
毛竹的花药发育研究
2016-05-09 05:27:34周建梅孙立方郭起荣
广西植物 2016年2期
周建梅,孙立方,2,冯 云,廉 超,冉 洪,张 莹,郭起荣∗
(1.国际竹藤中心国家林业局竹藤科学与技术重点开放实验室,北京100102;2.南开大学生命科学院,天津300071)
毛竹的花药发育研究
周建梅1,孙立方1,2,冯 云1,廉 超1,冉 洪1,张 莹1,郭起荣1∗
(1.国际竹藤中心国家林业局竹藤科学与技术重点开放实验室,北京100102;2.南开大学生命科学院,天津300071)
竹类植物因有着较长的开花周期,其生殖生物学研究的报道相对较少。该研究采用石蜡切片与野外观察的方法,对毛竹花药的发育以及花药发育与花序的关系进行了研究。结果表明:毛竹的花药壁结构包括4层细胞:表皮细胞、药室内壁细胞、中层细胞和绒毡层细胞。药室内壁和中层都只有一层细胞,而且细胞形状较扁,花药发育后期药室内壁会逐渐降解,而中层则会完全解体消失。花药壁的发育为单子叶型,绒毡层为腺质型,而且只有一层,细胞径向较长,最后也会消失。小孢子母细胞减数分裂时,胞质分裂方式为连续型。形成的小孢子经一次有丝分裂后逐渐形成成熟花粉粒,大多为二细胞型,很少产生三细胞型。此外,还发现毛竹花药的发育与花序形态变化存在着相对应的关系。野外连续观察和切片发现,随着花序形态的不断发育变化,首先花药开始形成并不断分化,药壁各层也逐渐形成;接着小孢子逐渐成熟,各层也慢慢随之解体、消失;最后花药逐渐开裂并开始散粉。该研究结果不仅丰富了毛竹和竹类生殖生物学的研究内容,而且对毛竹种质的研究也具有重要意义。
毛竹,花药,花药壁,小孢子,花序,发育
竹类植物是禾本科(Poaceae/Gramineae)竹亚科(Bambusoideae)植物的总称,全世界约有88属1 400余种,其中我国竹类资源十分丰富,据不完全统计,共有34属534种。我国以占3%森林面积的竹林,产生了超过18%的营林产值。据国家林业局2014年最新公布的第八次全国森林资源清查(2009 -2013)数据,我国竹林面积601万hm2,占整个森林总面积在3%以上,占世界竹林总面积的1/3。作为一类生物学特性特殊的植物,竹子的开花时间在空间上不确定,形态上较为相似。竹类植物一般开花周期较长,多数一生只开一次花(Isagi,2004),植株开花多数死亡,这给竹类植物的生殖生物学研究工作带来了很大的困难。近期已有对雷竹(Phyl-lostachys violascens)(何奇江等,2003)、巨龙竹(Den-drocalamus sinicus)(王曙光等,2006)、鹅毛竹(Shi-bataea chinensis)、异叶苦竹(Arundinaria simonii)(林树燕,2009,2012)、月月竹(Menstruocalamus sichua-nensis) (刘静,2008;林树燕等,2009)、寒竹(Chi-monobambusa marmorea)(胡成华等,1994)等竹种的生殖生物学进行了研究。
毛竹(Phyllostachys edulis)属禾本科(Poaceae)竹亚科(Bambusoideae)刚竹属(Phyllostachys)。竿高在20 m或以上,直径在20 cm以上,节间在40 cm以上。毛竹是我国特有的、最重要的经济竹种,也是我国分布最广、栽培历史长、栽培面积最大的竹种,据第八次全国森林资源清查报告显示,面积已达443万hm2,占竹林面积的74%,长期以来产生了巨大的经济、生态、社会效益(江泽慧,2002)。因此,毛竹在我国竹产业发展中占有极其重要的地位。毛竹种质资源是重要的遗传资源,蕴藏着对毛竹遗传改良和育种起关键作用的重要基因,是竹产业可持续发展的物质基础。像毛竹这样常见的种类,其生殖生物学研究仅见乔士义和廖光庐(1984)的报道,孙立方等(2011)报道了毛竹的花器官形态与结构。本研究探讨了毛竹的花药发育过程,不同花序发育时序中花药发育情况,为竹亚科及禾本科的生殖生物学研究提供资料。
1 材料与方法
1.1 材料
实验材料分别于2010-2012年,在广西桂北海洋山深山野生毛竹林中采集。该地属中亚热带地区,海拔近千米;年平均气温18~19℃,4-9月为雨季,年平均降水量1 750~2 000 mm。
1.2 方法
标记3株有开花征兆的毛竹植株,从小枝开始膨大变粗到果实成熟期间(6-9月),定期采样,每隔1~3 d采样,取不同发育时期的小穗、小花、花药,先用70%酒精的FAA固定液固定24~48 h,然后在4℃冰箱内保存。在变态小枝开始膨大变粗时进行拍照,观察花序发育的表型变化。采用常规石蜡切片法制片(李正理,1996),切片厚度为5~10 μm,番红—固绿染色,用加拿大树胶封片,在OLYMPUS CX31显微镜下观察、拍照。
2 结果与分析
大多数被子植物的花药具有4个小孢子囊(花粉囊),少数具有2个,它们由药隔组织分开。当花药达到成熟时,每一侧的2个小孢子囊往往随药隔消失而连通。每个小孢子囊包含药室和由数层细胞组成的花药壁。
2.1 花药壁的发育
研究中发现,毛竹的花药壁属于单子叶型,花药壁结构包括4层细胞:表皮细胞、药室内壁细胞、中层细胞和绒毡层细胞。
2.1.1 表皮 在毛竹花药发育过程中,表皮只进行垂周分裂增加细胞的数目以适应内部组织的迅速增长。初期表皮细胞(图版I:A)为近方形,随着花药的扩大,逐渐扩展成扁长形,其仍然是一般的行使保护的功能,因此通常也有明显的角质层。许多植物,特别是旱生习性的植物,表皮细胞最后彼此分离,当花药成熟时仅留下干枯的痕迹。毛竹花粉发育成熟时,其表皮细胞内物质逐渐减少和解体,仅剩下细胞壁,药室内花粉基本全部散出。
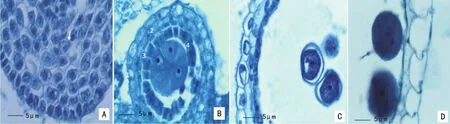
图版Ⅰ 毛竹的花药壁发育过程 A.初生壁细胞时期;B.小孢子母细胞时期4层药壁(1.表皮,2.药室内壁,3.中层,4.绒毡层);C.单核小孢子靠边期药壁,绒毡层解体;D.花粉粒成熟时期,仅有2层壁。PlateⅠ Anther wall development process of P.edulis A.Primary wall cells;B.4 layers of anther at the microspore mother stage(1.Epider-mis,2.Endothecium,3.Middle layer,4.Tapetum);C.Microspore is at binucleate stage,and tapetum is disintegrated;D.Only 2 layers of wall cells in the pollen grain mature period.

图版Ⅱ 毛竹小孢子与雄配子体发育 A.未分化花药;B.分化花药;C.初生壁细胞和初生造孢细胞;D.次生造孢细胞;E.小孢子母细胞;F.单核靠边期小孢子;G.小孢子收缩期;H.单核靠边期;I.2核花粉粒;J.成熟花粉粒和花药。PlateⅡ Microspore and development of male gametophyte of P.edulis A.Undifferentiated anther;B.Differentiated anther;C.Primary wall cells and primary sporogenous cells;D.Secondary sporogenous cells;E.Microsporoctyte mother cell;F.Tetrads;G.Contraction microspore;H. Middle microspore;I.2-nuclear pollen grain;J.Mature pollen grain and anther.
2.1.2 药室内壁 毛竹药室内壁只有一层细胞(图版I:B,2),药室内壁的内切向壁向外和向上发生带状或纤维状的加厚,因这一特点也称“纤维层”。药室内壁细胞的外切向壁仍保持是薄壁的,加厚的壁除带状的外,还有呈球状加厚、完整环带状等的变异形态。这种加厚壁的物质,一般认为是纤维素性质,在成熟时略为木质化。药室内壁的这种加厚的特点,有助于花粉囊的开裂。毛竹花药开裂时,靠近药隔维管束的药室外侧各层药壁细胞先逐渐溶解,然后开始开裂散粉。

表1 毛竹花序变化与花药发育的对应关系Table 1 Relationship between the changes of inflorescences and the development of anther of P.edulis
2.1.3 中层 毛竹中层1层细胞(图版I:B,3),由初生壁细胞内层分化而来,一般含有淀粉或其他贮藏物。中层出现在小孢子母细胞中后期,在小孢子母细胞末期时,中层的细胞已经开始发生变化,贮藏物质减少,细胞变为扁平,然后逐渐趋于解体和被吸收。在小孢子靠边期时,一般中层已完全解体消失(图版I:C)。
2.1.4 绒毡层 绒毡层是花药壁的最内层(图版I:B-4),也是由初生壁细胞内层分化而来,此层对小孢子或花粉的发育起重要的作用。在雄性不育系中花粉的发育与绒毡层机能失常相关,如绒毡层位置向药室中间移动或其原生质体侵向药室,挤压小孢子而导致不育,通过观察发现,毛竹绒毡层不存在此类情况。绒毡层通常为一层,少数植物具有二层或者多层,还有的植物是向表皮的绒毡层为单层,向药隔的为多层。小孢子后期时,绒毡层径向伸长。绒毡层的发育最终自我解体,小孢子靠边时期已经开始解体,雄配子体即成熟花粉阶段,绒毡层细胞仅留残迹或已不存在(图版I:C,D)。在整个发育过程中,缄毡层始终维持在原来的位置,因此,毛竹的绒毡层属于腺质绒毡层。
2.2 小孢子发生与雄配子体发育
2.2.1 小孢子发生 从毛竹雄蕊原基最初形成的一个很小的花药的横切面来看,外面是一层表皮,表皮以内是一群分裂活跃的细胞(图版Ⅱ:A)。后来,由于花药的四个角隅的细胞分裂较快,使花药变成为四个裂瓣的形状(图版Ⅱ:B),在每一瓣的表皮下分化出孢原细胞。孢原细胞经一次平周分裂和多次垂周分裂产生初生壁细胞和初生造孢细胞(图版Ⅱ: C),初生造孢细胞再次分裂,形成次生造孢细胞(图版Ⅱ:D),进而发育成小孢子母细胞(图版Ⅱ:E),小孢子母细胞核大,无明显的液泡,进一步发育,彼此进入减数分裂时期。小孢子母细胞减数分裂时,产生的小孢子四分体为左右对称型(乔士义和廖光庐,1984)(图版Ⅱ:F),胞质分裂方式为连续型。
2.2.2 雄配子体发育 小孢子四分体时期比较短暂,随着胼胝质溶解,小孢子释放出来,刚释放出来的小孢子细胞质浓厚,细胞核位于中央,为收缩期的小孢子(图版Ⅱ:G)。随后小孢子逐渐膨大,长大变圆,中央出现大液泡,细胞核移到细胞的一侧为单核靠边期的小孢子(图版Ⅱ:H)。整个细胞充满细胞质,细胞核进行一次有丝分裂,形成双核(图版Ⅱ:I),靠花粉壁的为生殖细胞,占据细胞中央的为营养细胞。毛竹的成熟花粉粒大多是二细胞型,很少产生三细胞型(图版Ⅱ:J)。
2.3 花药发育与花序发育的关系
根据观察发现,毛竹花药的发育变化与花序的发育有相对应的关系(表1)。随着花序的不断发育变化,花药不断分化(图版Ⅱ),药壁各层逐渐形成。小孢子逐渐成熟时,各层也慢慢随之解体、消失。
3 讨论与结论
毛竹的花药壁结构包括4层细胞:表皮细胞、药室内壁细胞、中层细胞和绒毡层细胞。药室内壁与中层细胞比较扁,在小孢子靠边期时,一般中层已完全解体消失;绒毡层细胞则径向较长,细胞质浓厚,绒毡层的发育最终自我解体。花药壁的发育为单子叶型,绒毡层为腺质型。小孢子母细胞减数分裂时,胞质分裂方式为连续型。成熟花粉粒大多是二细胞型,很少产生三细胞型。花药壁结构与前人所报道的巨龙竹、鹅毛竹、月月竹等竹类植物的花药壁结构一致,符合禾本科植物的基本特征。然而,不同于黄坚钦等(1999)报道的雷竹的花药壁由表皮层、2层中层和绒毡层4层细胞组成,且在整个发育过程中始终无药室内壁的形成与分化,表皮层发育起到药室内壁的作用。研究发现,毛竹花药壁的绒毡层为质腺型,与之前乔士义等报道的毛竹花药壁绒毡层为分泌型不同。
毛竹花序的外部形态特征与花药的发育有相对应关系。在发育早期,花序包裹在叶片内,为花原基、减数分裂等容易受外界影响的生理过程提供比较稳定的环境,保障生殖器官发育。随着花药的发育,成熟花粉粒形成,花序叶片变黄逐渐脱落暴露于空气中,为花粉散播做好准备。
HE QJ,WANG KH,HUA XQ,et al,2003.The observational study on spica and flower organs of Phyllostachys violascens[J].Zhe-jiang For Sci Technol,23(2):10-11. [何奇江,汪奎宏,华锡奇,等,2003.雷竹花穗和花器的观察研究[J].浙江林业科技,23(2):10-11.]
HU CH,YU FG,PANG YJ,et al,1994.The embryologic observa-tion and research of Chimonobambusa marmorea[J].J Bamb Res,13(4):6-11. [胡成华,喻富根,庞延军,等,1994.寒竹胚胎学观察与研究[J].竹子研究汇刊,13(4):6-11.]
HUANG JQ,HUANG HH,HE FJ,et al,1999.The formation of mi-crospore and the development of male gametophyte of Phyl-lostachys violascens[J].J Bamb Res,18(3):55-58. [黄坚钦,黄华宏,何福基,等,1999.雷竹的小孢子发生和雄配子体形成[J].竹子研究汇刊,18(3):55-58.]
JIANG ZH,2002.World bamboo[M].Shenyang:Liaoning Sci-ence and Technique Press:11-18. [江泽慧,2002.世界竹藤[M].沈阳:辽宁科技出版社:11-18.]
LI ZL,1996.lice-making of plant tissue[M].Beijing:Beijing U-niversity Press:129-137. [李正理,1996.植物组织制片[M].北京:北京大学出版社:129-137.]
LIN SY,DING YL,2012.The development of the female and male gametophyte of Shibataea chinensis[J].Acta Bot Boreal-Occident Sin,32(5):907-914. [林树燕,丁雨龙,2012.鹅毛竹大小孢子即雌雄配子体发育[J].西北植物学报,32(5):907-914.]
LIN SY,HAO JJ,XIN H,et al,2009.Megaspore and microspore occurre and female and male gametophyte of Menstruocalamus sichuanensis[J].J Nanjing For Univ:Nat Sci Ed,(3):9-12.[林树燕,郝娟娟,辛华,等,2009.月月竹大、小孢子发生和雌、雄配子体发育研究[J].南京林业大学学报:自然科学版,(3):9-12.]
LIN SY,2009.Studies on the reproductive biology of Shibataea chinensis and Arundinaria simonii f.albostriatus[D].Nanjing: Nanjing Forestry University. [林树燕,2009.鹅毛竹和异叶苦竹的生殖生物学研究[D].南京:南京林业大学.]
LIU J,2008.Studies on blossom biology of Chimonobambusa si-chuanensis[D].Nanjing:Nanjing Forestry University. [刘静,2008.月月竹开花生物学研究[D].南京:南京林业大学.]
QIAO SY,LIAO GL,1984.The embryologic observation of Phyl-lostachys edulis[J].Bamb Res,3(1):15-23. [乔士义,廖光庐,1984.毛竹的胚胎发育学观察[J].竹类研究,3(1):15-23.]
SUN LF,GUO QR,WANG Q,et al,2011.Flower organs morpholo-gy and structure of Phyllostachys edulis[J].For Sci,48(11): 124-129. [孙立方,郭起荣,王青,等,2011.毛竹花器官的形态与结构[J].林业科学,48(11):124-129.]
SUN LF,2012.The productive biological and seed yield increasing technique studies of Phyllostachys edulis[D].Beijing:Chinese Academy of Forestry Science. [孙立方,2012.毛竹生殖生物学和种实产量提高技术研究[D].北京:中国林业科学研究院.]
WANG SG,PU XL,DING YL,et al,2006.The structures of reproduc-tive organs and development of the female and male gametophyte of Dendrocalamus sinicus[J].Bull Bot Res,26(3):270-274. [王曙光,普晓兰,丁雨龙,等,2006.巨龙竹生殖器官形态结构及雌雄配子体的发育[J].植物研究,26(3):270-274.]
ZHOU FC,1998.Bamboo cultivation[M].Beijing:Chinese For-estry Press. [周芳纯,1998.竹林培育学[M].北京:中国林业出版社.]
Observation on anther development of Phyllostachys edulis
ZHOU Jian-Mei1,SUN Li-Fang1,2,FENG Yun1,LIAN Chao1,RAN Hong1,ZHANG Ying1,GUO Qi-Rong1∗
(1.International Center for Bamboo and Rattan;SFA Key Laboratory of Bamboo and Rattan Science and Technology,Beijing 100102,China;2.College of Life Sciences,Nankai University,Tianjin 300071,China)
Because of the long flowering stage,studies about the reproductive biology of bamboos were rarely fewer.In this research,the anther development of Phyllostachys edulis was studied by paraffin section,and the process of anther development of P.edulis was also analysed,including the development process of the anther,anther wall and microspore. The results showed that the anther wall of P.edulis contains four layer of cells,which were epidermal cells,endothecium cells,middle layer cells and tapetum cells.There was only a layer of cells with flat shape in endothecium cells and mid-dle layer cells.At the end of the anther development when microspore goes to pull over,endothecium cells would de-grade gradually and the middle layer cells would break up and disappeared.The development of anther wall was monocot-yledonous type,and the tapetum belonged to secretory type with a layer of cells growing in radial direction,the final de-velopment of tapetum disappeared by themselves.The type of cytokinesis of microsporocyte meioticdivision was succes-sive,and the generated microspore from which then fomated mature pollen by one mitosis process.Mature pollen grains were mostly two-celled,rarely three-celled.In addition,we found that there was a corresponding relationship between the development of anther and continuous morphology change of the inflorescence.Based on the results of continuous ob-servation on the inflorescence and paraffin section assays,we also found that unceasing development and division of an-ther occurred with the continuous development of inflorescence,and anther wall layers of cells were gradually formed. Then microspore became mature gradually and the layers of the anther degraded and disappeared during the process,and then mature pollen spilled out from the dehiscenced anther.Our research would enrich the studies on the reproductive bi-ology of bamboos and also was of great significance to the study on the germplasm of moso bamboo.
Phyllostachys edulis,anther,anther wall,microspore,inflorescence,development
S718.5
A
1000-3142(2016)02-0231-05
10.11931/guihaia.gxzw201405012
周建梅,孙立方,冯云,等.毛竹的花药发育研究[J].广西植物,2016,36(2):231-235
ZHOU JM,SUN LF,FENG Y,et al.Observation on anther development of Phyllostachys edulis[J].Guihaia,2016,36(2):231-235
2014-05-06
2015-07-12
国家科技支撑计划项目(2012BAD23B05)[Supported by National Key technology Research and Development Program of China(2012BAD23B05)]。
周建梅(1986-),女,重庆市人,硕士研究生,从事竹子种质研究,(E-mail)jianmeizhou@sina.cn。
∗通讯作者:郭起荣,男,教授、博导,一直从事森林(竹子)种质资源研究,(E-mail)QRGUO@icbr.ac.cn。
猜你喜欢
湖北农业科学(2021年19期)2021-11-15 07:01:38
云南农业大学学报(自然科学)(2021年3期)2021-06-11 04:19:38
广西植物(2016年10期)2016-11-11 06:51:39
西南农业学报(2016年4期)2016-05-17 05:41:45
地球学报(2015年5期)2015-06-06 10:45:11
地球学报(2015年5期)2015-06-06 10:45:11
中国瓜菜(2014年3期)2014-04-29 00:44:03
地方文化研究(2013年2期)2013-03-11 15:30:33
地方文化研究(2013年1期)2013-03-11 15:30:23
中国蔬菜(2013年8期)2013-01-28 04:52:48
