
栎空腔瘿蜂虫瘿在栓皮栎上的空间分布
2016-06-07 09:27:16王景顺张坤朋王相宏武三安
河南农业大学学报 2016年1期
王景顺,张坤朋,王相宏,武三安
栓皮栎(Quercus variabilis Blume)属于壳斗科Fagaceae栎属Quercus,分布范围广,经济和生态价值大,也是太行山区天然次生林的重要组成树种,在林业生产中起着重要作用[1]。栎空腔瘿蜂(Trichagalma glabrosa Pujade-Villar)2012年被定名为新种[2],该虫主要危害栓皮栎叶片,近年来,在南太行山区栓皮栎林内爆发成灾[3-4],受害叶片虫瘿密密麻麻,使叶片枯黄、灰褐、畸形或提前脱落,使树势不断衰弱,严重影响生态防护林安全及自然生态景观。研究发现,栎空腔瘿蜂有性世代虫瘿出现在栓皮栎雄花序上,成虫4月中旬羽化出瘿,由于该阶段羽化成虫既有雌成虫又有雄成虫,因此,称该阶段虫瘿称为有性世代虫瘿,雌雄交配后雌虫在栓皮栎嫩叶叶脉产卵,5月上旬虫瘿开始形成,虫瘿主要出现在叶片正面,6月下旬虫瘿发育成熟,9月上旬幼虫开始化蛹,11月成虫开始羽化出孔,由于此时羽化成虫全部为雌成虫,因此,称该阶段虫瘿为无性世代虫瘿。空间格局反映了种群的个体在某一时刻的行为习性和环境对种群的影响,是昆虫种群的重要特征之一[5-7]。由于种群的生物学特性,在特定的生境条件下形成了种群的空间分布形式,研究昆虫的空间格局有助于了解昆虫的生态特性,并能揭示种群的空间结构及种群分布规律,为此,本研究对南太行山栎空腔瘿蜂种群空间分布格局进行调查,并揭示空间分布格局形成的原因,从而为该虫的调查取样方式与防治决策提供必要的理论依据。
1 研究方法
1.1 调查地概况与调查时间
试验点设在河南省林州市姚村镇水河村的栓皮栎林内,地理坐标为北纬 36°13',东经 113°47',试验点面积2 hm2,栓皮栎 10~15 a,树高4~15 m,林分郁闭度0.65左右,栎空腔瘿蜂在近几年发生较为严重。
调查时间为2013年,分2次进行:1次是在4月中旬,调查栓皮栎花序上有性世代虫瘿数量;另1次是在5月中旬,调查无性世代虫瘿数量,此时虫瘿处于发育初期,即虫瘿初始期,调查初始期虫瘿数量。
1.2 调查方法
1.2.1 有性世代虫瘿数量调查方法 在试验区采用逐株系统调查法,共调查50(10×5)株。调查时每株树冠分上、中、下3层,每层又分为东、南、西、北4个方位,每个方位取一个标准枝条,系统调查各枝条所有花序的有性世代虫瘿数量。
1.2.2 无性世代虫瘿调查方法 在试验区采用逐株系统调查法调查50(10×5)株,调查每株树冠上、中、下3层,每层又分为东、南、西、北4个方位,每个方位取一个标准枝条,在每个枝条上取虫瘿数量最多的3片叶,统计该3片叶的虫瘿数量之和。
1.3 空间分布型的测定方法
1.3.1 聚集度测定的指标
1.3.1.1 I指标
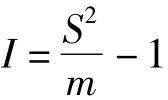
式中:S2为样本方差,m为平均密度,当I<0时为均匀分布,当I=0时为随机分布,当I>0时为聚集分布。
1.3.1.2 m*/m 指标
m*为平均拥挤度,表示生物个体在一个样方中的平均邻居数,它反映了样方内生物个体的拥挤程度。
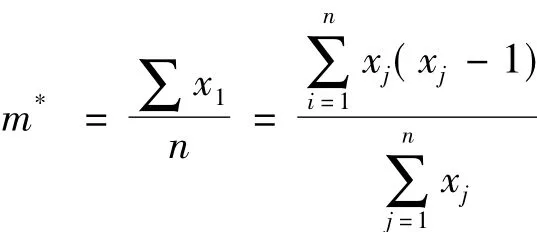
式中:xj为第j个样方的个体数,n为样方总数。判断指标为平均拥挤度m*与其平均值m之比值,即m*/m的比值,当 m*/m<1时为均匀分布;当m*/m=1时为随机分布;当m*/m>1时为聚集分布。
1.3.1.3 扩散系数 C
该指标C=S2/m用于检验种群是否偏离随机型。当C<1时为均匀分布,当C=1时为随机分布,C>1时为聚集分布。
1.3.1.4 K 值及 CA值法
在负二项分布中,参数K值作为种群聚集强度的1个指标,K值大小反映种群聚集程度的大小。K=m2/(S2-m),当K<0时为均匀分布,当K+∞时为随机分布,当K>0时为聚集分布。CA为K值倒数,即用1/K来表示,CA=1/K,当 CA<0,种群为均匀分布;CA=0,种群为随机分布;CA>0,种群为聚集分布。
1.3.2 Iwao m*-m直线回归法 根据Iwao提出的回归方程m*=α+βm。式中α表示分布的基本成分,或按大小分布的平均拥挤度。当α=0时,表示分布的基本成分为单个个体;当α>0表示种群个体间相互吸引,分布的基本成分为个体群;当α<0表示种群个体间相互排斥。β表示基本成分的空间分布形式,当β=1时种群基本成分为随机分布;β>1时种群基本成分为聚集分布;β<1时种群基本成分为均匀分布。而α与β的不同组合可提供种群的不同分布类型信息。当α=0,β=1时,种群分布型为随机型;当①α>0,β=1;②α=0,β>1;③α>0,β>1时,种群分布型为聚集分布。其中属于①时为泊松正二项分布、核心分布或奈曼-A型分布;属于②时,种群为具有公共K的负二项分布;属于③时,种群为一般的负二项分布[8]。
1.3.3 聚集原因分析 Blackith提出聚集均数(λ)分析害虫聚集原因,公式为[5,7]:
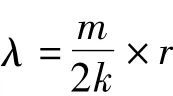
当λ<2时,昆虫聚集是由环境条件引起;当λ≥2时,昆虫聚集是由环境条件和昆虫本身聚集习性或由其中1个因素引起。公式中m为平均密度;k为负二项分布k值;r为具有自由度等于2k的分布函数,即r等于“x2值表”中自由度等于2k与概率(P=0.5)对应的值。
2 结果与分析
2.1 栎树空腔瘿蜂有性世代虫瘿和无性世代虫瘿分布型
2.1.1 聚集度指数测定 以株为抽样单元统计有性世代虫瘿和无性世代虫瘿密度,计算平均密度(m)和方差(S2),采用以上5种聚集度指标测定栎空腔瘿蜂空间格局,结果见表1。由表1可知,I>0,m*/m >1,CA>0,C >1,K >0,表明无论是栎空腔瘿蜂有性世代虫瘿还是无性世代虫瘿均呈聚集分布。
2.1.2 回归模型分析法 根据Iwao建立平均拥挤度(m*)与平均数(m)的回归模型[8],按照10组资料,分别求得有性世代虫瘿和无性世代虫瘿平均密度 mi(i=1,2...)和平均拥挤度 mi*(i=1,2...),见表2,建立m*-m 的回归。有性世代虫瘿回归方程为:m*= -4.444+1.374m,α =-4.444<0,说明有性世代虫瘿个体间相互排斥,β=1.374大于1,说明有性世代虫瘿为聚集分布。无性世代的回归方程为:m*=-28.465+1.239m,α = -28.465 <0,说明无性世代虫瘿个体间相互排斥,β=1.239>1,说明无性世代虫瘿为聚集分布。

表1 林内栎空腔瘿蜂虫瘿空间分布型参数Table 1 Spatial distribution pattern parameters of galls
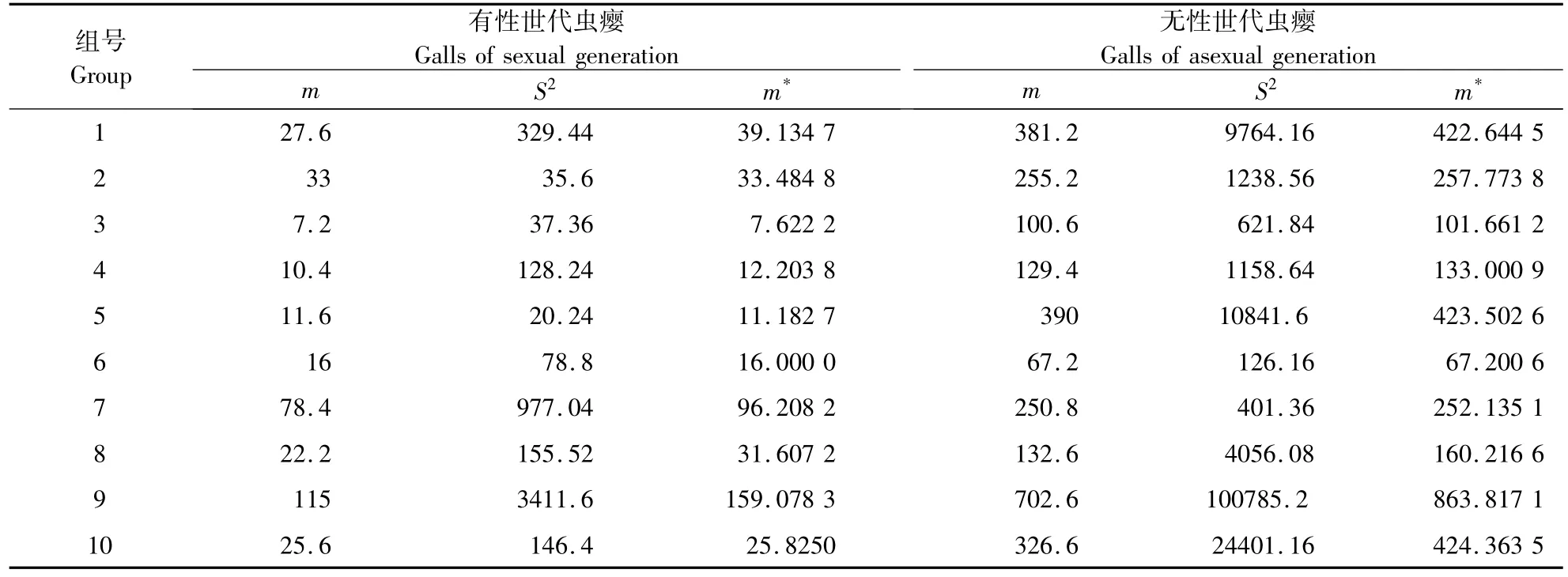
表2 栎空腔瘿蜂虫瘿平均密度m及拥挤度m*的计算Table 2 The calculation of average density(m)and congestion degree of gall wasp of Trichagalma glabrosa(m*)
2.1.3 虫瘿聚集原因分析 由表2数据计算,有性世代虫瘿的18.89>2;无性世代虫瘿 k=2.71,γ =4.77;λ =明栎空腔瘿蜂有性世代虫瘿以及无性世代虫瘿由环境条件和其本身聚集习性或由其中一个因素引起。
2.2 栎树空腔瘿蜂有性世代和无性世代虫瘿在树冠不同方位分布
在树冠不同方位上,对东、南、西、北4个水平方位和上、中、下3个垂直方位虫瘿密度进行比较,按不同方位计算有性世代和无性世代虫瘿的m值以及m* -m回归的α,β,r值,结果见表3和表4。由表3可以看出,栎空腔瘿蜂有性世代虫瘿在栓皮栎树冠各方位均为聚集型,水平方位虫瘿密度方差分析显示,各方位间虫瘿密度差异未达显著水平;在垂直分布上,上、中、下层均为聚集型,但虫瘿密度以下层最高。分析认为,栎空腔瘿蜂无性世代成虫飞行能力较差,每年从11月到12月为无性世代成虫羽化期,当成虫羽化后,一般选择树冠中下部的花芽产卵,树冠高大,又常多风,不利于其产卵。无性世代虫瘿在树冠各层均为聚集型(表4),聚集度相差不大,虫瘿密度上层最高,其差异达到显著水平。分析认为,有性世代成虫喜光,飞行能力较强,因为树冠上部的光照较强、生长较旺,比较适合成虫产卵。
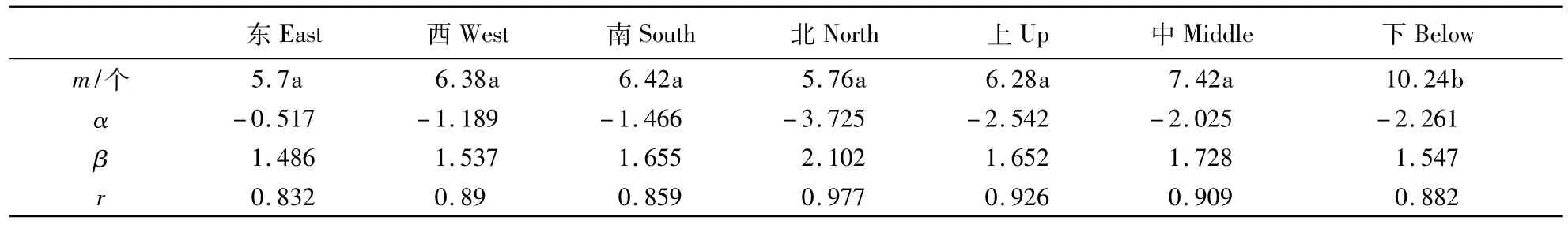
表3 栎空腔瘿蜂有性世代虫瘿栓皮栎不同方位上空间分布Table 3 Spatial distribution of sexual generation galls at different position of Quercus variabilis
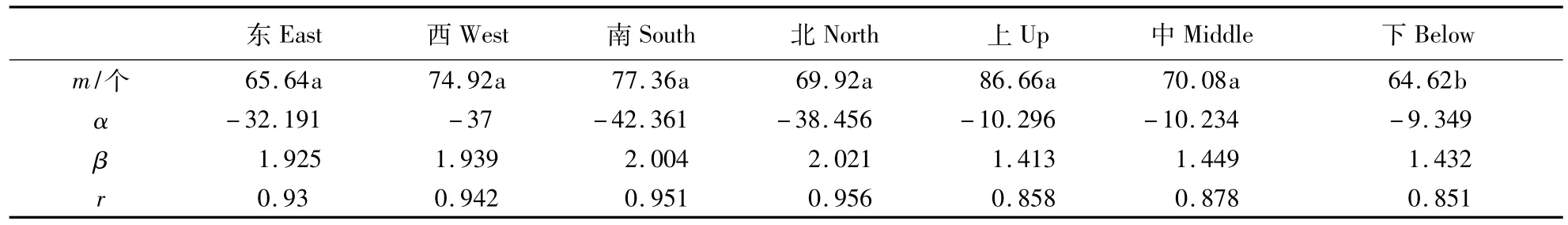
表4 栎空腔瘿蜂无性世代虫瘿在栓皮栎上不同方位空间分布Table 4 Spatial distribution of asexual generation galls at different position of Quercus variabilis
3 结论与讨论
种群空间分布型不仅受环境因素、生物学特性影响,而且还受发育虫态、季节变化、种群密度及农药使用等多个因素影响[9]。栎空腔瘿蜂在栓皮栎的空间分布反映其生物学特性及扩散能力,本研究中,无论是有性世代虫瘿还是无性世代虫瘿均为聚集分布,虫瘿在树冠东、西、南、北4个水平方位上,虫瘿密度差异不显著,而在树冠垂直分布上,有性世代虫瘿密度在树冠下层较大,与树冠上层虫瘿密度差异显著。而有性世代虫瘿是由上一年无性世代成虫产卵决定的,由于该成虫的飞翔能力不强,吊飞试验发现该虫基本不飞行,因此,在其成虫发生期的天气状况可能会影响其产卵位置,也就解释了有性世代虫瘿在树冠下层较多的原因。有性世代成虫在4月中下旬出现,成虫飞行能力较强,喜欢光,因此,也印证了无性世代虫瘿密度在树冠上层较高的原因。
野外调查发现,在栓皮栎林缘或道路两旁的虫瘿密度一般都比林中的种群密度大,孤立木以及幼树发生较多。分析原因认为,这些地方光照条件好,由于成虫喜光,从而造成了虫瘿分布规律的差异。近些年,随着旅游业发展,在山区不断修路,特别是生境破碎化严重地带,创造了更多的林缘环境,使栓皮栎林内小气候发生了改变,从而造成了种群数量的爆发。运用传统的生物学统计方法研究空间格局,只是获得一些空间分布的定性信息,无法知道分布的位置和程度。近年来,不断有学者运用地统计学方法研究种群空间分布[10-13],这样不仅可对区域化变量的空间相关进行定量化描述,还可对未抽样位置进行估值,可以更好地反映昆虫种群空间格局。探索新的统计方法以及完善和综合运用多种空间格局分析方法,使其相互补充,必将为更准确了解昆虫的空间结构提供理论依据。
[1] 万猛,田大伦,樊巍.太行山南麓栓皮栎群落结构特征分析[J].河南农业大学学报.2009,43(2):139-144.
[2] PUJADE-VILLAR J,WANG J.A new species of the genus Trichagalma Mayr from China(Hym.:Cynipidae)[J].Orsis,2012(26):91 -101.
[3] 王景顺,王相宏,PUJADE-VILLAR J,等.栎空腔瘿蜂形态及生物学特性[J].中国森林病虫,2013(1):8-11.
[4] 王景顺,王相宏,武三安.栎空腔瘿蜂自然种群生命表的组建与分析[J].中国农学通报.2014,30(28):29-33.
[5] 丁岩钦.昆虫数学生态学[M].北京:科学出版社,1994:22-69.
[6] 郑汉业,夏乃斌.森林昆虫生态学[M].北京:中国林业出版社,1993:120-167.
[7] 张志春,王楠,李娟,等.杨扇舟蛾卵和幼虫的空间分布型及抽样技术[J].昆虫知识,2006,43(2):192-195.
[8] 张国安,赵惠燕.昆虫生态学与害虫预测预报[M].北京:科学出版社,2012:63-73.
[9] 矫振彪,陆宴辉,吴孔明.棉田绿盲蝽的空间分布型及其抽样模型[J].应用昆虫学报.2012,49(3):605-609.
[10]宗世祥,骆有庆,许志春,等.沙棘木蠹蛾蛹的空间分布[J].生态学报.2006,26(10):3232-3237.
[11]宗世祥,骆有庆,许志春,等.沙棘木蠹蛾卵和幼虫空间分布的地统计学分析[J].生态学报.2005,25(4):831-836.
[12]闫香慧,赵志模,刘怀,等.白背飞虱若虫空间格局的地统计学分析[J].中国农业科学,2010,43(3):497-506.
[13]夏鹏亮,王瑞,王昌军,等.恩施烟区无翅桃蚜在烤烟田空间动态的地统计学分析[J].生态学报,2014,5(5):1198-1204.
猜你喜欢
现代苏州(2022年5期)2022-03-21 22:57:09
南方农业·下旬(2020年1期)2020-06-01 07:47:32
现代园艺(2019年24期)2019-01-07 01:19:36
科学之谜(2018年11期)2018-12-05 07:57:16
现代农业科技(2009年21期)2018-09-03 00:00:00
农村百事通(2017年18期)2017-02-08 11:23:54
中国现代中药(2016年9期)2016-09-25 08:38:10
新疆农业科学(2016年6期)2016-07-23 01:51:10
IT时代周刊(2015年9期)2015-11-11 05:51:51
少儿科学周刊·儿童版(2014年1期)2014-03-10 06:23:18
