
干旱胁迫对4种棕榈植物幼苗光合特性及抗氧化酶活性的影响
2016-05-30 07:03:36阮志平唐源江曾美涓
热带作物学报 2016年10期
阮志平 唐源江 曾美涓

摘 要 以4种棕榈植物幼苗为试材,通过盆栽控水试验,研究了干旱胁迫对其光合特性和抗氧化酶活性的影响。结果表明,随着干旱程度加剧,4种棕榈植物的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(E)均呈逐渐下降趋势,但下降幅度在不同种及不同胁迫处理之间存在差异。在轻度干旱胁迫下,气孔限制是皇后葵和砂糖椰子Pn下降的主要原因,而东澳棕和圆叶蒲葵Pn下降的主要原因是非气孔限制因素;在中度和重度干旱胁迫下,4种棕榈植物Pn下降的主要原因均为非气孔限制因素。4种棕榈植物的平均水分利用效率(WUE)依次为皇后葵>东澳棕>砂磄椰子>圆叶蒲葵。轻度干旱胁迫下4种植物的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性均高于对照,中度和重度胁迫下均低于对照。复水后各处理均可恢复达对照水平。4种棕榈植物以皇后葵抗干旱能力最好,其次为东澳棕和砂糖椰子,圆叶蒲葵最差,对干旱环境最为敏感。
关键词 棕榈植物;干旱胁迫;光合作用;抗氧化酶
中图分类号 Q948 文献标识码 A
Abstract A pot experiment was conducted to study the effects of different drought stress(normal irrigation, light drought, moderate drought, and severe drought)on the photosynthetic characteristics and antioxidant enzymes of the seedling in four species(Syagrus romanzoffiana, Carpentaria acuminata, Arenga pinnata and the activities of Livistona rotundifolia)of Palmaceae family. The results showed that their net photosynthetic rate(Pn), stomatal conductance(Gs), and transpiration rate(E), compared with those under normal irrigation, gradually decreased with increasing drought stress. However, the reducing rates were different among the four species and different treatments. The intercellular CO2 concentration (Ci) decreased, and the stomatal limitation value(Ls)increased under light drought stress in S. romanzoffiana and A. pinnata,thus the decrease of their Pn was mainly caused by stomatal limitation. But when Ci increased, and Ls decreased under the same stress in C. acuminata and L. rotundifolia, therefore the decrease of these Pn was mainly caused by non-stomatal limitation. However, the Ci increased while Ls decreased under moderate and severe stress in the four species, thus decrease of all Pn examined was mainly caused by non-stomatal limitation. Water use efficiency(WUE)of the four species was: S. romanzoffiana>C. acuminata>A. pinnata >L. rotundifolia. Superoxide dismutase(SOD), peroxidase(POD), and catalase(CAT)activities of the four species increased under light drought stress, decreased under moderate and severe drought stress, and renewed their activities to the level of normal irrigation after re-watering in all treatments. Thereby, S. romanzoffiana possessed the. strongest capability against drought stress among the four species, then C. acuminata and A. pinnata, and lastly L. rotundifolia although L. rotundifolia possessed the best sensitivity to the drought environment.
Key words Palms;Drought stress;Photosynthesis;Antioxidant enzyme
doi 10.3969/j.issn.1000-2561.2016.10.011
棕榈科(Palmaceae)植物是世界上3个最重要的经济植物类群之一,也是最为独特的观赏植物[1]。目前,在中国长江以南的亚热带到热带地区已广为引种,特别是位于南亚热带到热带的广大东南沿海地区,已成为园林景观营造的主要树种,且不乏“椰岛风情”为主题的独特景观。关于棕榈植物的研究目前多集中在分类、引种栽培、植保、抗寒、园林应用等方面[2-7]。在抗旱光合生理及抗氧化酶的研究方面,Repellin等[8]认为气孔导度、净光合速度、胞间CO2值等叶片气体交换参数不能作为椰子鉴定抗旱能力的指标。不过没有进一步的研究佐证这一结论以及在棕榈植物中是否具普遍性;孙程旭等[9]和Cha-um等[10]发现随着干旱胁迫时间的延长,油棕幼苗的SOD和POD酶活性出现先上升后下降变化趋势。曹建华等[11]则发现在一定干旱胁迫下,油棕的SOD和POD活性先下降,而随着胁迫时间延长,POD活性则上升,认为油棕对干旱的胁迫性有一定的限度。陈歆等[12]在研究土壤水分胁迫对槟榔幼苗的影响时发现光合速率、蒸腾速率、气孔导度和叶绿素含量均下降,SOD、POD活性也是先上升后下降变化趋势,而CAT则表现先下降后上升变化。随胁迫时间延长,SOD和POD活性下降现象越明显。综合来看,不同棕榈植物受到干旱胁迫后的光合生理及抗氧化酶活性的变化存在差异[7],然而由于目前研究过的棕榈植物材料有限,对棕榈植物抗旱光合生理生化的认识仍缺乏系统性,而同一条件下开展多种材料的比较研究尚未见报道。为此,本文通过盆栽控水试验,比较研究干旱胁迫及复水处理对4种重要观赏棕榈植物的光合参数和抗氧化酶活性的影响,以期为深入探究棕榈植物抗旱生理机制及响应规律积累数据,为棕榈植物的引种和抗旱筛选、景观配置和人工群落构建等生产实践工作提供理论依据。
1 材料与方法
1.1 供试材料
于2014年10月上旬将一年生皇后葵[Syagrus romanzoffiana](Cham.)]Glassm.]、东澳棕[Carpentaria acuminata(H.Wendl. Et Drude)Becc.]、砂糖椰子[Arenga pinnata(Wurmb)Merr.]、圆叶蒲葵[Livistona rotundifolia(Lam.)Mart.]4种植物盆栽苗从厦门市园林植物园移至华侨大学厦门校区园艺实验室通风遮雨的塑料棚内,各种幼苗生长良好且长势基本一致(每株6~片叶),无病虫害。试验用盆为高25 cm,直径 20 cm的塑料盆,栽培基质按腐叶土 ∶ 园土=1 ∶ 1配比而成,每盆种植3 株小苗,按生产栽培的管理水平,尽可能保持每盆供水和盆内土壤湿度的一致性,使其正常生长。
1.2 方法
1.2.1 试验设计 材料移入华侨大学厦门校区实验室正常养护2周后,将每盆充分灌水使土壤含水基本保持一致,待土壤自然落干后进行干旱胁迫处理,此时的田间持水量为38.62%。每盆中的水量用称重法控制,每天于17:30 向盆中补充水分。每个品种试验处理设4 组:(1)对照组(CK),按生产栽培正常供水,为田间持水量的70%~75%;(2)轻度干旱胁迫(Light drought stress, LS),为田间持水量60%~65%; (3)中度干旱胁迫(Moderate drought stress, MS),为田间持水量45%~50%;(4)重度干旱胁迫(Severe drought stress, SS),为田间持水量30%~35%。当各处理的土壤水分梯度形成时开始干旱胁迫,胁迫处理30 d后,测定各项指标。试验期间的环境空气相对湿度控制在均值为60%~70%, 气温均值最低22 ℃,最高32 ℃,有效光合辐射(685±50)μmol/(m2·s)。设定二氧化碳供应为开放式气路(385±20)μmol/mol。干旱胁迫结束后,开始按正常管理给各处理复水(Re-watering),为保证数据的稳定和可靠,分别于复水后第5天测量各项指标。每次测量时间为早上9:00-11:00,测量时随机选择长势及位置一致、无病虫害、向阳面的功能叶,每叶片重复3 次,取平均值。将测定后的叶片液氮速冻-80 ℃保存, 用于抗氧化酶活性的测定。
1.2.2 试验方法 采用CI-310便携式光合仪(美国CID公司),测定叶片的净光合速率[Pn,μmol/(m2·s)],蒸腾速率[E,μmol/(m2·s)], 叶片气孔导度[Gs,mmol/(m2·s)],胞间CO2浓度(Ci,μmol/mol)等光合参数。通过公式计算水分利用效率(WUE)=Pn/E[μmol/(m2·s)]和气孔限制值Ls(%)=1-Ci/Ca。超氧化物歧化酶(SOD)活性参照李合生等[13]方法测定;过氧化物酶(POD)活性采用Amalo等[14]的方法测定;过氧化氢酶(CAT)活性采用Abei等[15]的方法测定;均重复测定3次,取平均值。
1.3 数据处理
所用数据均用Microsoft Excel 进行计算制图,用SPSS 19.0单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同处理的差异,均值差的显著性水平设定为0.05。
2 结果与分析
2.1 干旱胁迫与复水处理对4种棕榈植物幼苗光合参数的影响
2.1.1 干旱胁迫与复水处理对4种棕榈植物幼苗Pn的影响 如图(1-A)可知,4种棕榈植物的Pn均随干旱胁迫程度的加剧呈下降趋势,与对照(CK)相比不同种不同处理变化的幅度不同。在LS下,各植物Pn均呈小幅下降;而在MS和SS下,下降幅度较明显。各处理复水后,各植物Pn均有一定程度的恢复,其中轻度干旱后复水(Re-watering for LS, RLS)4种植物的Pn均可恢复到与对照组相当的水平;中度干旱后复水(Re-watering for MS, RMS)和重度干旱后复水(Re-watering for SS, RSS)各植物Pn均仅可部分恢复。在RMS下,4种植物的恢复性强弱排序为东澳棕>皇后葵>砂糖椰子>圆叶蒲葵;而在RSS下,4种植物的恢复性强弱排序为皇后葵>东澳棕>砂糖椰子>圆叶蒲葵。方差和LSD分析显示,轻度干旱胁迫及复水处理后4种棕榈植物的Pn与各对照组均差异不显著,而中度干旱胁迫和重度胁迫及复水处理后Pn与各对照组均具显著性差异。
2.1.2 干旱胁迫与复水处理对4种棕榈植物幼苗Gs的影响 干旱胁迫下,随着干旱程度不断加深,4种棕榈植物的气孔导度Gs的变化表现不同(图1-B),但趋势与Pn的一致,即随着干旱胁迫加剧而呈下降趋势。方差和LSD分析显示,皇后葵的Gs除SS处理与对照具显著差异外,其它各处理均差异不显著;东澳棕的Gs则仅RLS处理与对照无显著性差异,其它各处理均差异显著,尤以SS下,Gs下降幅度最明显。砂糖椰子在LS和RLS下,其Gs与对照无显著差异,其余各处理的Gs与对照具显著差异。圆叶蒲葵的Gs在LS、RLS和RMS下与对照无显著差异,其余各处理与对照均具显著。可见,在LS下,4种棕榈植物的Gs并未受到明显影响,具有较好的恢复性,但圆叶蒲葵和东澳棕的Gs对干旱胁迫十分敏感,尤以圆叶蒲葵更甚;在MS下,皇后葵和圆叶蒲葵的Gs复水后均恢复性较好,说明未受到明显影响,而东澳棕和砂糖椰子的Gs虽恢复性良好,但受到了显著影响;在SS下仅皇后葵的Gs无明显影响,仍保持较好的恢复性。4种植物按Gs的恢复性强弱排序为皇后葵>圆叶蒲葵>东澳棕>砂糖椰子。
2.1.3 干旱胁迫与复水处理对4种棕榈植物幼苗E及WUE的影响 在干旱胁迫下,4种棕榈植物的E变化与Pn及Gs相似,即E随着干旱胁迫的程度加剧呈下降趋势(如图1-C)。方差和LSD分析显示,除皇后葵在LS下的E与对照组无显著差异外,皇后葵的MS和SS处理及其他3种植物的LS、MS和SS各处理的E与各对照组均具显著性差异。复水处理后4种棕榈植物的E恢复程度各异,皇后葵的各复水处理后的E均与对照无明显差异,砂糖椰子仅RLS处理与对照无显著差异,其它2个种各复水处理后E与对照组差异显著。从4种棕榈植物的水分利用效率(WUE)来看(如图1-D),皇后葵在LS、MS及RLS下,各WUE与对照组无明显差异,其它各处理与对照均差异显著。东澳棕在MS及SS下WUE与对照组有显著差异,其它各处理均无显著差异。砂糖椰子仅在LS及RLS下WUE与对照无显著差异。圆叶蒲葵除在RLS下无显著差异外,其它各处理的WUE均与对照差异显著。
2.1.4 干旱胁迫与复水处理对4种棕榈植物幼苗Ci和Ls的影响 在干旱胁迫下,随着干旱程度的加剧,4种植物的Ci和Ls值变化如图1(E-F)。皇后葵在LS下的Ci明显比对照低,Ls值明显比对照高,其它各处理的Ci与Ls与对照均无显著差异;东澳棕各处理下的Ci和Ls与对照均差异不显著;砂糖椰子在LS下的Ci和Ls变化与皇后葵一致,其它各处理中仅在SS下Ci和Ls与对照具显著差异。圆叶蒲葵的Ci在各处理下均高于对照,且与对照具显著差异,而Ls值则均低于对照且各处理与对照均差异显著。
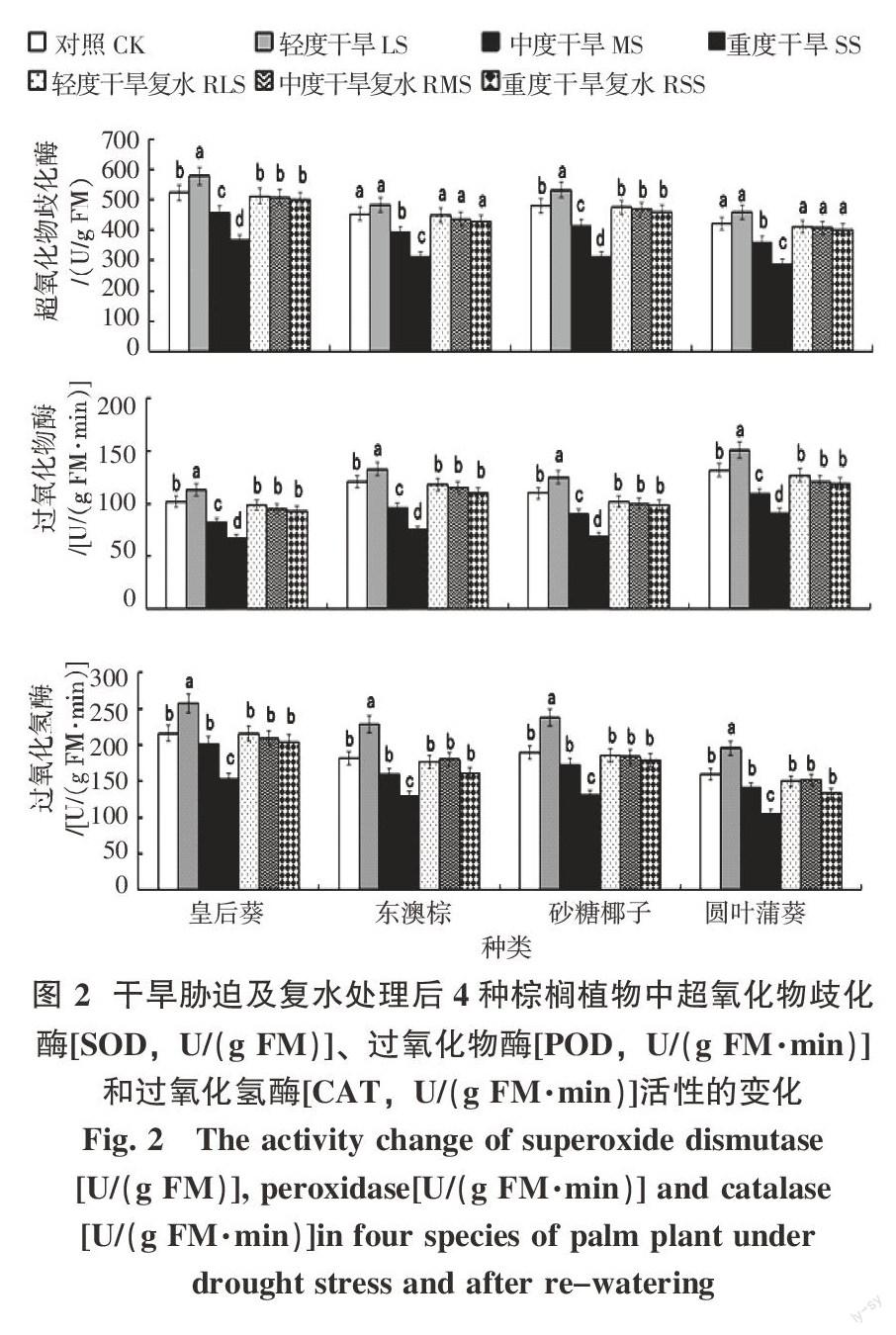
2.2 干旱胁迫与复水处理对4种棕榈植物幼苗抗氧化酶活性的影响
由图2可以看出,在干旱胁迫下,随着胁迫程度加剧,4种棕榈植物的SOD、POD和CAT活性均呈先上升(在LS下)后下降(在MS和SS下)的趋势。方差和LSD分析显示,在LS下,皇后葵和砂糖椰子的SOD活性与对照差异显著,东澳棕与圆叶蒲葵的与对照差异不明显,而在MS和SS下,4种棕榈植物的SOD活性与对照均具显著性差异;4种植物的POD活性则在LS、MS和SS下与对照均有显著差异;CAT活性在LS和SS下,与对照差异显著,而在MS下差异均不明显。复水后,4种植物各处理的SOD、POD和CAT活性均基本得到恢复,且与对照间无显著差异。
3 讨论
3.1 光合参数与抗旱性的关系
水是光合作用原料之一,没有水,光合作用无法进行。而缺水影响光合作用主要是通过气孔关闭、光合产物输出减慢、光合机构受损等间接影响[16]。这些原因可以概括为气孔因素和非气孔因素。气孔因素实际上主要是植物的气孔调节能力,植物通过气孔调节控制体内与外界环境之间的气体交换和水分蒸腾[17-18],是植物适应环境与抵御干旱的重要调节机制。非气孔因素则包括一系列复杂的生理生化过程和结构机制。一般认为Ci和Ls可以作为区分光合速率下降的气孔和非气孔因素的判断[19]。本研究显示,在LS下,皇后葵和砂糖椰子Ci下降,Ls增大,说明气孔限制因素是引起Pn下降的主要原因,而东澳棕和圆叶蒲葵Ci升高,Ls减小,说明引起Pn下降的主要原因是非气孔因素;另在LS及RLS处理下,4种棕榈植物的Pn与对照均无明显差异。这说明在LS下,无论是气孔因素还是非气孔因素导致Pn下降,都是一些适应性反应,未产生不可逆转的影响,这与陈昕等[20]、张诚诚等[21]及郭素枝等[22]研究相一致,他们从不同角度证明在LS下,叶片结构、光合色素、光合参数等大多与对照无显著差异。在MS和SS下,4种棕榈植物的Ci均上升,Ls下降,非气孔限制成为Pn下降的主要原因。从它们Pn的RMS和RSS处理结果看,虽均都可以得到一定程度的恢复,但与对照差异显著。这可能说明光合系统已产生了不可逆的损害。此外,本研究显示,在干旱胁迫条件下, 4种棕榈植物的Pn、Gs和E具有相同的变化趋势,即均随干旱胁迫强度加剧而下降,这与陈歆等[12]的结果一致。但不同种不同干旱胁迫下的Pn、Gs和E值变化差异较大且缺乏规律性。因此,仅从Pn、Gs和E等受干旱影响的变化情况来判断4种棕榈植物的抗旱能力确实困难,这与Repellin等[8]的结论相符。然而从复水后Pn、Gs和E的恢复能力强弱看,皇后葵恢复力最强,其次是东澳棕和砂糖椰子,再则是圆叶蒲葵,因此,结合干旱胁迫与复水处理整个过程的光合参数变化情况,是可以对4种棕榈植物的抗旱性作一个大致筛选的。
3.2 抗氧化酶活性与抗旱性的关系
SOD、POD和CAT是植物组织内重要的抗氧化酶,它们通过清除超氧阴离子自由基、羟自由基和过氧化氢来减少活性氧对细胞膜的伤害、减轻膜质过氧化和稳定膜的透性[23-26]。在非环境胁迫下,植物细胞内活性氧的产生与清除处于动态平衡之中,环境胁迫时活性氧产生与其清除之间的动态平衡将被打破,致使活性氧过量积累[27-28]。因此抗氧化系统对活性氧的清除能力是衡量植物对逆境抵抗的关键因素之一[29]。本研究显示,在LS下,4种棕榈植物的SOD、POD和CAT活性均出现上升,而在MS和SS下,均呈下降现象,且在SS下显著下降,三种酶的活性均呈LS>MS>SS,这表明在LS下,植物体内的超氧阴离子自由基、羟自由基和过氧化氢等不断产生,促使4种棕榈植物启动了相应的适应性反应,SOD、POD和CAT活性相应增强,以消除活性氧的积累,以求达到一种新的动态平衡,但随着胁迫继续加深,在MS和SS下,活性氧的积累超出了4种植物的抗氧化酶系统可能清除的范围,从而整个抗氧化酶系统功能降低。因此,LS下SOD、POD和CAT的表达水平可能是4种棕榈植物干旱胁迫下的容许限度,一旦进入MS和SS,防御功能将逐渐下降直至失去。但从复水的结果看,4种棕榈植物的抗氧化酶系统在各处理条件下均具有良好的恢复性。这可能说明4种棕榈植物的抗氧化酶系统具有较好的干旱适应能力,另一方面也可能与试验中干旱胁迫处理的时间长度较短有关。
参考文献
[1] 刘海桑. 观赏棕榈[M]. 北京: 中国林业出版社, 2002: 1.
[2] 林有润. 略论棕榈科与新分出的省藤科的系统分类、 演化、 区系地理及主要的经济用途[J]. 植物研究, 2002, 22(3): 341-365.
[3] 蔡邦平, 王振忠, 张秀英. 厦门观赏棕榈植物引种影响因素的研究[J]. 林业科学, 2005, 41(3): 63-67.
[4] 林秀香, 陈振东. 我国棕榈科植物的研究进展[J].热带作物学报, 2007, 28(3): 115-119.
[5] 孙程旭, 刘立云, 李 杰,等. 棕榈科植物抗寒生理研究进展及展望[J]. 中国农学通报, 2008, 24(11): 475-477.
[6] 王莉芳, 熊忠臣, 欧蒙维, 等. 棕榈植物在桂林的引种和适应性研究[J]. 广西科学, 2012, 19(2): 196-200.
[7] 刘艳菊,曹红星. 棕榈科植物抗寒、抗旱生理生化研究进展[J]. 中国农学通报, 2015, 31(22): 46-50.
[8] Repellin A, Braconnier S, Laffray D, et al. Water relations and gas exchange in young coconut palm(Cocos nucifera L.)as influenced by water deficit[J]. Canadian Journal of Botany, 1997, 75(1): 18-27.
[9] 孙程旭, 曹红星, 马子龙, 等. 干旱胁迫对油棕幼苗生理生化特性的影响[J]. 西南农业学报, 2010, 23(2): 383-386.
[10] Cha-um S, Yamada N, Takabe T, et al. Physiological features and growth characters of oil palm (Elaeis guineensis Jacq.) in response to reduced water-deficit and rewatering[J]. Australian Journal of Crop Science, 2013, 7(3): 432-439.
[11] 曹建华, 李晓波, 陶忠良, 等. 油棕新品种对干旱胁迫的生理响应及其抗旱性评价[J]. 热带农业科学, 2014, 34(7): 27-32.
[12] 陈 歆. 土壤水分胁迫对槟榔幼苗生理生态特性的影响[D]. 海口: 海南大学, 2010.
[13] 李合生. 植物生物化学实验原理与技术[M]. 北京: 高等教育出版社, 2000: 164-165.
[14] Amalo K, Chen GX, Asade K. Separate assays specific for ascorbate peroxidase and guaiacal peroxidase and for the chloroplastic and cytosolic isozymes of ascorbate peroxidase implants[J]. Plant Cell Physiology, 1994, 35: 497-504.
[15] Aebi H. Catalase in vitro[J]. Methods in Enzymology, 1984, 105: 121-126.
[16] 李合生. 现代植物生理学[M]. 北京: 高等教育出版社, 2002: 135.
[17] Hetherrington A M, Woodward F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 434: 901-908.
[18] 杨再强, 谭 文, 刘朝霞, 等. 土壤水分胁迫对设施番茄叶片气孔特性的影响[J]. 生态学杂志, 2015, 34(5): 1 234-1 240.
[19] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annuals Reviews Plant Physiology, 1982, 33: 317-345.
[20] 陈 昕, 徐宜凤, 张振英. 干旱胁迫下石灰花楸幼苗叶片的解剖结构和光合生理响应[J]. 西北植物学报, 2012, 32(1): 111-116.
[21] 张诚诚, 文 佳, 曹志华, 等. 水分胁迫对油茶容器苗叶片解剖结构和光合特性的影响[J]. 西北农林科技大学学报(自然科学版), 2013, 41(8): 79-84.
[22] 郭素枝, 张明辉, 邓传远, 等. 干旱胁迫对茉莉3个品种叶片光合特性和超微结构的影响[J]. 热带亚热带植物学报, 2013, 21(3): 123-129.
[23] 马旭俊, 朱大海. 植物超氧化物歧化酶(SOD)的研究进展[J]. 遗传, 2003, 25(2): 225-231.
[24] 王红梅, 包维楷, 李芳兰. 不同干旱胁迫强度下白刺花幼苗叶片的生理生化反应[J]. 应用与环境生物学报, 2008, 14(6): 757-762.
[25] Fazeli F, Ghorbanli M, Niknam V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars[J]. Biology Plantarum, 2007, 51(1): 98-103.
[26] Selote D S, Khanna C R. Drought acclimation confers oxidative stress tolerance by inducing coordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings[J]. Physiology Plantarum, 2006, 127: 494-506.
[27] Smirnoff N. The role of active oxygen in the response of plants to water deficit and desiccation[J]. New Phytologist, 1993, 125: 27-58.
[28] Bowler C, Montagu M V, Inzc D. Superoxide dismutase and stress tolerance[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1992, 43(1): 83-116.
[29] 赵丽英, 邓西平, 山 仑. 活性氧清除系统对干旱胁迫的响应机制[J]. 西北植物学报, 2005, 25(2): 413-418.
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
江苏农业科学(2016年8期)2017-02-15 19:38:57
江苏农业科学(2016年8期)2017-02-15 19:26:11
安徽农学通报(2016年24期)2017-01-12 21:06:57
安徽农学通报(2016年24期)2017-01-12 21:04:01
现代园艺(2016年7期)2017-01-09 14:06:28
山东农业科学(2016年11期)2016-12-17 21:16:16
Coco薇(2016年5期)2016-06-03 09:17:41
现代园艺(2016年2期)2016-03-15 16:05:02
考试周刊(2016年6期)2016-03-11 08:14:32
