
生态位构建作用对捕食-竞争系统的影响
2016-05-17 11:24莫昌健韩晓卓
生态科学 2016年5期
莫昌健, 韩晓卓
生态位构建作用对捕食-竞争系统的影响
莫昌健1,2, 韩晓卓1,*
1. 广东工业大学, 应用数学学院, 广州 510512
2. 广东松山职业技术学院, 基础部, 韶关 512000
生态位构建是指有机体所具有的改变其生存环境以促进自身生存与发展的能力。文章通过数值模拟竞争环境下具有捕食关系的各物种间的相互作用, 探讨了生态位构建作用对于捕食-竞争系统中物种数量变化及其空间分布模式的影响。研究结果表明: 物种的生态位构建行为不会破坏生态环境的稳定性, 但能有效提高构建物种的竞争力, 维持其在不利环境中与竞争物种共处的平衡状态; 同时, 生态位构建行为加强了构建物种的空间扩散能力, 促使构建物种迅速迁移至有利于自身生存的空间生境中, 成为捕食-竞争系统中的稳定生存物种; 而由于可供分配的资源有限, 有机体的构建行为与扩张行为之间存在的trade-off关系使其空间分布区域维持在一定的平衡状态, 不会出现无限制的空间扩散。
生态位构建; 反应扩散方程; 妥协; 平衡状态
1 前言
生态位构建是指有机体通过新陈代谢、活动和选择, 部分的创建和部分的毁灭自身生态位、改变环境, 进而改变其环境中生物与非生物的自然选择源的一种能力[1–5]。自生态位构建理论提出以来, 一系列关于生态位构建的研究成果的发表, 使得该理论逐渐引起生态学家与进化生物学家的兴趣[6–8]。2004年惠苍[9]首先将生态位构建作用架构到集合种群水平上; 2006年韩晓卓等人[10–11]讨论了具有生态位构建的有机体或种群对破坏生境的重新修复作用以及相应的集合种群动态与空间分布的变化。Kylafis和Loreau于2008年[12]研究了单一物种的生态位构建对于环境及进化的影响, 并于2011年[13]以Leibold[4]所提出的模型为基础, 构建了具有生态位构建行为的捕食-竞争系统模型。2014年韩晓卓和惠苍[14]研究了环境梯度下生态位构建行为对于物种基因分布及适应性景观的影响, 同年Buser[15]研究了生态位构建行为对于互利共生系统下生物进化的影响。经过学者的不断补充完善, 生态位构建理论已在种的适合度测定[16]、优势度分析[17]、种间竞争与共存[18–19]、群落结构与多样性维持[20–23]、物种生态位构建与进化对策[24–25]等研究中广泛应用, 关于生态位构建的定量研究[26]亦逐渐受到重视。
本文以Kylafis和Loreau提出的捕食-竞争系统为基础, 建立了捕食-竞争系统的反应扩散模型。通过数值模拟研究, 探讨了生态位构建对于捕食-竞争中各物种变化趋势及其空间扩散的影响。研究结果表明, 生态位构建行为在不破坏系统稳定性的同时,明显提升了构建物种的竞争力, 使其在不利环境中依然能与竞争物种共处; 同时, 生态位构建行为加强了物种的空间扩散能力, 使得物种能够迅速迁徙到更适合自己生存的空间, 成为捕食-竞争系统中的稳定生存物种。这一研究结果从数值角度阐释了生态位构建对于物种数量以及空间扩展的影响, 拓展了Kylafis和Loreau的研究结果。
2 模型
2.1捕食-竞争系统
生态位构建是指有机体对其环境上自然选择源的改变, 而在生态时间尺度上, 其重点强调物种的构建行为对其自身所在环境的改变和影响。在物种的实际生存中, 物种数量主要受到两个因素的限制:分别是有限的自然资源和捕食者自身对于物种的捕食现象。据此, 可以建立一个基本的捕食-竞争系统(系统1): 该系统包含A和B两个竞争物种和一个捕食者P。为了引入生态位构建的作用, 在该模型中同时考虑自然资源R的动态, 其中两竞争物种通过摄取自然资源R而生存; 捕食者P则以捕食两竞争物种作为自身生存的前提。模型具体表示如下(系统1):

其中i=A, B,im和ic分别表示捕食者对物种i 的捕食效率和转化效率,if和ia表示竞争物种i对资源的摄取效率及转化效率,pd,id则代表捕食者P和竞争物种i的自然死亡率, k代表自然资源自身的转化效率, 而S则代表自然资源的供应总量。
生态位构建的作用可以通过两方面来体现, 一种是直接的方式, 即物种通过构建有利于自身生存的环境以加强自身对于环境的适应性, 另一种则是间接的方式, 即物种通过构建抑制竞争物种生存的环境以降低竞争物种对于环境的适应性。本文重点考虑前者, 在系统(1)的基础上针对物种A, 通过增加其抵抗捕食者的被捕食能力, 建立具有生态位构建作用的捕食-竞争系统, 模型表示如下(系统2):

其中参数Pn为竞争物种A对捕食者的生态位构建强度, 当pn=0时, 系统(2)退化为系统(1), 其他参数含义同系统(1)。
为简化模拟与讨论, 考虑到物种对资源的转化效率对于其在生存竞争中所产生的影响甚微, 假设各竞争物种对于自然资源R的转化效率以及捕食者P对于各竞争物种的转化效率均为理想状态, 即设定参数ci, ai, k, s均取值为1; 同时, 为了更好的反映各物种间的关系, 进一步假设捕食-竞争系统中竞争物种A和物种B具有如下层次特征: 即物种A相对于B而言, 具有较强的对自然资源的摄取能力,但更易受到捕食者P的捕食; 这两者的竞争特征反映在系数上则体现为: mA>m B, fA>fB。在数值模拟中, 通过固定参数fA、fB及mB, 选择变化mA和nP的方法, 对上述两个系统的稳定性及各物种数量的变化趋势进行对比分析, 以清楚地了解生态位构建作用对捕食-竞争系统的影响。
2.2捕食-竞争系统反应扩散模型
从空间生态学角度上讲, 环境的异质性是导致物种形成及分布的重要因素, 也是空间模拟的主要原因。其中, 偏微分方程及反应扩散方程一直是数学生态学研究的主要手段。经典的常微分方程描述生物系统的时变动态, 而偏微分方程则可以给出空间上每一点的时间动态及密度分布[27]。因此, 为了进一步探讨生态位构建作用对物种空间分布动态的影响, 我们在上述系统(1)和(2)的基础上分别建立了具有空间扩散项的偏微分方程。其中, 反应项即为上述系统(1)和(2)的主体部分, 描述物种自身的增长动态; 而扩散项则由布朗随机运动描述, 视有机体在空间生境中做布朗随机运动且运动的速率是恒定的无方向差别量。依据假设, 相应的扩散模型分别为基本捕食-竞争系统的反应扩散模型(系统3)和具有生态位构建的捕食-竞争系统的反应扩散模型(系统4), 表示如下:

其中PD、AD、BD分别表示物种P、A、B物种的扩散系数。首先, 为了单纯观察生态位构建对于物种扩散的影响, 将扩散系数统一设置为1; 其次, 进一步考虑到有机体自身用于生长、生命维持以及繁殖的时间、能量和其他资源总是有限的, 其必须在这些可能的需求方面分配有限资源[28]。对于该系统,需求方面的冲突会导致生态位构建能力与扩散能力之间的妥协。因此, 假设AD+Pn=1。选取一个大小为nn×(n = 300)的空间显含模式的正方形网格区域,其边界条件为零流量输出边界条件。通过空间模拟方法, 对两种不同扩散因子假设下的模型中分别进行对照分析, 从而了解生态位构建对于捕食-竞争系统中各物种空间扩散的影响。
3 结果与分析
3.1捕食-竞争系统的稳定性分析
为了更好的反映生态位构建对于捕食—竞争系统中各物种数量变化趋势的影响, 首先利用常微分方程的稳定性理论分析系统的非平凡稳定平衡点及平衡条件, 以确保后续的讨论是在系统中捕食者P和竞争物种A、B共存的条件下进行的。
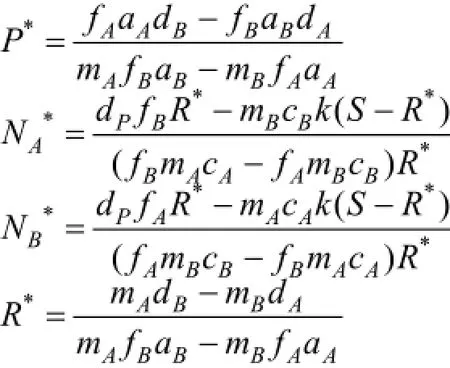
类似的, 通过求解系统(2)的平衡点, 不难得出系统(2)稳定平衡的限制条件为: (2.1)。通过对比两系统的限制条件, 发现相较于系统(1)的稳定条件, 系统(2)的差异在于构建系数Pn的引入, 因此,下文将着重分析该参数对系统稳定性及物种间数量关系的影响。
3.1.1 捕食-竞争系统(系统1)的时间动态
数值模拟结果显示: 捕食者P, 竞争物种A、B动态呈现阻尼振荡趋势(见图1a), 三者的相空间曲线最终螺旋汇聚于内部平衡点(见图1b)。进一步观察系统(1)的动态变化趋势(见图2)发现: 当mA=0.21时, 物种A处于优势并渐趋稳定, 物种B处于劣势并渐趋灭亡; 当mA=0.2119时, 两竞争物种维持平衡状态; 当mA=0.4时, 竞争物种A迅速消亡, 物种B取代物种A成为竞争中的稳定生存物种。以上动态变化趋势显示与的接近程度影响着两物 种的数量变化, 当与十分接近时, 物种A在竞争中处于优势, 当远大于时, 物种B 取代物种A, 成为竞争中的稳定生存物种。

图1 系统(1)的三物种动态图(图1a)及相空间图(图1b)Fig. 1 The diagrams of species temporal dynamics (a) and phase plane (b) of system (1)
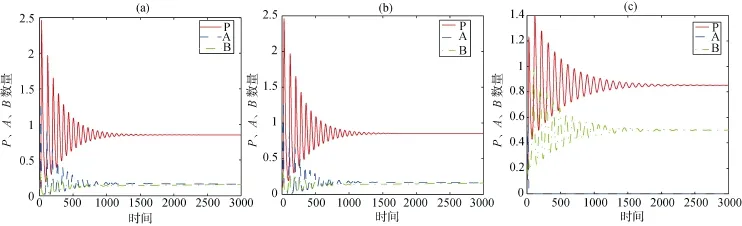
图2 系统(1)的物种数量动态图, (a)、(b)、(c)所对应的mA分别为0.21, 0.2119和0.4Fig. 2 The temporal dynamics of three species in system (1); the parameters of mAin (a), (b) and (c) are 0.21, 0.2119 and 0.4.
3.1.2 具有生态位构建项模型(系统2)的时间动态
数值模拟结果显示, 系统(2)的相空间图(见图3)及物种数量动态趋势图(见图4)与系统(1)类似, 均为围绕内部平衡点的阻尼振荡动态。仔细观察, 不难发现: 当Pn=0.1885时, 物种A和B的数量水平相当; 而当构建系数继续增加时, 系统(2)出现了异于系统(1)的情形: 物种A取代物种B成为竞争中的优势方, 物种B渐趋消亡。
对比上述两组结果, 可以看出生态位构建系数nP的引入并未改变捕食-竞争系统的稳定性, 但是生态位构建行为提升了构建物种A的竞争力, 使该物种不仅能在不利环境中(Am的增加表明竞争物种A所面临的被捕食压力越来越大)维持与物种B共处的平衡状态, 而且在不断增加构建系数的情况下,取代物种B成为竞争中的稳定生存物种。
3.2捕食-竞争系统的空间动态
对于反应扩散模型的模拟, 根据其数值模拟对于时间空间步长的依赖性和敏感度, 我们合理的选择了较小的时空步长以尽可能降低模拟误差。结果显示, 对于系统3, 当其不具有构建因素时, 捕食者以及竞争物种主要聚集在初始生存区域, 并未发生明显的空间扩散行为(图5)。当竞争物种A具有生态位构建行为时, 各物种的生存区域发生了明显变化(图6): 特别是具有构建行为的物种A, 因为构建行为提升了其躲避捕食者P的能力, 所以其表现出离开原有的生存区域、并迁移至生存风险较低的生存区域的空间扩张行为; 对于不具有构建行为的竞争物种B, 其生存环境与能力并未发生改变, 但其相对于构建物种A, 其生存能力在减弱, 因此其生存区域逐渐收缩; 而捕食者P则因为竞争物种A的迁移也相应的扩大了自己的捕食范围, 进而扩大了自身的生存区域。

图3 系统2的相空间图, (a)、(b)、(c)所对应的nP分别为0.1, 0.1885和0.2Fig. 3 The diagram phase plane of system (2); the parameters of npin (a), (b) and (c) are 0.1, 0.1885 and 0.2.
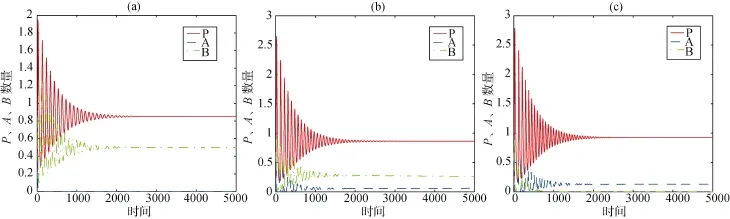
图4 系统2的物种数量动态图, (a)、(b)、(c)所对应的nP分别为0.1, 0.1885和0.2Fig. 4 The temporal dynamics of three species in system (2); the parameters of npin (a), (b) and (c) are 0.1, 0.1885 and 0.2.
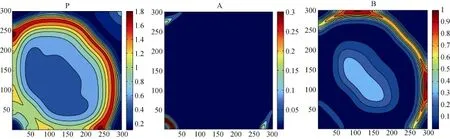
图5 模型3中各物种的空间分布图, 参数为: fA=0.2, mA=0.4, fB=0.1, mB=0.1, T=3000.Fig. 5 The spatial distribution of three species in model (3); the parameters are fA=0.2, mA=0.4, fB=0.1, mB=0.1, T=3000.
可以注意到, 在其他环境参数不变的情况下,一方面, 若不考虑生态位构建与扩散能力间的妥协关系, 构建系数的变化也会影响系统中各物种的空间分布变化。模拟结果(图7,图8和图9)表明: 当构建系数逐渐加强时, 在物种B的生存区域逐渐收缩的同时, 构建物种A和捕食者P的生存区域及空间密度发生显著变化.其中构建物种A的生存区域逐渐扩大, 空间密度也在逐渐增加, 当达到一定阈值(Pn=0.1885)后, 物种A出现了回溯迁移的空间扩展行为; 另一方面, 对于捕食者P而言, 当构建系数较小时, 其生存区域以及空间密度都逐渐提升, 但随着构建系数的逐渐增加, 其生存区域开始出现收缩, 边界空间密度开始降低。

图6 模型4中各物种的空间分布图, 参数值Pn=0.1, 其他参数同图5.Fig. 6 The spatial distribution of three species in model (4);Pn=0.1, other parameters are same as those in Fig.5.
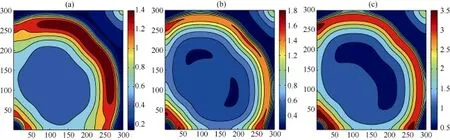
图7 模型4中物种P的空间区域在不同Pn下的快照, (a)、(b)、(c)中的Pn分别为0.1, 0.1885, 0.3Fig. 7 Snapshots of species P distribution with differentPn; the parameters in (a), (b) and (c) are 0.1, 0.1885, 0.3.

图8 模型4中物种A的空间区域在不同Pn下的快照,Pn参数同图7.Fig. 8 Snapshots of species A distribution with differentPn; the parameters are same as those in Fig.7.
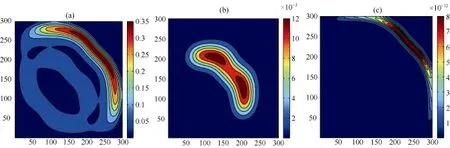
图9 模型4中物种B的空间区域在不同Pn下的快照,Pn参数同图7.Fig. 9 Snapshots of species B distribution with differentPn; the parameters are same as those in Fig.7.
同时, 物种的构建行为对于其空间扩散的作用会受到trade-off效应的制约, 从而影响着系统中各物种的空间分布, 模拟结果(见图10, 图11和图12)显示: 当模型4受到trade-off效应影响时, 对于不具有构建行为的物种B而言, 其空间区域及空间密度较之原模型(见图7)明显减弱; 而对于构建物种A及捕食者P而言, 虽然二者的空间密度并未发生变化,但它们的空间分布区域基本维持在Pn=0.5时的区域面积, 并未出现进一步的扩散。由此可以看出, 构建行为虽然会促进物种空间扩散, 但由于物种自身能力所存在的trade-off效应, 这种促进作用会随着构建系数的增加而逐渐降低, 使得物种的空间区域维持在一定的平衡状态, 不会出现无限制的空间扩散。
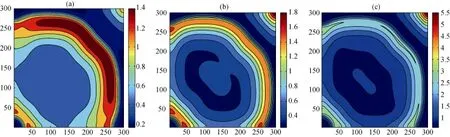
图10 具有trade-off效应的模型4中物种P的空间区域在不同Pn下的快照,Pn参数同图7.Fig. 10 Snapshots of species P distribution with trade-off effects; the parametersPn are same as those in Fig.7.
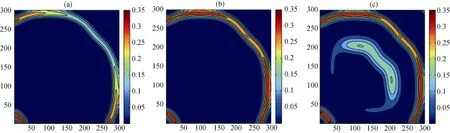
图11 具有trade-off效应的模型4中物种A的空间区域在不同Pn下的快照,Pn参数同图7.Fig. 11 Snapshots of species A distribution with trade-off effects; the parameters are same as those in Fig.7.
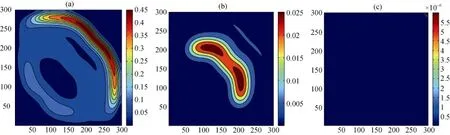
图12 具有trade-off效应的模型4中物种B的空间分布在不同Pn下的快照,Pn参数同图7.Fig. 12 Snapshots of species B distribution with trade-off effects; the parameters are same as those in Fig.7.
4 讨论与总结
本文通过对具有生态位构建行为的捕食-竞争系统和考虑到空间扩散因素而建立的反应扩散模型的数值模拟和比较分析, 研究了物种的构建行为对捕食-竞争系统演化动态和空间分布模式的影响。首先, 当物种不存在生态位构建行为时, 对自然资源的摄取能力较强但更易受到捕食的物种A的数量变化趋势是前期优势、后期劣势并渐趋消亡; 而摄取能力较弱且不易被捕食的物种B的数量变化趋势则与之恰好相反, 并最终取代竞争物种A; 当竞争物种A具有生态位构建行为时, 系统的稳定性并未发生改变, 但明显的, 物种A通过构建行为提高了自身竞争力, 使得其在不利环境中依旧能与竞争物种B共存, 甚至取而代之成为系统的稳定生存物种。因此,生态位构建行为也是导致物种稳定共存的因素之一,这一结果得到之前多项研究的支持[30–31], 特别是惠苍等人2004年[9]的研究表明生态位构建的机制可能是解释许多群落中竞争共存的潜在模式。本文的研究结果再一次说明有机体的生态位构建行为理应在生态学研究中备受重视[7]。
本文的重点在于考虑了具有扩散因子的生态位构建物种在竞争与捕食双重作用下的反应扩散方程。严格意义上来讲, 这是首次在生态位构建的研究工作中使用偏微分方程这一工具。本研究的模拟结果充分展示了构建行为对具有扩散因子的物种的空间分布动态的影响。其中, 具有生态位构建作用的物种A会离开原有的生存区域, 并因扩散因素而迁移至生存风险更低的空间区域; 捕食者P则由于物种A的迁徙而相应的扩大了自身的捕食区域, 进而也扩展了自身生存区域。尤其是随着构建系数的增加, 物种A和捕食者P的生存区域及空间密度发生了明显的变化。其中构建物种A的生存区域逐渐扩大, 当达到一定阈值后, 物种A出现了回溯迁移的空间扩展行为, 此时, 捕食者P生存区域也开始出现收缩, 且边界空间密度降低。在这一结果中, 阈值的出现和空间扩张行为的回溯现象是源于具有构建行为的有机体与环境之间的反馈机制。这也是生态位构建理论的核心, 即有机体与环境之间相辅相成、相互影响与制约的正反馈作用能够产生丰富的进化现象与生态学动态[5]。
Trade-off关系是指有机体由于可供分配的资源有限, 特定两个特征间的相互作用, 即提高一种特征的优势的时候, 另一种特征的优势将降低。现已开展了诸多关于trade-off关系的理论研究, 例如均质斑块环境上的集合种群竞争共存的关键是侵占能力与竞争能力之间的妥协[32]。Tilman的模型提供了一个由固着个体组成的许多群落中高物种丰富度的完美解释。另外, 到目前为止, 促进稳定共存的因子已经被很好地研究和讨论过。例如, 空间细分和局部扩散会导致奇异的空间动态并使捕食系统动态稳定[31]。侵占-竞争妥协会形成竞争者与逃逸者的稳定共存[32]。惠苍等人2004年[9]的研究则说明空间尺度的物种共存可能得益于竞争力、侵占力以及生态位构建力之间巧妙的妥协。而本文基于生态位构建能力和扩散能力间妥协关系的假设, 研究结果表明:生态位构建行为对于物种空间扩散的促进作用并不会导致物种无限制的空间扩张, 而是随着构建系数的增加而逐渐降低, 进而维持物种空间区域的平衡状态。
Kylafis和Loreau于2011年[13]曾对捕食-竞争系统进行了初步的理论研究, 他们认为: 生态位构建有助于增加构建物种抵抗捕食者的能力, 维持构建物种在不利条件下与竞争物种共处的平衡状态, 这与本文的数值模拟结果一致。进一步对系统中各物种添加扩散项后的模拟结果证实, 构建行为还有助于提升构建物种的空间扩展能力, 扩展其生存区域。这一研究将原有的研究从之前单一的物种数量变化扩展至生存区域的变化, 对于探讨生态位构建行为对物种生态分布及种群变化趋势的影响有着重要意义。由于本文的模型是在理想化情况下构建的,并未考虑实际物种共存中存在的Allee效应等问题, Allee效应会对具有构建行为的捕食-竞争系统中各物种的数量变化及其空间扩展造成怎样的影响有待进一步研究。
致谢本文得到国家自然科学青年基金资助项目-生态位构建对环境梯度下物种分布格局的作用(31100308)的资助。
参考文献
[1] LALAND K N, ODLING-SMEE F J, FELDMAN M W. The evolutionary consequences of niche construction: a theoretical investigation using two-locus theory[J]. Journal of Evolutional Biology, 1996, 9: 293–316.
[2] DAY R L, LALAND K N, ODLING-SMEE F J. Rethinking Adaptation. the niche-construction perspective[J]. Perspectives in Biology and Medicine, 2003, 46: 80–95.
[3] ODLING-SMEE F J, LALAND K N, FELDMAN M W. Niche construction: the neglected process of evolution[M]. Princeton University Press, 2003.
[4] LEIBOLD M A. A graphical model of keystone predators in food webs trophic regulation of abundance, incidence, and diversity patterns in communities[J]. American Naturalist, 2011, 147: 784–812.
[5] ODLING-SMEE F J, ERWIN D H, PALKOVACS E P, et al. Niche Construction Theory: A Practical Guide for Ecologists[J]. The Quarterly Review of Biology, 2013, 88(1): 3–28.
[6] O’BRIEN M J, LALAND K N. Genes, culture, and agriculture: an example of human niche construction[J]. Current. Anthropology, 2012, 53: 434–470.
[7] MATTHEWS B, MEESTER L D, JONES C G, et al. Under niche construction: an operational bridge between ecology, evolution, and ecosystem science[J]. Ecological Monographs, 2014, 84(2): 245–263.
[8] PALKOVACS E P, KINNISON M T, CORREA C, et al. Fates beyond traits: ecological consequences of humaninduced trait change[J]. Evolutionary Applications, 2012, 5: 183–191.
[9] HUI Cang, LI Zizheng, YUE Dongxia. Metapopulation dynamics and distribution, and environmental heterogeneity induced by niche construction[J]. Ecological Modelling, 2004, 177: 107–188.
[10] HAN Xiaozhuo, LI Zizheng, HUI Cang, et al. Polymorphism maintenance in a spatially structured population: two-locus genetics model of niche construction[J]. Ecological Modelling, 2006, 192: 160–174.
[11] HAN Xiaozhuo, LI Zizheng, ZHANG Feng, et al. Effect of niche construction on metapopulation dynamics by modifying destroyed habitat[J]. Journal of Lanzhou University, 2006, 43(1): 56–64.
[12] KYLAFIS G, LOREAU M. Ecological and evolutionary consequences of niche construction for its agent[J]. Ecology Letters, 2008, 11: 1072–1081.
[13] KYLAFIS G, LOREAU M. Niche construction in the light of niche theory[J]. Ecology Letters, 2011, 14: 82–90.
[14] HAN Xiaozhuo, HUI Cang. Niche Construction on Environmental Gradients: The Formation of Fitness Valley and Stratified Genotypic Distributions [J]. PLoS One, 2014, 9(6): e99775.
[15] BUSER C C, NEWCOMBANNE R D, GASKETT A C, et al. Niche construction initiates the evolution of mutualistic interactions[J]. Ecology Letters, 2014, 17: 1257–1264
[16] BRODAW N, BUSING P T. Niche versus chance and tree diversity in forest gaps[J]. Trends in Ecology & Evolution, 2000, 15: 183–188.
[17] HUI Cang, ROURA-PASCUAL N, BROTONS L, et al. Flexible dispersal strategies in native and non-native ranges: environmental quality and the ‘good-stay, bad-disperse’rule[J]. Ecography, 2012, 35: 1024–32.
[18] HURFORD J R. Niche construction, co-evolution and domain specificity[J]. Behavioral and brain Sciences, 2008, 31(5): 526–526.
[19] IHARA Y, FELDMAN M W. Cultural niche construction and the evolution of small family size[J]. Theoretical Population Biology, 2004, 65: 105–111.
[20] BERTHOULY-SALAZAR C, VAN RENSBURG B J, LEROUX J J, et al. Spatial sorting drives morphological variation in the invasive bird, Acridotheres tristis[J]. PLoS ONE, 2012, 7: e38145.
[21] ERWIN D H. Macroevolution of ecosystem engineering, niche construction and diversity[J]. Trends in Ecology & Evolution, 2004, 23(6): 304–311.
[22] HORAK J, HUI Cang, ROURA-PASCUAL N, et al. Changing roles of propagule, climate, and land use during extralimital colonization of a rose chafer beetle[J]. Naturwissenschaften, 2013, 100: 327–36.
[23] HUI Cang, LI Zizheng, YUE Dongxia. Metapopulation dynamics and distribution, and environmental heterogeneity induced by niche construction[J]. Ecological Modelling, 2004, 177: 107–188.
[24] HAN Xiaozhuo, LI Zizheng, ZHANG Yanyu. Effects of time-lagged niche construction on metapopulation dynamics and environmental heterogeneity[J]. Applied Mathematics & Computation, 2009, 215: 449–458.
[25] PALMER M E, FELDMAN M W. Survivability is more fundamental than evolvability[J]. PLoS One, 2012, 7: e38025.
[26] HAN Xiaozhuo, LI Zizheng, HUI Cang, et al. Theory of niche construction and application of its two-locus population genetic model[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(3): 558–562.
[27] HUI Cang. Simulated research on spatial Ecology: the ecological and evolutionary responses of metapopulation to environmental changes[D]. Lanzhou: Lanzhou University, 2004: 7–11.
[28] SILVERTOWN J W, DOUST J L. Introduction to plant population biology[M]. Blackwell, Oxford. 1993.
[29] 崔丽娟, 李伟, 赵欣胜, 等. 采砂迹地型湿地恢复过程中优势种群生态位研究[J]. 生态科学, 2013, 32(1): 073–077.
[30] 袁丛军, 喻理飞, 安明态, 等. 岩生红豆天然林主要树种生态位研究[J]. 生态科学, 2013, 32(4): 487–493.
[31] HASSELL M P, COMINS H N, MAY R M. Spatial structure and chaos in insect population dynamics[J]. Nature, 1991, 353: 255–258.
[32] TILMAN D. Competition and biodiversity in spatially structured habitats[J]. Ecology, 1994, 75: 2–16.
The effects of niche construction on prey-competition system
MO Changjian1,2, HAN Xiaozhuo1,*
1.School of Applied Mathematics,Guangdong University of Technology,Guangzhou510512,China
2.Foundation Department,Guangdong Songshan Polytechnic College,Shaoguan512000,China
Niche construction is the process whereby an organism improves its environment to enhance its growth and persistence. Based on a prey-competition system with two competitive consumers and one predator, we included direct and indirect ways of niche construction by consumers and also the factor of spatial diffusion. Through numerical simulation, the effect of niche construction on temporal dynamics and spatial distribution of different species in the system were explored. Our results indicated that the niche construction did not destroy the stability of the prey-competition system. Niche- constructing species, however, can enhance its own competitive strength and also maintain the system balance in adverse environments. In addition, the niche construction has the potential to improve a diffusivity of the niche-constructing species, which promotes them to immigrate for more suitable habitats and become the stable species in the system. Because of the limit resource, the trade-off relationship between the niche construction and the diffusion of species imposed restrictions on the distribution range, inhabiting species unlimited expansion.
niche construction; reaction-diffusion equations; trade-off; equilibrium state
10.14108/j.cnki.1008-8873.2016.05.011
Q141
A
1008-8873(2016)05-073-09
莫昌健, 韩晓卓. 生态位构建作用对捕食‐竞争系统的影响[J]. 生态科学, 2016, 35(5): 73-81.
MO Changjian, HAN Xiaozhuo. The effects of niche construction on prey-competition system[J]. Ecological Science, 2016, 35(5): 73-81.
2015-03-12;
2015-04-25
国家自然科学青年基金项目(31100308)
莫昌健(1986—), 男, 广东韶关人, 硕士, 主要从事数学生态学研究
*通信作者: 韩晓卓(1978—), 女, 青海西宁人, 博士, 副教授, 主要从事数学生态学研究, E-mail: hanxzh@gdut.edu.cn
猜你喜欢
数学物理学报(2021年6期)2021-12-21
科学大众(中学)(2019年3期)2019-05-17
数学物理学报(2018年5期)2018-11-16
汽车观察(2018年10期)2018-11-06
太原师范学院学报(自然科学版)(2018年2期)2018-08-17
现代企业文化·综合版(2017年5期)2017-06-14
科技知识动漫(2017年1期)2017-02-06
中外文摘(2016年13期)2016-08-29
人生十六七(2015年26期)2015-08-22
少儿科学周刊·少年版(2015年1期)2015-07-07
