊血清)封闭30 min。加入稀释的一抗(anti-collgenⅠ或anti-α-SMA),4℃冰箱孵育过夜。PBS洗10 min × 3次。次日,加入稀释过的偶联荧光二抗。室温下,暗盒中避光孵育1 h。PBS洗10 min×5次。Gold Antifade Reagent with DAPI封片过夜。借助于共聚焦激光共聚焦扫描、摄像并分析。
2结果
2.1观察各组小鼠体征改变各组小鼠随吸入乙酰甲胆碱剂量的增加,均会出现典型的哮喘症状,包括毛发竖起、呼吸急促、易激惹、烦躁不安等。但当吸入同等剂量的乙酰甲胆碱剂量时,HDM组小鼠和同型对照组小鼠的反应明显强于生理盐水组和anti-TSLP组,有个别小鼠甚至出现腹肌抽搐、大小便失禁等重度哮喘表现。此外,Anti-TSLP组小鼠开始出现过激反应的乙酰甲胆碱浓度高于同型对照组。
2.2拮抗TSLP可抑制慢性哮喘小鼠气道结构改变HDM持续暴露导致气道结构病理改变,包括支气管壁炎症细胞大量浸润(图1)、杯状细胞增生(图2)、α-SMA阳性的气道平滑肌层增厚(图3) 和上皮下ColⅠ大量沉积(图4)。与同型对照组相比,anti-TSLP组小鼠的支气管壁周围炎症细胞和杯状细胞数量显著减少。此外,阻断TSLP还可防止气道平滑肌层增厚,抑制ColⅠ分泌,减轻上皮下纤维化。
2.3拮抗TSLP可减少肌成纤维细胞在气道壁的浸润HDM组小鼠气道组织中聚集的肌成纤维细胞数(26.33±2.186)mm-2明显高于生理盐水组(7.667±0.881 6)mm-2,两组差异有显著统计学意义(t值为7.920,P<0.01);Anti-TSLP组小鼠气道组织中肌成纤维细胞(10.33±1.202)mm-2明显低于同型对照组(20.67±2.603)mm-2,两组差异有统计学意义(t值为-3.604,P<0.05)。见图5、6。
2.4拮抗TSLP可下调其他上皮源性细胞因子的表达水平HDM组小鼠BALF中TSLP水平[(206.6±19.7)pg/ml]高于生理盐水组[(93.49±5.062)pg/ml],两组差异有显著统计学意义(t值为5.558,P<0.01);Anti-TSLP组小鼠BALF中TSLP水平[(115.6±11.59)pg/ml]低于同型对照组为[(236.5±34.18)pg/ml],两组差异有统计学意义(t值为-3.35,P<0.05)。Anti-TSLP组小鼠BALF中TGF-β1水平[(75.02±9.48)pg/ml]和IL-33水平[(83.21±4.072)pg/ml]低于同型对照组[分别(405.9±33.09)pg/ml和(146.0±8.832)pg/ml],两组差异有统计学意义(t值分别为-9.612和-6.457,均P< 0.05)。Anti-TSLP组小鼠BALF中IL-25[(84.56±7.685)pg/ml]水平和同型对照组[(107.9±8.434)pg/ml],两组差异有统计学意义(t值为-2.044,P> 0.05),但均高于生理盐水组[(44.68±2.267)pg/ml](t值分别为4.727和7.007,均P> 0.05)。见图7。

图1 H&E染色观察气道壁增厚以及支气管壁周围炎症细胞浸润情况(×400,标尺=50 μm)Fig.1 Representative photographs of H&E staining revealed thickness of airway walls and level of inflammatory cell infiltration(×400,bar=50 μm)Note: A.Saline group;B.HDM-exposed group;C.IgG isotype-treated group;D.Anti-TSLP-treated group.

图2 PAS染色观察气道上皮细胞黏液化生情况(×400,标尺=50 μm)Fig.2 Representative photographs of PAS staining revealed hyperplasia of airway goblet cell(×400,bar=50 μm)Note: A.Saline group;B.HDM-exposed group;C.IgG isotype-treated group;D.Anti-TLP-treated group.

图3 荧光染色观察支气管上皮下α-SMA阳性(绿色荧光表示)的气道平滑肌层增厚情况(×400,标尺=50 μm)Fig.3 Representative photographs of α-SMA (α-SMA:green) staining revealed thickness of airway smooth muscle(×400,bar=50 μm)Note: A.Saline group;B.HDM-exposed group;C.IgG isotype-treated group;D.Anti-TLP-treated group.
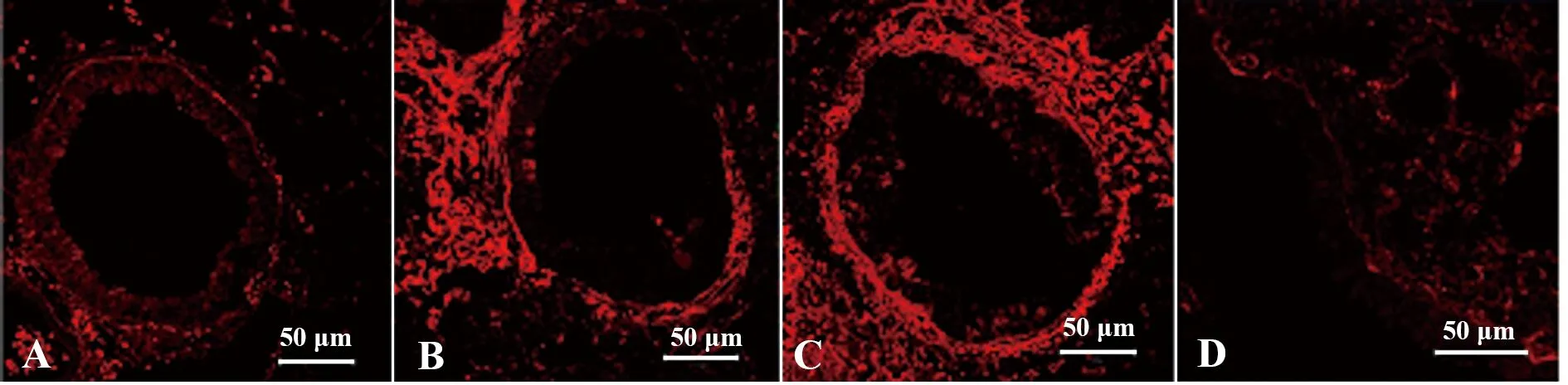
图4 荧光染色观察支气管壁周围Ⅰ型胶原蛋白(红色荧光表示)沉积厚度(×400,标尺=50 μm)Fig.4 Representative photographs of collagen Ⅰ (collagen Ⅰ:red) staining revealed thickness of peribronchial collagen Ⅰ(×400,bar=50 μm)Note: A.Saline group;B.HDM-exposed group;C.IgG isotype-treated group;D.Anti-TSLP-treated group.

图5 激光共聚焦检测气道组织中出现的Col I/α-SMA双阳性的肌成纤维细胞(×400,标尺=50 μm)Fig.5 Fluorescence-labeled Col I/α-SMA-dual-positive fibrocytes in lung were examined by confocal microscopy (×400,Bar=50 μm )Note: α-SMA.Green;Col Ⅰ.Red;DAPI.Blue.

图6 定量分析各组小鼠肺组织中出现的肌成纤维细胞数Fig.6 Quantification of Col Ⅰ+/α-SMA+myofibroblasts in lungNote: Data are expressed as the mean number of myofibroblasts in per mm2 of airway wall (n=3 per group);*.P< 0.01,**.P< 0.001 compared with the saline control group;#.P< 0.05 compared with the IgG isotype control group.

图7 各组小鼠肺泡灌洗液中TSLP、TGF-β1、IL-33和IL-25的蛋白表达水平Fig.7 Expression levels of TSLP,TGF-β1,IL-33 and IL-25 in BALFNote: *.P< 0.05,**.P< 0.01,***.P< 0.001 compared with the saline control group;#.P< 0.05,##.P< 0.001 compared with the IgG isotype control group.
3讨论
肌成纤维细胞是一群具有高度迁移能力的气道平滑肌样细胞,在许多正常和病理组织中存在,既具有平滑肌细胞的收缩特性,又兼有成纤维细胞的基质重塑特性[8,9]。一些研究在慢性阻塞性哮喘患者的气道平滑肌组织中检测到大量肌成纤维细胞,且其数量和基底膜厚度以及哮喘严重程度成正相关[10,11]。此外,细胞外基质过度沉积和上皮下纤维化是哮喘气道重塑发生最早和最关键的部分。而肌成纤维细胞被认为是组织损伤时最具活性的分泌组织细胞之一。因此,肌成纤维细胞是诱导哮喘气道重构的关键间质细胞,在哮喘气道重塑过程中扮演了重要角色。和先前的研究结果一致,我们在长期接受过敏原暴露的小鼠支气管上皮下发现了大量Ⅰ型胶原蛋白沉积Col Ⅰ和α-SMA双阳性的肌成纤维细胞,且其主要定居在胶原沉积的区域。
目前研究发现,哮喘重塑的气道组织中出现的肌成纤维细胞主要有4种来源:间充质干细胞、肺间质成纤维细胞、肺泡上皮细胞以及循环纤维细胞[9,12]。虽然内皮素-1、表皮生成因子等很多细胞因子在肌成纤维细胞增殖和转化中的作用已经被相继报道[10,11],但仍有一些未知影响因素需要进一步的探索和验证。哮喘患者的上皮细胞具有易感性,在外来物质的侵袭下容易发生损伤。越来越多的学者认为上皮-间充质营养单位(Epithelial mesenchymal trophic unit,EMTU)的活化是气道重塑发生发展的中心环节[13]。该学说认为,损伤的上皮细胞可分泌大量上皮源性细胞信号分子,如TSLP、TGF-β1、ET-1、成纤维生长因子等[14,15]。这些信号分子与上皮下的间质细胞相互作用,使开始于修复上皮的重构信号向黏膜下层传播,进而导致气道壁下层炎症和重塑反应的放大。因此,EMTU的活化既维持了哮喘患者慢性炎症微环境,又促进了气道重塑,是慢性持续性哮喘患者的主要发病机制。EMTU的活化在肌成纤维细胞向哮喘重塑的气道组织中募集的过程中起关键的作用。
TGF-β1是上皮细胞和间质细胞之间进行信息交流的经典生长因子。TGF-β1可诱导纤维祖细胞、支气管上皮细胞及肺成纤维细胞等多种细胞转化为α-SMA阳性的肌成纤维细胞[16-18]。然而,很多结构细胞不只对TGF-β1起作用,还同时表达TSLP受体。其配体TSLP是气道上皮损伤后分泌的一种类IL-7样细胞因子,在慢性哮喘小鼠肺组织表达上调,是哮喘气道重塑的重要参与者[2]。已有文献证实,在哮喘气道重塑过程中,TSLP可通过Th2极化之外的途径促进上皮细胞增殖[19]、趋化气道平滑肌细胞迁移[5]。我们在本研究中发现,拮抗TSLP可明显抑制肌成纤维细胞在气道组织的聚集。此外,我们还发现拮抗TSLP可降低BALF中TGF-β1的蛋白浓度。TSLP和TGF-β1在哮喘发病机制中可能不是独立发挥作用的。综上,我们认为TSLP影响肺组织中肌成纤维细胞数量的机制可能包括:(1)TSLP直接诱导肌成纤维细胞增殖或促进其他来源的细胞转分化;(2)通过促进TGF-β1表达,间接促进肌成纤维细胞的迁移和转化。在未来的实验中,我们将通过体外实验对TSLP和TGF-β1在哮喘气道重塑中的相互作用做进一步的探索。
气道上皮是外界和内环境进行交流的主要物理屏障,为了进一步探索损伤的气道上皮与其下的间充质之间的交流信号,我们检测了肺泡灌洗液中TSLP、TGF-β1、IL-25和IL-33的表达水平。数据显示,屋尘螨持续暴露5周可诱导TSLP、TGF-β1、IL-25和IL-33过表达,中和TSLP可降低肺泡灌洗液中TSLP、TGF-β1和IL-33的表达水平。尽管anti-TSLP组和同型对照组BALF中的IL-25表达水平没有统计学差异,但还是可以观察到下降的趋势。IL-33可促进循环纤维细胞和造血祖细胞向哮喘急性加重病人的气道迁移[6,20]。动物研究表明,肺组织IL-33和IL-25的增加可导致气道高反应性和杯状细胞的增生以及IL-13、IL-5、IL-4等的高表达,IL-25还可直接诱导血管生成参与气道重塑过程[21]。Gregory[22]和Yao[23]等通过经鼻滴注IL-25细胞因子诱导了与OVA刺激类似的经典气道重塑表现,拮抗IL-25小鼠气道异常的病理结构改变可被抑制或逆转。尽管IL-25和IL-33也是哮喘气道重塑的重要影响因素,单独抑制任何一个并不能完全阻断气道重塑的发展[24-26],意味着单一细胞因子在哮喘气道重塑中的作用是有限的。我们的数据提示,TSLP的表达水平和IL-25、IL-33、TGF-β1密切相关。由于人支气管上皮细胞本身表达TSLPR和IL-7Rα[27],因此TSLP在体内可以通过自分泌或旁分泌的形式反作用于上皮细胞,进而调节其他细胞因子的表达。先前Gregory等认为拮抗IL-25下调TSLP和 IL-33的表达[24]。因此,在TSLP和其他细胞因子之间存在双向调节机制,相互协调以更好的调控纤维化反应。这些正反馈通路是否真正存在需要进一步的探索。
气道高反应性是哮喘的另一重要特征,表现为气道对各种刺激因子产生过强过早的收缩反应。Gabehart等认为气道炎症和气道重塑都可引发气道高反应性,气道高反应性主要与气道结构改变有关而不依赖于气道炎症[28]。新近有学者提出,气道炎症和气道重塑是平行发生的,Gabehart等的研究进一步解释了,为何常规吸入激素抗炎治疗不能使哮喘患者的临床症状得到有效控制。我们的研究结果表明,拮抗TSLP治疗可降低气道高反应性,改善肺功能。由于TSLP不仅诱导气道炎症,还参与气道重塑,是气道炎症和气道重塑的总开关。所以抑制了TSLP,可以显著降低气道高反应性。
综上,本研究首次证实拮抗TSLP可抑制肌成纤维细胞向慢性哮喘小鼠气道组织募集。因此,我们今后对哮喘的研究不能仅局限于控制Th2型免疫炎症反应,保护气道上皮免受损伤和针对肌成纤维细胞的迁移机制进行靶向干预有望成为未来根治哮喘的新思路。
参考文献:
[1]Grainge CL,Lau LCK,Ward JA,etal.Effect of bronchoconstriction on airway remodeling in asthma[J].N Engl J Med,2011,364(21):2006-2015.
[2]Chen ZG,Zhang TT,Li HT,etal.Neutralization of TSLP inhibits airway remodeling in a murine model of allergic asthma induced by chronic exposure to house dust mite[J].PLoS One,2013,8(1):e51268.
[3]Berair R,Saunders R,Brightling CE.Origins of increased airway smooth muscle mass in asthma[J].BMC Med,2013,11:145.
[4]Cianferoni A,Spergel J.The importance of TSLP in allergic disease and its role as a potential therapeutic target[J].Expert Rev Clin Immunol,2014,10(11):1463-1474.
[5]Redhu NS,Shan L,Movassagh H,etal.Thymic stromal lymphopoietin induces migration in human airway smooth muscle cells[J].Sci Rep,2013,3:2301.
[6]Smith SG,Gugilla A,Mukherjee M,etal.Thymic stromal lymphopoietin and IL-33 modulate migration of hematopoietic progenitor cells in patients with allergic asthma[J].J Allergy Clin Immunol,2015,135(6):1594-1602.
[7]Semlali A,Jacques E,Koussih L,etal.Thymic stromal lymphopoietin-induced human asthmatic airway epithelial cell proliferation through an IL-13-dependent pathway[J].J Allergy Clin Immunol,2010,125(4):844-850.
[8]Hinz B,Phan SH,Thannickal VJ,etal.Recent developments in myofibroblast biology[J].Am J Pathol,2012,180(4):1340-1355.
[9]Phan SH.Genesis of the myofibroblast in lung injury and fibrosis[J].Proc Am Thorac Soci,2012,9(3):148-152.
[10]Wang CH,Huang CD,Lin HC,etal.Increased activation of fibrocytes in patients with chronic obstructive asthma through an epidermal growth factor receptor-dependent pathway[J].J Allergy Clin Immunol,2012,129(5):1367-1376.
[11]Weng C,Chen B,Wang C,etal.The endothelin A receptor mediates fibrocyte differentiation in chronic obstructive asthma.The involvement of connective tissue growth factor[J].Am J Respir Crit Care Med,2013,188(3):298-308.
[12]Lo C,Michaeloudes C,Bhavsar PK,etal.Increased phenotypic differentiation and reduced corticosteroid sensitivity of fibrocytes in severe asthma[J].J Allergy Clin Immunol,2015,135(5):1186-1195.
[13]Hackett TL,Warner SM,Stefanowicz D,etal.Induction of epithelial-mesenchymal transition in primary airway epithelial cells from patients with asthma by transforming growth factor-beta1[J].Am J Respir Cri Care Med,2009,180(2):122-133.
[14]Divekar R,Kita H.Recent advances in epithelium-derived cytokines (IL-33,IL-25,and thymic stromal lymphopoietin) and allergic inflammation[J].Curr Opin Allergy Clin Immunol,2015,15(1):98-103.
[15]Royce SG,Li X,Tortorella S,etal.Mechanistic insights into the contribution of epithelial damage to airway remodeling.Novel therapeutic targets for asthma[J].Am J Respir Cell Mol Biol,2014,50(1):180-192.
[16]Itoigawa Y,Harada N,Harada S,etal.TWEAK enhances TGF-β-induced epithelial-mesenchymal transition in human bronchial epithelial cells[J].Respiratory Res,2015,16(1):48.
[17]Yang ZC,Yi MJ,Ran N,etal.Transforming growth factor-beta1 induces bronchial epithelial cells to mesenchymal transition by activating the Snail pathway and promotes airway remodeling in asthma[J].Mol Med Rep,2013,8(6):1663-1668.
[18]Wojcik KA,Skoda M,Koczurkiewicz P,etal.Apigenin inhibits TGF-beta1 induced fibroblast-to-myofibroblast transition in human lung fibroblast populations[J].Pharmacol Rep,2013,65(1):164-172.
[19]Semlali A,Jacques E,Koussih L,etal.Thymic stromal lymphopoietin-induced human asthmatic airway epithelial cell proliferation through an IL-13-dependent pathway[J].J Allergy Clin Immunol,2010,125(4):844-850.
[20]Bianchetti L,Marini MA,Isgrò M,etal.IL-33 promotes the migration and proliferation of circulating fibrocytes from patients with allergen-exacerbated asthma[J].Biochem Biophys Res Commun,2012,426(1):116-121.
[21]Morita H,Arae K,Unno H,etal.IL-25 and IL-33 contribute to development of eosinophilic airway inflammation in epicutaneously antigen-sensitized mice[J].PLoS One,2015,10(7):e134226.
[22]Gregory LG,Jones CP,Walker SA,etal.IL-25 drives remodelling in allergic airways disease induced by house dust mite[J].Thorax,2013,68(1):82-90.
[23]Yao X,Wang W,Li Y,etal.Characteristics of IL-25 and allergen-induced airway fibrosis in a murine model of asthma[J].Respirology,2015,20(5):730-738.
[24]Gregory LG,Jones CP,Walker SA,etal.IL-25 drives remodelling in allergic airways disease induced by house dust mite[J].Thorax,2013,68(1):82-90.
[25]Bianchetti L,Marini MA,Isgrò M,etal.IL-33 promotes the migration and proliferation of circulating fibrocytes from patients with allergen-exacerbated asthma[J].Biochem Biophys Res Commun,2012,426(1):116-121.
[26]Mizutani N,Nabe T,Yoshino S.Interleukin-33 and alveolar macr-ophages contribute to the mechanisms underlying the exacerbation of IgE-mediated airway inflammation and remodelling in mice[J].Immunology,2013,139(2):205-218.
[27]Miazgowicz MM,Elliott MS,Debley JS,etal.Respiratory syncytial virus induces functional thymic stromal lymphopoietin receptor in airway epithelial cells[J].J Inflamm Res,2013,6:53-61.
[28]Gabehart KE,Royce SG,Maselli DJ,etal.Airway hyperresponsi-veness is associated with airway remodeling but not inflammation in aging Cav1-/-mice[J].Respiratory Res,2013,14(1):110.
[收稿2016-03-21修回2016-06-04]
(编辑张晓舟)
TSLP promotes recruitment of myofibroblasts into airway in asthmatic mice
MENG Ping,CHEN Zhuang-Gui,ZHANG Tian-Tuo,LI Hong-Tao,ZOU Xiao-Ling,YANG Hai-Ling.
Department of Pulmonary Diseases,the Third Affiliated Hospital of Sun Yat-Sen University,Guangzhou 510630,China
[Abstract]Objective:To investigate whether thymic stromal lymphopoietin (TSLP) participate in asthmatic airway remodeling partially by promoting myofibroblast accumulating in the lung.Methods: Twelve mice evenly were randomly divided into four groups:a saline group;an HDM-exposed group;an IgG isotype-treated group and an anti-TSLP-treated group.The supernatant of bronchoalveolar lavage fluid (BALF) was used to analyze the levels of TSLP,IL-25 and IL-33 by ELISA.Fluorescence-labeled collagen Ⅰ(Col Ⅰ)/α-smooth muscle actin (α-SMA) -dual-positive myofibroblasts were examined by confocal microscopy.Results: Chronic allergen exposure induced obviously abnormal airway structural changes,which were inhibited by blocking TSLP.We detected a highly increased number of myofibroblasts in the sub-epithelial zone in mice from HDM-challenged group.However,TSLP neutralization significantly reduced myofibroblasts recruitment.Moreover,blocking TSLP not only decreased the level of TSLP,but also inhibited the expression levels of TGF-β1 and IL-33 in BAL fluid.Conclusion: The results suggest that orchestrating myofibroblasts recruiting into the lungs is one of the main pathogenesis that TSLP involves in airway remodeling in asthmatic mice.
[Key words]TSLP;Myofibroblast;Asthma;Airway remodeling
doi:10.3969/j.issn.1000-484X.2016.06.002
作者简介:孟平(1990年-),女,在读硕士,主要从事哮喘气道重塑的发病机制的研究,E-mail:814985146@qq.com。 通讯作者及指导教师:张天托(1962年-),男,教授,博士生导师,主要从事哮喘及肺部细菌感染方面的研究,E-mail:zhtituli@163.com。
中图分类号R562.2+5
文献标志码A
文章编号1000-484X(2016)06-0777-06
①本文受国家自然科学基金(81470219、81370114)和广东省自然科学基金(S2013010016330)资助。
②共同第一作者。
猜你喜欢
保健医苑(2022年1期)2022-08-30 08:40:04
中老年保健(2021年5期)2021-08-24 07:06:38
中老年保健(2021年6期)2021-08-24 06:53:48
家庭医学(下半月)(2020年4期)2020-05-30 12:42:42
家庭医学(下半月)(2019年11期)2020-01-16 08:39:10
家庭医学(下半月)(2019年10期)2019-11-16 08:59:52
启蒙(3-7岁)(2018年10期)2018-10-13 02:19:32
中成药(2017年6期)2017-06-13 07:30:35
中国中医药现代远程教育(2014年22期)2014-03-01 04:32:52
中国中医药现代远程教育(2014年17期)2014-03-01 04:29:17