
龄级和密度变化对黔东南杉木人工纯林结构及有机碳密度的影响
2016-06-24 07:03:23闵启利张喜张佐玉田炼红杨汉远石扬文杨永彰
中国林副特产 2016年3期
闵启利,张喜,张佐玉,田炼红,杨汉远,石扬文,杨永彰
(1.贵州省锦屏县林业局,贵州锦屏556700;2.贵州省林业科学研究院,贵州贵阳550005;3.贵州省黔东南州林业科学研究所,贵州凯里556000;4.贵州省黎平县林业局,贵州黎平557300;5.贵州省榕江县林业局,贵州榕江557200)
龄级和密度变化对黔东南杉木人工纯林结构及有机碳密度的影响
闵启利1,张喜2*,张佐玉2,田炼红2,杨汉远3,石扬文4,杨永彰5
(1.贵州省锦屏县林业局,贵州锦屏556700;2.贵州省林业科学研究院,贵州贵阳550005;3.贵州省黔东南州林业科学研究所,贵州凯里556000;4.贵州省黎平县林业局,贵州黎平557300;5.贵州省榕江县林业局,贵州榕江557200)
摘要:采用空间替代时间、野外样地调查和室内分析相结合的方法,研究了黔东南杉木人工纯林结构及有机碳密度随龄级和密度变化的规律。结果表明,随杉木林龄增大和密度降低:①林分平均胸径和高度值升高、密度值降低,龄级及密度级内的相应指标值差异显著。②除Pielou指数值外,草本层植物多样性指数增加。灌木层植物多样性指数值随林龄增大而增加、随密度降低而减小。不同龄级及密度级灌木层植物Pielou指数值差异显著。草本层植物Gleason指数值在龄级间差异不显著、密度级间差异显著,Pielou指数值在龄级间差异显著、密度级间差异不显著。③土壤密度值增大、土壤有机碳含量等其它理化指标降低。土壤密度值在龄级间差异显著、密度级间差异不显著,土壤有机碳含量等其它理化指标值在龄级及密度级间差异不显著。④乔木、灌木、凋落物和土壤-植被层有机碳密度值增加,草本和土壤层有机碳密度值在龄级间降低、密度级间升高。乔木层植物有机碳密度值在龄级间差异显著、密度级间差异不显著。⑤林龄和密度对乔木层植物有机碳密度值的贡献率分别为99.06%、0.94%。土壤有机碳密度受土壤层部分指标影响,植被有机碳密度受植被-土壤部分指标综合影响。⑥培育大径级林和增加土壤有机碳含量是提高杉木人工林有机碳密度的有效途径。
关键词:杉木人工林;龄级;密度;林分结构;有机碳密度
杉木(Cunninghamialanceolata)速生丰产、优质,是欧亚大陆东南部湿润亚热带地区的重要用材树种之一[1]。杉木栽培历史悠久、研究领域广泛,其中有机碳密度的相关研究主要集中在不同自然地带林分生物生产力对比[1-2],不同生长阶段林分碳储量[3-8]变化、土壤理化性质[9]和微生物[10]变化以及对林下植被[11]的影响,不同密度林分生物量及土壤肥力变化[12]和林内植被演替[13],林下植物组成和生物量变化[14]以及对土壤肥力的影响[15],林内凋落物动态[16-17]以及对有机碳含量[19]等土壤生态功能指标影响[19]的诸多方面。除小流域定位观测[7-8,16]和地带性对比研究[1-2]外、杉木林的大部分研究以经营场[4-6,12-15,17-19](站[3,9-10,20])和县(市)[11,21-22]为对象,缺少由若干县(市)组成的生态区(小区)尺度的杉木林研究、也缺少龄级和密度等级变化对杉木林结构及有机碳密度的影响研究。
贵州省黔东南地区历史上即为杉木中心产区、素有苗杉之称,杉木人工林生物量及分配[21]和有机碳储量[22]研究等已有少量报道。本文以黔东南州锦屏-黎平-榕江3县的杉木人工林为对象,研究龄级和密度变化对杉木林结构及有机碳密度的影响规律,旨在为杉木人工林经营及碳汇管理提供科学依据。
1研究区概况及研究方法
1.1研究区概况
研究在国家林业局黎平生态站进行。调查区域108°04′~109°31′E,25°26′~ 26°47′ N。属雪峰台凸,出露地层主要为前震旦纪下江群的板岩、变质砂岩和砂页岩,所发育土壤粘重深厚、保水保肥性好;其次为二叠纪石灰岩,分布于从江县贯洞和皮林至黎平县中潮和德凤到锦屏县敦寨一带,或为第四纪粘土覆盖、或出露发育为石灰土。研究区属中亚热带季风湿润气候区,年均温15.4~18.5℃,1月均温5.9℃、7月均温26.3℃,≥10℃积温5192℃174~209 天;年降雨量1300 mm左右,分布均匀、雨日相近,相对湿度80%~85%,日照时数1212h/a、太阳辐射量3.3496×105~3.7683×105J/cm2·a。植被属地带性中亚热带常绿阔叶林和岩溶性常绿落叶阔叶混交林。杉木林下灌木植物主要有白栎(Quercusfabri)、茅栗(Castaneasequinii)、柃木(EuryaSP.)、油茶(Camelliaolifera)、山胡椒(Linderaglauca)、杜茎山(Maesajaponica)、糯米团(Memorialishirta)、鼠刺(Iteachinensis)、水竹(Phyllostachyscongesta)和方竹(ChimonobambusaSP.)等,草本植物主要有山姜(Alpiniajaponica)、麦冬(Liriopespicata)、瘤足蕨(Plagiogyriajaponica)、白茅(Imperatacylindericavar.major)、金茅(Eulaliaspeciosa)、野古草(Arundinellahirta)、鸭咀草(Ischaemumaristatum)、五节芒(Miscanthusfloridulus)、芒萁(Dicranopterisdichotoma)、狗脊(Woodwardiajaponica)、渐尖毛蕨(Cyclosorusacuminatus)、金星蕨(Parathelypterisglanduligera)、蕨(Pteridiumaquilinum)和卷柏(SelaginellaSP.)等。
1.2研究方法
1.2.1野外样地调查
依据榕江、黎平和锦屏县第3次(2004~2006 年)森林资源二类调查小班数据库,在相似生境及经营水平的杉木纯林林班(小班)中,分别龄级、密度选择调查林分,经实地查验后进行样地调查。样地总量23 个,其中榕江、黎平和锦屏县的样地数量分别为7、8和8 个,均为地带性黄壤,海拔高度变幅340~669 m、坡度0~45°、土壤A层厚度为2~28 cm、B层为8~60 cm、A+B层为15~77 cm。按不同龄级和密度等级组合为6 个类型(表1)。

表1 杉木人工纯林不同年龄和密度等级调查样地数量分布
调查样地面积20m×20m,样地内部按灌木、草本及凋落物发育较好、一般和较差3个等级分别设置3个5m×5m的灌木层、3个1m×1m的草本层植物和凋落物调查样方。调查内容在乔木层为种名、胸径、高度及郁闭度,灌木层为种名、株数、平均地径、高度、冠幅及盖度,草本层为种名、株数、平均高度及盖度。杉木生物量分别构件按相关模型推算,灌木层(2m×2m)、草本层(1m×1m)和凋落物层(1m×1m)植物生物量按收获法测定。记录样地的海拔高度,坡度、坡位、坡向,母岩类型,土壤名称。每块样地中部挖掘土壤剖面1 个,记录自然发生层剖面特征、各层提取土壤环刀1 牧,环刀密封后带回室内分析土壤物理主要指标值;样地内按S型布点5 个,自然发生层取样、等量混合,样品重量1 kg左右,带回室内风干后测定土壤化学主要指标值。
1.2.2室内资料分析
(1)植物多样性指数计算[23]
其中:S、A、Ni和N分别为样方物种数、面积、某物种株数和总株数。
(2)土壤理化指标值测定[24]
土壤密度、孔隙度、田间持水量和最大持水量采用环刀法,pH值采用电位法,有机碳含量采用重铬酸钾氧化-外加热法,全氮含量采用半微量凯氏扩散法,水解氮含量采用碱解-扩散法,全磷含量采用碱溶-钼锑抗比色法,有效磷含量采用盐酸-硫酸浸提法,全钾含量采用碱溶-火焰光度法,速效钾含量采用乙酸铵浸提-火焰光度法,阳离子交换量采用乙酸铵交换法。
(3)杉木单株生物量模型筛选
国内杉木生物量模型众多[1-2,4,7-8,20-21],本文模型选择依据:①以研究区为中心选择模型,近者优先;②树杆(皮)、树枝、树叶和根系生物量模型相对完整组优先,生物量随胸径、树高变化规律符合研究区杉木生长的一般规律;③承认单一树种生物量变化在不同地区和龄级区段间模拟的同一性,以忽略气候、立地要素及经营措施等对生物量模型的影响。通过胸径(DBH)、树高(H)和构件生物量(W)散点图对比,以及和已测杉木林样地生物量[7,20-22]的关联度,认定冯宗炜等[2]在湖南会同所建杉木单株生物量模型为最优模型(表2)。

表2 杉木单株构件生物量模型
(4)有机碳密度计算
杉木林有机碳密度(C,t/hm2)由植被层(Cp)和土壤层(Cs) 2 个部分组成。
C=Cp+Cs
其中:i为植被分层,包括乔木层、灌木层、草本层和凋落物层4层;j为构件数量,乔木层分杆(皮)、树枝、树叶和根系4部分,灌木和草本层植物生物量包括地上和地下2部分,枯落物生物量仅地上部分;k为土层,林地土壤中第四纪粘土C层厚度变幅较大、其它母岩C层厚度变幅较小,本文仅进行A层和B 层土壤分析,土层厚度(Dk)分别取2层平均值(DA=15cm、DB=25cm)以利于比较不同调查样地间土壤有机碳密度值差异;Wij为构件生物量(t/hm2),Cpij、Csk分别为植物构件和土层有机碳含量率(g/kg),其中Cpij参照文献[22]、Csk为土壤实测值;Vk为土壤密度(g/cm3),α、β为单位转换系数。
(5)数据分析
数据处理使用Excel2003,统计分析参见相关文献[25]。其中方差分析采用不等次重复单因素方差分析,双均数比较采用T检验法、多均数比较采用S检验法,P<0.05时差异显著、P<0.01时差异极显著。直线相关分析使用Pearson相关系数,P<0.05时相关显著、P<0.01时相关极显著。
2结果分析
现有研究发现杉木林生长受林分结构和立地要素的综合影响[1-4,6,21-22]。23 个调查样地分布范围较大,水和热量因素变化较小,同属清水江-都柳江下游低山丘陵区。杉木林乔木层植物平均胸径、高度、密度及有机碳密度,以及土壤A层及B层有机碳含量和海拔高度、坡度、土壤A层、B层及A+B层厚度的相关性不显著。表明研究区调查样地杉木林乔木层植物有机碳密度及土壤有机碳含量变化受林分结构主要指标的影响。
2.1林分结构主要指标值变化
方差分析发现不同龄级和密度级杉木人工纯林中,乔木层植物郁闭度值差异不显著,平均胸径(P<0.01)、高度(P<0.01)及密度值(P<0.01)差异显著。灌木层植物盖度、平均地径、高度及密度值差异不显著。草本层植物盖度和高度值差异不显著,密度在龄级中差异不显著、在密度级中差异显著(P<0.05)(表3)。层片植物密度呈草本层>灌木层>乔木层、高度呈乔木层>灌木层>草本层、粗度呈乔木层>灌木层的趋势。
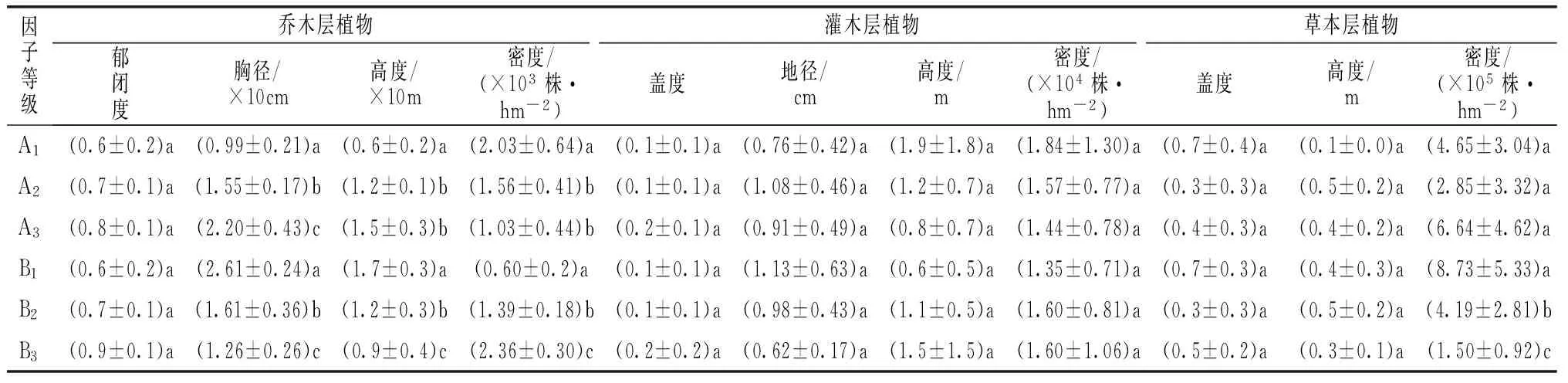
表3 不同龄级和密度级杉木人工林结构主要指标值变化
注:A和B的标注同表1。表中数值为“平均数±标准差”格式,数值后小写字母表示年龄或密度因子内相同指标不同等级值间的S检验结论,相异差异显著、相同差异不显著。
随林龄增大,乔木层植物郁闭度及灌木层盖度值增大、草本层植物盖度值减小,乔木层植物平均胸径和高度值增大、密度值降低,其中杉木幼龄林平均胸径、高度及密度值和中龄林及成熟林的相应值差异显著(P<0.05),平均胸径值在幼龄林、中龄林和成熟林间差异显著(P<0.05)。灌木层植物平均地径值升高、高度及密度值除低,草本层植物高度和密度值增大。随密度增大,乔木层植物郁闭度及灌木层植物盖度值增大、草本层植物盖度值减小,乔木层植物平均胸径和高度值降低、密度值增大,平均胸径、高度及密度值在幼龄林、中龄林和成熟林间差异显著(P<0.05)。灌木层植物平均地径值降低、高度及密度值升高,草本层植物高度和密度值减小、密度值在密度级间差异显著(P<0.05)。符合杉木林生长及乔木-灌木-草本层植物协同演化的一般规律[1-2,8,11-15]。
2.2植物多样性主要指标值变化
方差分析发现不同龄级和密度级杉木人工林中,灌木层植物Gleason指数、Simpson指数和Shannon-Wiener指数值差异不显著,Pielou指数值差异显著(P<0.05);草本层植物Simpson指数和Shannon-Wiener指数值差异不显著,Gleason指数值在龄级间差异不显著、在密度级间差异显著(P<0.05),Pielou指数值在龄级间差异显著(P<0.05)、在密度级间差异不显著(表4)。不同龄级和密度级杉木林的Gleason指数呈草本层>灌木层、Simpson指数呈灌木层>草本层的趋势,Shannon-Wiener指数除幼龄林和高密度林外、呈灌木层>草本层的趋势,Pielou指数除成龄林和低密度林及中密度林外、呈灌木层<草本层的趋势。
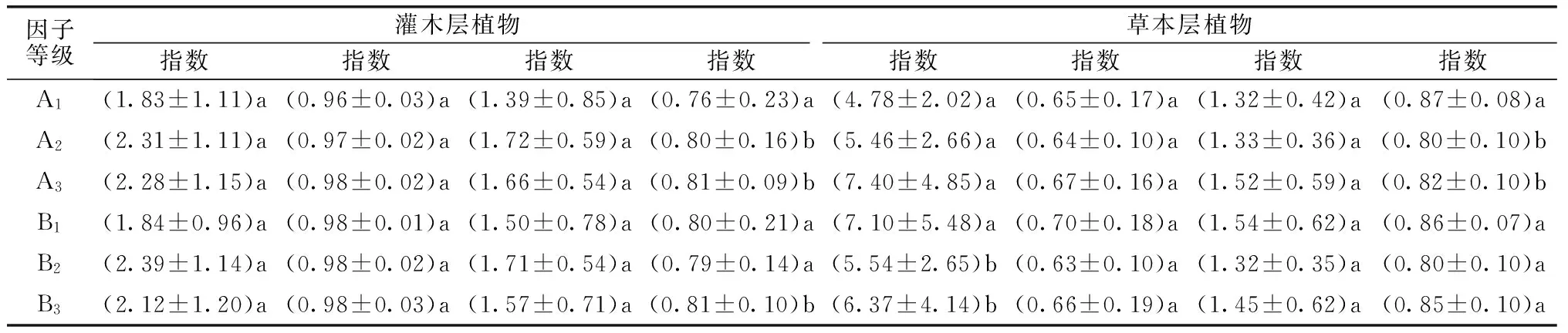
表4 不同龄级和密度级杉木人工林植物多样性主要指数值变化
注:表内标注同表3。
随林龄增大,Pielou指数值在灌木层增大、草本层降低,灌木层及草本层植物Pielou指数值在幼龄林和中龄林及成熟林间差异显著(P<0.05),其它灌木层与草本层植物多样性指数增加。随密度增大,灌木层植物Gleason指数、Simpson指数、Shannon-Wiener指数和Pielou指数值升高,草本层植物相应多样性指数值降低,其中灌木层植物Pielou指数值在高密度林和中密度林及低密度林间差异显著(P<0.05)、草本层植物Gleason指数值在低密度林和中密度林及高密度林间差异显著(P<0.05)。杉木人工林内植物多样性指数的变化同林分结构演化具有相似性规律。
2.3土壤理化主要指标值变化
方差分析发现杉木人工林不同龄级土壤A层密度值差异显著(P<0.05),B层土壤密度、最大持水量、毛管持水量及田间持水量值差异显著(P<0.01),其它理化指标值差异不显著;不同密度级土壤理化主要指标值差异不显著(表5~6)。不同龄级及密度级土壤物理主要指标值的层间差异不显著,全氮含量、水解氮含量及有效磷含量在中密度林、中龄林及成龄林土壤层间差异显著,水解氮含量在低密度和高密度林土壤层间差异显著,全氮含量在低密度林土壤层间差异显著。土壤理化指标呈A层

表5 不同龄级和密度级杉木人工林土壤物理主要指标值变化
注:大写字母表示同一龄级或密度等级类型相同指标在不同土层(A、B)间的T-检验结论,相异差异显著、相同差异不显著。表内其它标注同表3。
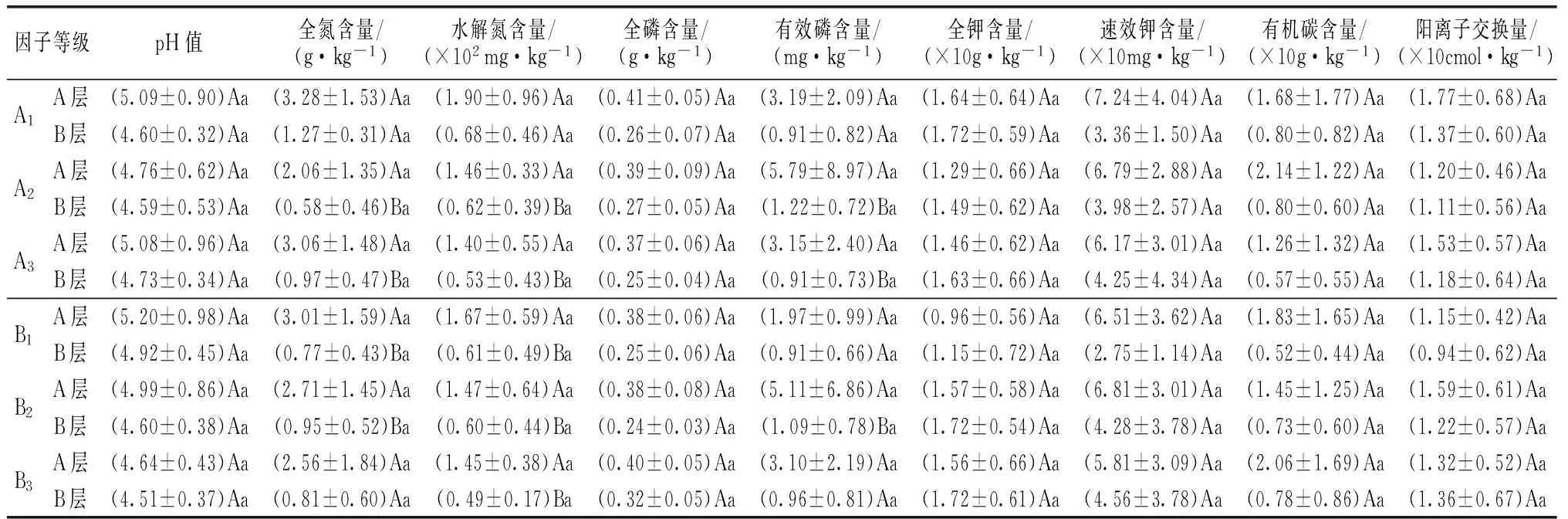
表6 不同龄级和密度级杉木人工林土壤主要化学指标值变化
注:表内标注同表5。
随林龄增大,除土壤A层与B层密度值增大外,B层土壤田间持水量、毛管孔隙度、pH和速效钾含量值也有增加趋势,土壤有机碳含量等其它理化指标值均呈降低趋势,土壤A层及B层呈降低趋势的指标数量为93.75%、68.75%。随密度增大,除土壤A层与B层密度及pH值降低外,A层土壤速效钾含量和阳离子交换量、B层土壤水解氮含量和有效磷含量也呈降低趋势,土壤有机碳含量等其它理化指标值均呈增加趋势, 土壤A层与B层呈增加趋势的指标数量分别为75.00%。杉木人工林土壤质量有随年龄增大、密度减小而呈降低的趋势。
2.4有机碳密度值变化
方差分析发现不同龄级杉木林乔木层植物有机碳密度值差异显著(P<0.01)、灌木层、草本层、凋落物层、土壤层及土壤-植被层有机碳密度值差异不显著,不同密度级杉木林植被各层片、土壤层及土壤-植被层有机碳密度值差异不显著(表8)。不同龄级及密度级杉木林有机碳密度值呈乔木层>凋落物层>灌木层>草本层的趋势,除幼龄林外、中龄林和成熟林有机碳密度值呈乔木层>土壤层的趋势,除高密度林外、低密度林和中密度林有机碳密度值呈乔木层>土壤层的趋势。
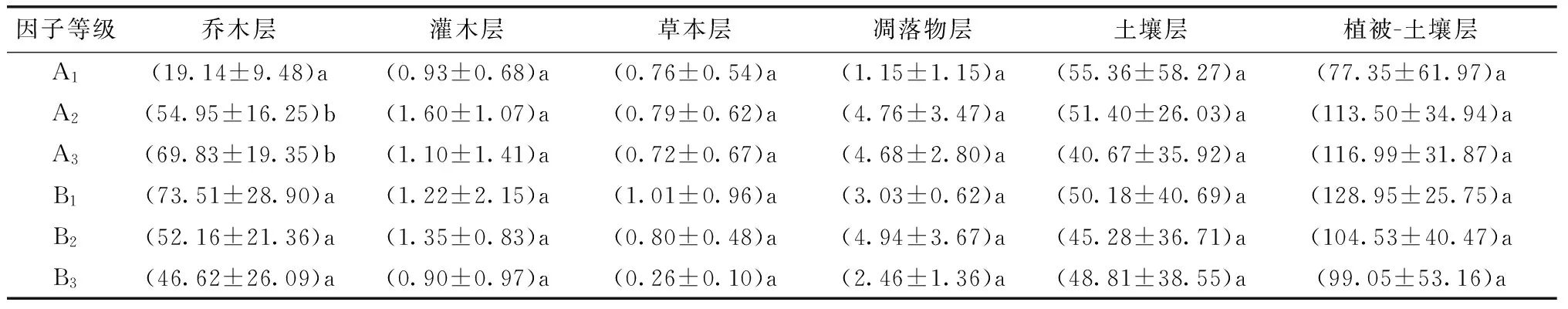
表7 不同龄级和密度级杉木人工林有机碳密度组成变化 t·hm-2
注:表内标注同表3。
随林龄增大,植被层中乔木层、灌木层、凋落物层和土壤-植被层有机碳密度值增加、草本层和土壤层有机碳密度值降低,其中乔木层植物有机碳密度值在幼龄林和中龄林及成龄林间差异显著(P<0.05)。随密度增大,植被层中乔木层、灌木层、草本层、凋落物层、土壤层和土壤-植被层有机碳密度值减小。杉木人工林有机碳密度值随林龄增大和密度减小,乔木层、灌木层、凋落物层和土壤-植被层有机碳密度值呈增大趋势,草本层和土壤层有机碳密度值有随林龄增大而减小、随密度减小而增大的趋势。
2.5有机碳密度的影响因素
杉木林有机碳密度由植被层和土壤层构成,其中乔木层、灌木层、草本层、凋落物层和土壤层(0~40 cm)有机碳密度值的组成比例为51.28%、1.14%、0.69%、3.76%和43.13%,乔木植物和土壤是杉木林有机碳密度值的构成主体部分,凋落物、灌木及草本植物有机碳密度值仅占5.59%、但是杉木人工林不可或缺的组成单元,有重要的生态价值。
相关分析发现林龄和乔木层植物有机碳密度值(P<0.01)相关显著,和灌木层、草本层、凋落物层及土壤层有机碳密度值相关不显著;林分密度和草本层植物有机碳密度值(P<0.05)相关显著,和乔木层、灌木层、凋落物层及土壤层有机碳密度值相关不显著。进一步分析发现乔木层植物有机碳密度值和林龄及密度的直线相关式(P<0.01)显著、林龄和密度对乔木层植物有机碳密度的贡献率为99.06%、0.94%,土壤层及土壤-植被层有机碳密度的相关式未达显著水平。分析还发现灌木层植物有机碳密度和土壤A层水解氮含量(P<0.05)及土壤B层非毛管孔隙度值(P<0.05)相关显著。草本层植物有机碳密度和乔木层植物平均胸径(P<0.05)及高度(P<0.05)、土壤A层pH值(P<0.05)相关显著。凋落物层有机碳密度和土壤A层有效磷含量值(P<0.05)相关显著。土壤有机碳密度和土壤A层及B层全钾含量(P<0.01)、速效钾含量(P<0.05)、有机碳含量(P<0.01)、阳离子交换量(P<0.05)、土壤A层全氮含量(P<0.01)及B层有效磷含量值(P<0.05)相关显著。表明土壤有机碳密度受土壤层部分指标的影响,植被有机碳密度受植被-土壤层部分指标的综合影响。
3讨论与建议
3.1林龄和密度级对杉木人工林有机碳密度值影响的区域性变化
研究发现杉木人工林乔木层植物[4,6-8]、凋落物[3-4,6-8,17]及植被层[4,6-7]有机碳密度值随林龄增大而增加,同已有文献报道相一致。幼龄、中龄及成龄林乔木层植物有机碳密度值高于河南信阳杉木林[6](111.09%、132.17%、112.94%,以本文为基数的相对值,下同),部分龄级低于福建南平杉木林[4](89.61%、105.58%、86.69%)、部分龄级高于贵州开阳杉木林[7](69.96%、115.46%、121.69%),中龄期[2](生物量碳转换系数为0.5)是广西玉林、湖南会同和河南信阳杉木林的109.65%、92.98%、139.82%,幼龄期[2]是贵州麻江杉木林的125.68%。不同龄级乔木层植物有机碳密度值存在区域性差异,是杉木林对水和热量因素配合的长期适应性结果[1]。林下植物有机碳密度变化相似于部分文献[4,6,8]、相异于另一些文献[3,7,9,17]。土壤层有机碳密度值随林龄增大而降低,相似于贵州开阳杉木林[7]、相异于福建南平[4]和河南信阳杉木林[6],土壤有机碳密度值均低于相关文献[4,6-7]。培育大径级林和增加土壤有机碳含量是提高杉木林有机碳密度的有效途径。
研究发现杉木人工林乔木层、灌木层、草本层、凋落物层及土壤层有机碳密度值随密度升高而降低,同福建辛口[12]杉木成龄林乔木层植物有机碳密度变化相似、凋落物层有机碳密度变化相异。中龄林乔木层植物有机碳密度值随密度变化尚存有拐点效应[8],间伐[14]能改变林下植物有机碳密度值、主要受郁闭度和林内植物类型[13]以及土壤腐殖质层厚度的影响。
3.2林下植物对杉木人工林结构及土壤理化主要指标值的影响
研究发现灌木及草本层植物对杉木人工林有机碳密度的贡献率占1.83%,分析也发现相关显著的指标对数量率中,灌木层和乔木层及草本层、以及草本层和乔木层植物结构指标的相应值为12.50%、0%、8.33%,土壤A层和B层理化主要指标的相应值为10.93%。灌木层植物结构指标及多样性指数和土壤A层及B层理化主要指标的相应值为3.13%、3.13%,17.19%、3.13%,草本层植物的相应值为2.08%、2.08%,3.13%、7.81%,表明林下植物不仅是植被-土壤有机碳库的组成部分,其生长发育受乔木层植物影响、也影响土壤理化主要指标值[11、15]的大小。在合理调控乔木层植物密度的同时适度保留灌木及草本层植物,既有利于增加林地有机碳密度值、也有利于改善林地土壤质量。
研究发现杉木人工中龄林土壤部分理化指标值升高、成龄林土壤理化主要指标值降低,有别于相关文献[5,9,11],也有文献[9-11]表明近熟-成熟-过熟林阶段土壤理化主要指标值有一个波动过程,表明中-成龄林土壤有机碳含量等理化指标变化对土壤碳库的影响有进一步研究的必要。增加杉木凋落物[18]和改变其多样性[19]能显著改善土壤酶活性、有机碳分解及微生物碳密度,提高土壤水源涵养能力与林地养分循环速率[17],科学管理杉木人工林凋落物既利于森林防火、也有利于林地有机碳密度值和土壤生态功能的提高。
3.3长期定位观测是研究龄级及密度级和杉木人工纯林有机碳密度关系的有效手段
空间替代时间的方法是生态学研究的常用手段,本研究区主要立地因子和杉木人工林结构指标及有机碳密度(含量)值相关性不显著、有机碳密度主要受林龄和密度等林分结构指标的影响,由于缺少A1B1、A2B1和A3B3组合类型,无法进行龄级和密度的双因素方差分析,是本研究中实验设计的缺陷。建立在不同龄级和密度级变化之上的杉木人工林结构及有机碳密度变化结论仍然是可信的,更为科学的方法是长期定位观测[7-8,16,18-19]、以利于阐述林龄和密度变化对杉木林结构及有机碳密度影响的形成机理及科学制定营林调控措施。
参考文献
[1] 吴中伦. 杉木[M]. 北京:中国林业出版社,1984.
[2] 冯宗炜, 陈楚莹, 张家武, 等. 不同自然地带杉木林的生物生产力[J]. 植物生态学与地植物学丛刊, 1984,8(2): 93-100.
[3] Ma X Q, Liu C J, Ilvesniemi H, et al. Biomass, litter fall and the nutrient fluxes in Chinese fir stands of different age in subtropical China [J]. Journal of Forestry Research, 2002, 13(3): 165-170.
[4] 尉海东, 马祥庆. 中亚热带不同发育阶段杉木人工林生态系统碳贮量研究[J]. 江西农业大学学报, 2006,28(2): 239-243.
[5] 李平, 郑阿宝, 阮宏华, 等. 苏南丘陵不同林龄杉木林土壤活性有机碳变化特征[J]. 生态学杂志, 2011, 30(4):778-783.
[6] 邓华平, 李树战, 何明山, 等. 豫南不同年龄杉木林生态系统碳储量及其空间动态特征[J]. 中南林业科技大学学报, 2011, 31(8): 83-95.
[7] 刘延惠, 丁访军, 崔迎春, 等. 黔中地区不同林龄杉木人工林碳贮量及其分配特征[J]. 水土保持学报, 2015, 29(4):278-283.
[8] 姚利辉, 康文星, 赵仲辉, 等. 会同杉木人工林不同生长阶段植物固碳特征[J]. 生态学报, 2015, 35(4):1187-1197.
[9] 王丹, 戴伟, 王兵, 等. 杉木人工林不同发育阶段土壤性质变化的研究[J]. 北京林业大学学报, 2010, 32(3):59-63.
[10] 刘丽, 段争虎, 汪思龙, 等. 不同发育阶段杉木人工林对土壤微生物群落结构的影响[J]. 生态学杂志, 2009, 28(12):2417-2423.
[11] 郭琦, 王新杰, 衣晓丹. 不同林龄杉木纯林林下生物量与土壤理化性质的相关性[J]. 东北林业大学学报, 2014, 42(3):85-88, 98.
[12] 林开敏, 俞新妥, 何智英, 等. 不同密度杉木林分生物量结构与土壤肥力差异研究[J]. 林业科学, 1996, 32(5):385-392.
[13] 盛炜彤. 不同密度杉木人工林林下植被发育与演替的定位研究[J]. 林业科学研究, 2001, 14(5): 463-471.
[14]姚茂和, 盛炜彤, 熊有强. 杉木林林下植被及其生物量的研究[J]. 林业科学, 1991, 27(6): 644-648.
[15]林开敏, 俞新妥, 洪伟, 等. 杉木人工林林下植物对土壤肥力的影响[J]. 林业科学, 2001, 37(Sp.1):94-99.
[16] 宁晓波, 项文化, 王光军, 等. 湖南会同连作杉木林凋落物量20年动态特征[J]. 生态学报, 2009, 29(9):5122-5129.
[17] 张鹏, 王新杰, 衣晓丹, 等. 杉木不同生长阶段凋落物持水性与养分储量[J]. 东北林业大学学报, 2015, 43(10):58-62.
[18] 王晓峰, 汪思龙, 张伟东. 杉木凋落物对土壤有机碳分解及微生物生物量碳的影响[J]. 应用生态学报, 2013, 24(9):2393-2398.
[19] 汪思龙, 黄志群, 王清奎, 等. 凋落物的树种多样性与杉木人工林土壤生态功能[J]. 生态学报, 2005, 25(3):474-480.
[20]安和平, 金小麒, 杨成华. 板桥河小流域治理前期主要植被类型生物量生长规律及森林生物量变化研究[J]. 贵州林业科技, 1991, 19(4):20-34.
[21] 贺红早, 宋久祥, 刘盈盈, 等. 黔东南州杉木林生物量及其分配研究[J]. 广东农业科学, 2013, 28:58-60.
[22]李默然, 丁贵杰. 贵州黔东南主要森林类型碳储量研究[J]. 中南林业科技大学学报, 2013, 33(7):119-124.
[23]张金屯. 数量生态学[M]. 北京:科学出版社, 2004.
[24]国家林业局. 森林土壤分析方法(LY/T1210-1275-1999) [M]. 北京:中国标准出版社, 1999.
[25]余建英, 何旭宏. 数据统计分析与SPSS应用[M]. 北京:人民邮电出版社, 2003.
Influence of Age and Density Classes on Forest Structure Indicators and Organic Carbon Density ofCunninghamialanceolataPlantation in Southeastern Guizhou Province, China
Min Qili1, Zhang Xi2*, Zhang Zuoyu2, Tian Lianhong2,Yang Hanyuan3, Shi Yangwen4, Yang Yongzhang5
(1. Forestry Bureau of Jinpin County of Guizhou Province, Jinpin, Guizhou 556700; 2. Guizhou Provincial Academy of Forestry, Guiyang ,Guizhou 550005;3. Forestry Institute of Southeast Prefecture of Guizhou Province, Kaili, Guizhou 556000;4. Forestry Bureau of Liping County of Guizhou Province, Liping ,Guizhou 557300;5. Forestry Bureau of Rongjiang County of Guizhou Province, Rongjiang ,Guizhou 557200)
Abstract:Using the space instead of time and the combining method of field sample plot survey and indoor analysis, this article studied the structural indicators and organic carbon density changes of different age and density classes of artificial pure forest of Cunninghamia lanceolata in southeast Guizhou province, China. Results showed that with the age increase and density decrease of Chinese fir plantation: (1) Values of average diameter at breast height and height increased and density reduced, corresponding parameter values within the age or density classes were difference significantly. (2) Beside Pielou index,another herb plant diversity indices increased. Shrub plant diversity indices values increased with forest age increasing and decreased with density reducing. Pielou index values of shrub layer had significant difference in the different age or density classes. Gleason index values had no significant difference in the age classes and significant difference in the density classes, and Pielou index values had significant difference in the age classes and no significant difference in the density classes of herb layer. (3) Soil density increased, soil organic carbon content, etc, decreased. Soil density value was significant difference in age classes and no significant difference in density classes, soil organic carbon content, etc, had not significant difference in age or density classes. (4) Organic carbon density values of tree layer, shrub layer, litter layer and soil-vegetation layer increased, and organic carbon density values of herb layer and soil stratum (0-40 cm) went down in the age classes and went up in the density classes. Organic carbon density values of tree layer were significant difference in age classes and no significant difference in the density classes. (5) Age and density contribution rates for organic carbon density of tree layer were 99.06% and 0.94%, respectively. Soil organic carbon density was influenced by some factors of soil stratum. Vegetation organic carbon density was influenced comprehensively by part factors of vegetation-soil. (6) To foster large diameter and increase soil organic carbon content is an effective way to improve the organic carbon density of Chinese fir plantation.
Key words:Artificial pure forest of Cunninghamia lanceolata; Age classes; Density classes; Forest structure; Organic carbon density
收稿日期:2016-04-30
基金项目:贵州省科技厅攻关项目(黔科合SY字[2012]3010)和贵州省森林可持续经营管理试点项目(2012-18)资助
*通讯作者:张喜,E-mail:zhangxigzfa@tom.com。
中图分类号:S791.27
文献标识码:A
DOI.:10.13268/j.cnki.fbsic.2016.03.001
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
农业科技与信息(2022年12期)2022-08-09 05:32:24
中学生数理化·八年级物理人教版(2021年12期)2021-12-31 03:23:08
中学生数理化·八年级物理人教版(2021年12期)2021-12-31 03:23:02
贵州林业科技(2021年4期)2021-12-23 12:14:16
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
中学生数理化·八年级物理人教版(2019年12期)2019-05-21 07:26:36
中学生数理化·八年级物理人教版(2019年12期)2019-05-21 07:26:36
中学生数理化·八年级物理人教版(2017年12期)2017-04-18 12:59:39
